
Abstract
Purpose
This work was planned for providing a useful screening tool for the selection of Populus alba clones suitable for phytoremediation techniques. To this aim, we investigated variation in arsenic, cadmium, copper, and zinc tolerance, accumulation and translocation in three poplar clones through an in vitro screening. Poplars have been widely proposed for phytoremediation, as they are adaptable to grow on contaminated areas and able to accumulate metals. The investigation of possible differences among poplar clones in metal tolerance and accumulation deserves to be deeply studied and exploited for the selection of the more suitable tool for phytoremediation purposes.
Methods
In vitro multiplied microshoots of a commercial and two autochthonous P. alba clones were subcultured on hormone-free WPM medium for 1 month and then transferred for 2 weeks onto media containing different concentrations of the metals investigated. At the end of the treatments, plantlets were sampled, weighed, and mineralised by wet ashing. Metal concentrations were determined by ICP-OES.
Results
For the metal concentration used in the experiments, our clones of P. alba showed variation in metal tolerance, metal accumulation and content. The fast-growing commercial clone, even if rarely showing the highest plant metal concentration, displayed the highest metal content, suggesting biomass production as the key factor in evaluating the phytoextraction capacity of P. alba clones for the metals studied.
Conclusions
Data demonstrated that in vitro screening of cuttings represents a valuable way of assessing the ability of different poplar clones to take up, tolerate and survive metal stress.
Similar content being viewed by others

Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
1 Introduction
To remove contaminants from polluted sites, unconventional techniques involving biological processes can have strong potentialities. In particular, the so-called phytoextraction technology makes use of plants to extract metals from soil, thus being less expensive and less damaging to the environment than conventional remediation systems consisting mainly in soil excavation and incineration (Vassilev et al. 2004; Van Nevel et al. 2007). On the other hand, phytostabilization technology uses plants for stabilising contamination in place preventing further spreading and transfer into food chains (Vangronsveld et al. 2009).
The efficiency of phytoextraction depends, besides the substrate type, on several characteristics of the plant, such as the ability to accumulate and translocate metals to the aerial parts, a fast growth and a deep and extended root system.
Phytoextraction by woody plants (dendroremediation) presents several advantages. The large biomass yield can be used for economic opportunities, such as bioenergy, solid wood production and reconstituted products (Rockwood et al. 2004). The extended and deep root apparatus can explore a large volume of soil; the impact on trophic chains is low, and the adaptability of some tree species to grow in marginal soils is well-established. Metal uptake by trees is reported to be not as high as hyperaccumulators but, due to a greater yield of biomass, the removal of metals from soil could be more effective respect to hyperaccumulating plants (Fischerová et al. 2006; Mench et al. 2009; Vangronsveld et al. 2009). So, these plants may provide a viable alternative to the usual low-yielding hyperaccumulators that, in any case, are not well-represented at temperate latitudes (Dickinson et al. 2009).
Several hardwood species have been proposed for phytoremediation, and poplars are among these. In fact, these Salicaceae are known not only to be adaptable to grow on contaminated areas, but also to be capable of accumulating heavy metals (Pulford and Watson 2003). Moreover, poplar cultural management by means of short rotation coppice cultures is another advantageous feature to be considered in phytoremediation strategies (Rockwood et al. 2004).
Poplars, whose genome has been entirely sequenced (Tuskan et al. 2006), present also the advantage of being generally easy to propagate and, showing fast growth and high biomass production, are therefore useful for short-rotation harvest. In addition, they can be propagated by in vitro techniques and are conveniently exploited in genetic engineering (Confalonieri et al. 2003).
For instance, transgenic white poplar has been obtained expressing a gene for a metallothionein-like protein, thus enhancing tolerance to CuCl2 in in vitro culture (Balestrazzi et al. 2009); transgenic yellow poplar overexpressing mer A9 and mer A18 genes, showed increased tolerance to high mercuric concentration, developing higher biomass and higher amount of Hg (0), which evaporates through the cell surface, compared with non-transformed plants (Che et al. 2003). Anyhow, that evidence of gene transfer and expression in poplar is just related to sterile culture environment.
