
Abstract
Background, aim, and scope
Organisms use chemical cues in their surrounding, so-called infochemicals, as important source of information about their biotic and abiotic environment. The scope of this work is to transfer the knowledge on infochemicals obtained in chemical ecology into ecotoxicology, compare the observations with ecotoxicological standard tests, with other sublethal effects, and deduce consequences for the legal situation of environmental chemicals.
Main features
General principles were elaborated from the compiled information from literature on the structures and roles of natural infochemicals. The experiences gained in chemical ecology and in ecotoxicology led to the discovery of the infochemical effect: Anthropogenic substances can influence the chemical communication of environmental organisms. This finding is supported by a close look at fragrances and other common anthropogenic substances in the environment.
Results
Increasing scientific knowledge shows how complex the chemical communication of environmental organisms is. Infochemicals are released by senders and detected by receivers. The relevant concentrations of infochemicals are very low, usually in the nano- to micromolar range and they do not seem to have common structural features. Knowledge about natural infochemicals is still poor and not consistent. The chemical cues fluctuate specifically in time and space resulting in dynamic response patterns in the ecosystem. Organisms can react to infochemicals in very specific ways by behavioral, morphological, or physiological responses; activities that are relevant for their survival as vital reactions such as flight, food uptake, or mating are affected. Anthropogenic substances at minor concentrations can interfere in the complex chemical communication web of infochemicals, possibly leading to increased vulnerability of populations.
Discussion
The findings show clearly that the actual description of the interplay of organisms in the ecosystem is still very simplified and we are far from understanding the interactions completely. Anthropogenic discharges may play a role on the chemical communication and, hence, on the behavior and interactions of organisms in the ecosystem. The description of the infochemical effect opens a new chapter in ecotoxicology. It is a challenge to develop a suitable test system for the infochemical effect with the knowledge of the multitude of possible reactions and of the high specificity of infochemicals. Problems during the performance and evaluation of standard tests might be related to reactions due to infochemicals in the test systems which have not been considered so far.
Conclusions
The roles of anthropogenic infochemicals in the environment and the role of natural infochemicals in laboratory tests have been underestimated up to now.
Recommendations and perspectives
The discrepancy between the biological relevance and the lack of data about infochemicals in the environment reveals the necessity of further research. According to the actual findings, infochemicals are so decisive for the interactions in the ecosystem that they should not be neglected in ecotoxicology. The discovery of the infochemical effect is comparable to the detection that environmental substances can act as hormones. Sublethal effects with impacts on the ecosystem, such as the infochemical effect, will receive higher appraisal in the ecotoxicology of the future. It needs to be clarified to which extent anthropogenic discharges disturb the natural chemical communication web. A systematic analysis of this very complex field will be needed to know whether a new ecotoxicological endpoint, the infochemical effect, will have to be taken up in the standard repertoire. The knowledge on infochemicals might require some adjustments of the legal framework on environmental chemicals in future. Looking closer at the infochemical effect will lead to a new understanding of the complexity of environmental communities.
Similar content being viewed by others


Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
1 Background, aim, and scope
Fathead minnows, Pimephales promelas, do not need to see their predators to know that pikes are nearby, to know how big they are, how many they are, whether they are hungry, and whether they feed on fathead minnows. They are able to smell all this information very precisely (Chivers and Mirza 2001; Ferrari et al. 2006a, b; Kusch et al. 2004). In the last years, many new findings showed the complexity of chemical sensing (e.g., Atzmüller and Grammer 2000; Brönmark and Hansson 2000; Burks and Lodge 2002; Chivers and Smith 1998; Dodson et al. 1994; Wyatt 2003; Zimmer and Butman 2000). Chemical cues in the water or the air inform an organism about its biotic or abiotic environment. These substances can be, for example, sex pheromones, alarm substances, fear substances, pollinator attractants, or substances that convey other messages. One chemical may stand for more than one message; therefore, the general term infochemicals is preferred here for the multitude of chemical cues. Some natural infochemicals that were identified in chemical ecology are identical with substances used and discharged by man, such as fragrance ingredients (Klaschka and Kolossa-Gehring 2007), and might interfere with the natural chemical communication. This so-called infochemical effect (Klaschka 2008) describes that anthropogenic substances can influence the chemical perception by an organism. The infochemical effect can have lethal consequences for the individuum, even if the respective substance is not toxic, for example, if the organism does not recognize its predator and does not seek shelter in time. Most consequences of the infochemical effect might be of a sublethal quality on the level of the single organism, but they can have a major significance for the population or the ecosystem if, for example, the organism does not recognize the odor of the sexual partner and does not mate or if it does not recognize the correct food, eats less, loses in strength, and shows a reduced reproduction rate. So far, it is not clear whether and to what extent these effects play a role in the natural ecosystem.
Organisms meet a plethora of environmental stimuli every day which they interpret and use for appropriate responses. The chemical environment plays an important role for the fine tuning of the activities that ensure survival. Chemical communication has many advantages compared to visual or acoustic communication strategies. It is independent of light and applicable at night, at the bottom of lakes or the sea, in groundwater, in turbid water, in water habitats with hiding places such as plants or rocks, or in caves. The chemical communication is always effective as most animals cannot shut their noses or inactivate their chemoreceptors. The stimulus can be actively reduced by going away from the emission source or it can be passively reduced if organisms that are exposed to constant concentrations of an infochemical become insensitive due to adaptation. The signals disappear with diffusion or degradation of the compounds. Chemical signals are well suited for medium time frames: They last longer than sounds, vibrations, or short-lived visual cues such as movements, but they do not last as long as most morphological changes. Chemical signals are also well suited for the medium spatial range: They do not reach as far as penetrating sounds but reach farther than touch or taste. In contrast to light or sound, there is no linear scale such as the spectra of wavelengths or frequencies; instead, there is an unlimited number of odor qualities, depending on the number of chemicals and of the olfactory receptors in the receiver. One reason for the universality of chemical communication is the fact that chemicals can start the biochemical signal transduction chain directly without the need of transforming a physical signal into a biochemical one.
The understanding of the chemical communication gives new insights in ecotoxicology and reconfirms the complexity of ecosystemary interactions. Ecotoxicology as it is now is not perfect. The lessons learned in the past with substances showing endocrine effects or with