Some studies have reported differences among poplar clones in heavy metal accumulation (Laureysens et al. 2004) and in the partitioning of elements within the tree organs (Fischerová et al. 2006; Robinson et al. 2000, 2005; Unterbrunner et al. 2007). So, for improving the possible application of these plant species to phytoextraction techniques, there is the need of investigating variation in metal tolerance, accumulation and translocation among the different poplar clones. To this purpose, we used an in vitro screening as an effective tool to exploit Populus alba L. clonal variability. This kind of study can represent a valuable method for the set-up of phyto-recurrent selection cycles, as proposed by Zalesny et al. (2007), from which clones representing the right combination of high levels of trace element uptake and high productivity can be rapidly individuated and chosen for field tests. An in vitro screening reduces not only the growth period and the treatment time length of the plants but also the space required for the experiments. Moreover, environmental factor variability is also reduced. On the other hand, such screenings suffer from the lack of guarantee that tolerance/accumulation patterns found in that short-term experiments will persist with time and occur also in field. So, data obtained by an in vitro screening always need to be confirmed by field performance trials; anyhow, Watson et al. (2003) and Pulford et al. (2002) pointed out that results obtained in hydroponics and in field experiments have a good agreement.
This study was aimed at evaluating the response of three clones of P. alba L. to different heavy metals, in particular cadmium, zinc and copper, and to the metalloid arsenic in terms of tolerance, accumulation and translocation in an in vitro system. Finding a way to quickly characterise poplar clones for their effectiveness in tolerating and accumulating metals is of fundamental importance in an initial screening for the individuation of the most promising clones for field trials on metal-polluted soils.
2 Materials and methods
2.1 Plant material and in vitro growth conditions
In vitro proliferating microshoots of a commercial and two autochthonous P. alba L. clones, Villafranca (Vil), Fiorentini (Frt) and Querce (Qrc), respectively, were subcultured on Woody Plant Medium (WPM, Lloyd and McCown 1980), added with 2% sucrose, 0.72% agar (B&V, Reggio Emilia, Italy) and 0.2 mg l-1 benzyl adenine, at pH 5.5, in 100-ml baby-food glass jars (Sigma-Aldrich, Milano, Italy) sterilised by autoclaving at 121°C and 108 kPa for 20 min. Culturing microshoots originated from axillary bud proliferation of shoot tips collected from adult trees of the mentioned clones, according to Confalonieri et al. (2003).
The aseptic cultures were incubated in a growth chamber at 23 ± 1°C with a 16-h photoperiod (40 μE m-2 s-1) and routinely subcultured (approximately every 4 weeks). Microshoots were finally transferred to phytohormone-free WPM medium for 4 weeks before metal treatments to remove the cytokinin carry-over effect and enhance shoot elongation.
2.2 Metal treatments
Well-developed shoots were cut at uniform size (two internodes; 1.5-cm tall) and transferred to phytohormone-free WPM medium containing a series of Na2HAsO4 (0, 5, 50 and 250 μM), CuSO4 (0, 5, 50 and 250 μM), CdSO4 (0, 5, 50 and 250 μM) and ZnSO4 (0, 250, 1,000 and 2,000 μM) concentrations; the other medium composition and conditions being the same as previously described. Twenty plants per clone per concentration (four glass jars per treatment per clone containing five shoots each) were used in the experiment. Other 20 rootless shoots per clone were dried at 80°C for 24 h and then weighed to estimate a mean value of the dry biomass at the beginning of the experiment.
2.3 Determination of metal toxicity, metal concentration and content
After 15 days of treatment, plantlets were gently removed from the medium and roots were carefully washed. In the case of heavy metal-treated plants, roots were also desorbed with 10 mM CaCl2 solution for 10 min to remove the adhering metals from the cell walls.
Plantlets were separated into shoots and roots, dried at 80°C for 24 h and then weighed. The dry biomass production (dry biomass at the end of the experiment minus dry biomass at the beginning of the experiment previously estimated) was used as a measurement of the metal toxic effects (Baker and Walker 1989). To compare clones, we calculated a tolerance index as the ratio of root or shoot dry biomass production on metal medium to root or shoot dry biomass production on metal-free medium, thus obtaining for each metal three series of tolerance indexes for the three clones.
Roots and shoots of plantlets exposed to CuSO4, CdSO4 and ZnSO4 were mineralised by wet ashing on hot plate (100–300°C) in plugged beakers with 65% HNO3 and 60% HClO4 (5:2, v/v). Instead, root and shoots of As-exposed plantlets were digested in 10 ml 65% HNO3 in closed glass tubes at room temperature because of metal volatility. Reference material was included in the analysis.
Metal concentrations were determined by inductively coupled plasma optical emission spectroscopy (ICP-OES, Optima 2000 DV, Perkin–Elmer Instruments). Metal contents were calculated as the product between mean dry biomass of plantlets and mean metal concentration in each plant.
Data were rearranged in graphs using the SigmaPlot 8.0 programme (SPSS, Chicago, IL).
2.4 Statistical analysis
All treatments were performed in quintuplicate and repeated in at least four independent experiments.
Statistical analysis was carried out with one-way and two-way analysis of variance (ANOVA) using the statistical programme Statistica 6.0 (StatSoft, 2003). A posterior comparison of individual means was performed using Tukey's test. Clones and elements were considered as independent variables. Means were considered different at probability values of P < 0.05.
3 Results
3.1 Biomass production and tolerance index
Root and shoot dry biomass in P. alba clones in presence of increasing metal concentrations is reported in Tables 1 and 2.
In all clones, a significant negative effect of metal treatment on root dry biomass was shown only at the highest concentration for arsenic and zinc treatments.
The reduction in dry biomass due to the presence of cadmium in the culture medium was always significant in the Vil clone, whereas no effect was detected in the other two clones.
The clones displayed a very strong root growth inhibition at the highest copper concentrations, showing roots so reduced to be excluded from the analysis, while the lower concentrations had no significant effect.
The highest values of root dry biomass were always significantly shown by the Vil clone, except in the case of cadmium treatment.
In all clones, roots were significantly more affected by metal toxicity than shoots. In fact, even if a decrease in the weight of shoot dry biomass was shown for some metal treatments, it was never significant in this plant organ.
As in roots, in shoots, the highest values of dry biomass were also shown by the Vil clone, even if only for copper and zinc treatment and significantly only in respect to the Qrc clone.
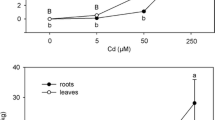
Expression of the tolerance to metal treatments as tolerance indexes (Fig. 1) enabled a comparison of clones. Factorial ANOVA demonstrated that the three clones of P. alba showed significant differences in root dry biomass only in their responses to cadmium treatments. For shoots, no significant difference was detected (data not shown).

Metal tolerance indexes of the three P. alba clones. Letters above the histograms, when present, indicate the significant differences among clones according to the Tukey's test
3.2 Metal accumulation in roots and shoots
Although in all plants, the metal concentration in the tissues increased with the external metal concentration, and metal concentrations were generally higher in roots than in shoots, the three clones differed significantly in root and shoot metal accumulation (Fig. 2).

Root (solid line) and shoot (dashed line) metal accumulation (microgrammes per gramme dry weight) of the three clones of P. alba (mean ± standard error) after exposure to increasing metal concentrations
Root arsenic concentrations were significantly higher in Qrc clone as compared with the other ones at the highest concentration used (P < 0.05). Arsenic accumulation in shoots was not significantly different in the three clones.
As for cadmium accumulation, among the clones Frt showed the lowest cadmium values in both roots (P < 0.05) and shoots (P < 0.05).
Copper concentrations in roots were not measurable at the highest concentration used for the absence of rooting. At the other concentrations, copper accumulation was generally higher in the Frt clones (P < 0.05). In shoots, for the copper concentrations that allowed the root to grow, copper concentrations were not significantly different among the clones. At 250 μM CuSO4, the Vil clone showed statistically significant higher values of copper accumulation in shoot, but this datum will be excluded from the discussion due to the too much higher toxicity of this concentration.
Zinc concentrations were more homogeneous among the clones; the only statistically significant difference was that the Vil clone accumulated this metal to a lesser extent in the roots in respect to the other clones (P < 0.01).
Within each clone, linear correlation between metal concentration in root and metal concentration in shoot was highly significant (0.946 < r < 0.999, at least P < 0.05, Table 3). This correlation was not calculated for Cu as it prevented sufficient root growth at the higher concentration used. As the shoot/root metal concentration ratio never changed in an exposure-dependent way for any of the clones at the metal concentrations used (data not shown), the angular coefficient of this regression line was used to estimate the shoot/root metal concentration ratio itself. This coefficient showed significant differences among the clones for the different metals, even though its range of variation was only from 0.17 (Qrc for arsenic) to 0.89 (Vil for zinc; Table 3). The highest coefficients were shown for zinc and cadmium, the Vil clone displaying the highest, while the lower coefficient was for arsenic, without any significant difference among the clones. Within each clone, the angular coefficient for arsenic was always significantly lower than the other (P < 0.05); the one for zinc was significantly higher only in Vil clone (P < 0.05).
3.3 Metal content
Root and shoot metal content in P. alba clones in the presence of metal treatments is reported in Tables 4 and 5. In all the clones, metal content was always higher in the shoots than in the roots, except for the lowest arsenic concentrations used.
The presence of the metal in the culture medium significantly resulted in an increased root metal content for all the metals, but this increase was never proportional to metal treatment. Among the clones, the Vil clone showed the significantly highest contents for arsenic, copper and zinc, whereas in the case of cadmium the Qrc clone displayed a significantly higher root content for the highest concentration used.
Shoot metal content varied significantly both intra and inter clones. For all the metals tested, shoot content increased proportionally to metal treatment. The significantly highest values of metal content were shown by Vil clone for all the metals.
4 Discussion
The success of phytoremediation techniques is indeed based on the availability of properly selected plant material. Among the species indicated as potential candidates for the remediation of contaminated soil, poplars play an important role, due to their growth, genetic and cultural characteristics (McGrath et al. 2001; Pulford and Watson 2003; Vassilev et al. 2004). In this work, three clones of P. alba were studied to evaluate their patterns of variation in metal tolerance, accumulation and translocation, as these fundamental features should form the criteria to be followed in screening plants for the recovery of polluted soils.
A plant, to be utilised in phytoextraction, must have the essential requisite of metal tolerance, to guarantee the defence of the major physiological and metabolic processes. Nonetheless, this feature should be the result of a combination of metal uptake and reduction of harmful effects and not be simply due to metal exclusion (Krämer 2005; Vangronsveld et al. 2009). Root tolerance means the preservation of the selective property of the cell membrane and so represents the first step in metal uptake and loading into the xylem vessels (Zacchini et al. 2009).
In the present work, the root system of poplar clones, analysed by mean of dry biomass production per plant, showed some differences in metal tolerance. Generally, all the metals, except cadmium for Qrc and Frt clones, showed a significant toxic effect on root dry biomass production at some or all the concentrations used. Comparing the clones among them through the calculation of the tolerance index, significant variation was shown only in respect to cadmium, the Qrc and Frt clones being the most tolerant for the concentrations used. Our findings are in accordance with previous investigations and confirm poplar variability in metal tolerance (see for example Zacchini et al. 2009; and Dos Santos Utmazian et al. 2007).
In P. alba clones, the root responded more sensitively than the shoot to toxic metal concentrations, and the reduction in shoot biomass production was never significant for all the clones and all the metals for the concentration used.
The highest biomass values were generally showed by the Vil clone, confirming the well-known ability of fast growth of this commercial clone. This feature was present either with or without metal treatment, suggesting that even if this clone was generally the most sensitive, the reduction in its growth led to a biomass production that was anyway higher than the autochthonous, and more tolerant, clones.
Acclimation of trees to metal stress has been studied using a variety of indices, such as cell suspension cultures and callus cultures, but seedling growth is the most frequently used index, regardless of their greater sensitivity to unfavourable conditions than mature trees (Pulford and Watson 2003). In fact, also in the present work, this index was able to find out variation in metal tolerance in the studied clones.
The accumulated metal concentration in roots and shoots was affected by the external concentration. In shoots, metal concentration widely exceeded the leaf approximate metal concentrations thought to be toxic (Kabata-Pendias and Pendias 2001). Considering the non-significant effect of metal treatments on shoot growth for the concentrations used, data showed that poplars can accumulate high levels of these metals with no adverse effect on their biomass production and matched results already obtained in field (Laureysens et al. 2004).
Metal concentration in the roots was always higher than in the shoots, suggesting for our P. alba clones and the studied elements a metal exclusion strategy from stems and reproductive tissue by retaining the metal in the roots, thus avoiding its toxicity. Similar results on metal partitioning in hydroponics were found by Zacchini et al. (2009) and Dos Santos Utmazian et al. (2007). Trees can differ greatly in their metal accumulation pattern (Pulford and Watson 2003) but a preferential root allocation of metals remains a widespread behaviour to face metal toxicity in the majority of the plants.
Metal accumulation was variable among clones in root, showing the Qrc clone the highest values, except in the case of copper. In shoot, variation was detected, but it was significant only for cadmium, with Frt clone displaying the lowest concentrations. So, clones with contrasting phenotypes for accumulation-related traits were found, thus suggesting that such screenings deserve to be deepened as they can provide real opportunities for improving both phytostabilization and phytoextraction techniques.
Only in the case of cadmium did the clones with the highest root concentration also show the highest shoot concentration and a clearly correlated pattern for root and shoot metal accumulation.
P. alba clones did not show any tendency to lower the metal shoot/root concentration ratio as the metal concentration in the medium increased. We can speculate that, in P. alba, the lack of this behaviour, generally thought to represent a defence strategy against the toxic effect of the metal, could be due to the fact that, with the external metal concentrations selected for this study, shoot metal concentrations did not reach levels toxic enough to trigger such a response. Among the populations, significant variation was found in this ratio, identified as a fundamental trait for the plant suitable for phytoextraction (Vangronsveld et al. 2009), showing the Vil clone significantly higher values for cadmium and zinc although the concentrations of heavy metals used in this study were different. So, data suggested that each clone can be characterised by a different efficiency in root-to-shoot transport that deserve to be investigated, as variability in shoot/root concentration ratio could contribute to the generation of the differences found in the levels of metal accumulation in the shoot. Anyway, the lack of any relation between arsenic and zinc shoot concentration and root concentration suggests that, among P. alba clones, the higher shoot concentrations are due to other feature than higher shoot/root concentration ratios for these metals.
Despite the higher root metal concentration, metal content was generally higher in the shoots than in the roots in all the clones. This result depends on the huge difference in both biomass production and metal sensitivity between the two organs and is one of the reasons why not only metal hyperaccumulators but also high biomass-producing excluder plants can be effectively used in phytoextraction techniques (Vangronsveld et al. 2009).
Also, metal content showed significant variation among the clones. In root, metal content did not increase proportionally to metal medium concentration probably because of the severe effect on root growth imposed by the metal themselves, whereas, in shoot, a proportional increase was found and, in fact, this organ showed a higher metal tolerance, probably due to the lower metal concentration accumulated. Generally, and always in the shoot, the Vil clones showed the highest values of content for all the metals tested. So, even if the plant metal concentration was rarely the highest, this clone showed the highest metal content, suggesting the biomass production as the key factor in evaluating the phytoextraction capacity of P. alba clones for the metals studied.
Finally, variation was present at a more significant degree in metal accumulation and content than in tolerance, so this is another reason to carry on in vitro screening even if different clones seem to respond in the same way to metal stress in the terms of biomass production.
5 Conclusions
The tree long generation time acts to prevent a fast selection of metal-tolerant genotypes, the generation of which is random or induced by the pollutant (Dickinson et al. 1991). Nonetheless, their basal tolerance mechanisms may allow them to cope with higher metal concentrations than agricultural crops (Riddell-Black 1993) and to survive in contaminated soil, although usually with a reduced growth rate (Dickinson et al. 1992). In this context, the present data demonstrated that in vitro cultures of cuttings really represent a valuable way of assessing the ability of different poplar clones, to take up, tolerate and survive metal stress, thus representing an initial effective screening for the individuation of the most promising clones for field trials on metal-polluted soils. In fact, for the metal concentration used in the experiments, our clones of P. alba showed variation in metal tolerance, metal accumulation and content that deserve to be studied and exploited for the selection of the more suitable tool for phytoremediation purposes. Nevertheless, studies on the effect of the synergistic presence of different metals on clone behaviour and studies proving if cuttings accurately reflect the responses of older trees are still needed.
References
Balestrazzi A, Botti S, Zelasco S, Biondi S, Franchin C, Calligari P (2009) Expression of the PsMTA1 gene in white poplar engineered with the MAT system is associated with heavy metal tolerance and protection against 8-hydroxy-2′-deoxyguanosine mediated-DNA damage. Plant Cell Rep 28:1179–1192
Baker AJM, Walker PL (1989) Physiological responses of plants to heavy metals and the quantification of tolerance and toxicity. Chem Speciat Bioavailab 1:7–17
Che D, Meagher RB, Heaton ACP, Lima A, Rugh CL, Merkle SA (2003) Expression of mercuric ion reductase in Eastern cottonwood (Populus deltoides) confers mercuric ion reduction and resistance. Plant Biotechnol J 1:311
Confalonieri M, Balestrazzi A, Bisoffi S, Carbonera D (2003) In vitro culture and genetic engineering of Populus spp.: synergy for forest tree improvement. Plant Cell Tissue Organ Cult 72(2):109–138
Dickinson NM, Turner AP, Lepp NW (1991) How do trees and other long-lived plants survive in polluted environments? Funct Ecol (Print) 5:5–11
Dickinson NM, Turner AP, Watmough SA, Lepp NW (1992) Acclimation of trees to pollution stress: cellular metal tolerance traits. Ann Bot 70:569–572
Dickinson NM, Baker AJM, Doronila A, Laidlaw S, Reeves RD (2009) Phytoremediation of inorganics: realism and synergies. Int J Phytoremediat 11(2):97–114
Dos Santos Utmazian MN, Wieshammer G, Vega R, Wenzel WW (2007) Hydroponic screening for metal resistance and accumulation of cadmium and zinc in twenty clones of willows and poplars. Environ Pollut 148(1):155–165
Fischerová Z, Tlustoš P, Száková J, Šichorová K (2006) A comparison of phytoremediation capability of selected plant species for given trace elements. Environ Pollut 144:93–100
Kabata-Pendias A, Pendias H (2001) Trace elements in soils and plants. CRC Press, Boca Raton, Florida
Krämer U (2005) Phytoremediation: novel approaches to cleaning up polluted soils. Curr Opin Biotechnol 16:133–141
Laureysens I, Blust R, De Temmerman L, Lemmens C, Ceulemans R (2004) Clonal variation in heavy metal accumulation and biomass production in a poplar coppice culture: I. seasonal variation in leaf, wood and bark concentrations. Environ Pollut 131(3):485–494
Lloyd G, McCown BH (1980) Commercially feasible micropropagation of mountain laurel Kalmia latifolia by use of shoot-tip culture. Comb Proc Int Plant Propag Soc 30:421–427
McGrath SP, Zhao FJ, Lombi E (2001) Plant and rhizosphere processes involved in phytoremediation of metal-contaminated soils. Plant Soil 232:207–214
Mench M, Schwitzguébel JP, Schroeder P, Bert V, Gawronski S, Gupta S (2009) Assessment of successful experiments and limitations of phytotechnologies: contaminant uptake, detoxification and sequestration, and consequences for food safety. Environ Sci Pollut Res 16:876–900
Pulford ID, Watson C (2003) Phytoremediation of heavy metal-contaminated land by trees—a review. Environ Int 29(4):529–540
Pulford ID, Riddell-Black D, Stewart C (2002) Heavy metal uptake by willow clones from sewage sludge-treated soil: the potential for phytoremediation. Int J Phytoremediat 4(1):59–72
Riddell-Black D (1993) A review of the potential for the use of trees in the rehabilitation of contaminated land. WRc Report CO 3467, Water Research Centre, Medmenham.
Robinson BH, Mills TM, Petit D, Fung LE, Green SR, Clothier BE (2000) Natural and induced cadmium accumulation in poplar and willow: implications for phytoremediation. Plant Soil 227(1–2):301–306
Robinson BH, Mills TM, Green SR, Chancerel B, Clothier BE, Fung LE, Hurst S, McIvor I (2005) Trace element accumulation by poplars and willows used for stock fodder. N Z J Agric Res 48(4):489–497
Rockwood DL, Naidu CV, Carter DR, Rahmani M, Spriggs TA, Lin C, Alker GR, Isebrands JG, Segrest SA (2004) Short-rotation woody crops and phytoremediation. Opportunities for agroforestry? Agrofor Syst 61(1):51–63
Tuskan GA, DiFazio S, Jansson S, Bohlmann J, Grigoriev I, Hellsten U et al (2006) The Genome of Black Cottonwood, Populus trichocarpa (Torr. & Gray). Science 131:1596–1604
Unterbrunner R, Puschenreiter M, Sommer P, Wieshammer G, Tlustoš P, Zupan M, Wenzel WW (2007) Heavy metal accumulation in tree growing on contaminated sites in Central Europe. Environ Pollut 148(1):107–114
Vangronsveld J, Herzig R, Weyens N, Boulet J, Adriaensen K, Ruttens A, Thewys T et al (2009) Phytoremediation of contaminated soils and groundwater: lessons from the field. Environ Sci Pollut Res 16:765–794
Van Nevel L, Mertens J, Oorts K, Verheyen K (2007) Phytoextraction of metals from soils: how far from practice? Environ Pollut 150:34–40
Vassilev A, Schwitzguébel JP, Thewys T, van der Lelie D, Vangronsveld J (2004) The use of plants for remediation of metal contaminated soils. Scientific World J 4:9–34
Watson C, Pulford ID, Riddell-Black D (2003) Screening of willow species for resistance to heavy metals: comparison of performance in a hydroponics system and field trials. Int J Phytoremediat 5(4):351–365
Zacchini M, Pietrini F, Scarascia Mugnozza G, Iori V, Pietrosanti L, Massacci A (2009) Metal tolerance, accumulation and translocation in poplar and willow clones treated with cadmium in hydroponics. Water Air Soil Pollut 197:23–34
Zalesny JA, Zalesny RS, Wiese AH, Hall RB (2007) Choosing tree genotypes for phytoremediation of landfill leachate using phyto-recurrent selection. Int J Phytorem 9(6):513–530
Acknowledgements
This study was funded by the CNR (Istituto di Genetica Vegetale) and the University of Firenze (Fondi di Ateneo 2007–2008).
Author information
Authors and Affiliations
Corresponding author
Additional information
Responsible editor: Elena Maestri
Rights and permissions
About this article
Cite this article
Di Lonardo, S., Capuana, M., Arnetoli, M. et al. Exploring the metal phytoremediation potential of three Populus alba L. clones using an in vitro screening. Environ Sci Pollut Res 18, 82–90 (2011). https://doi.org/10.1007/s11356-010-0354-7
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11356-010-0354-7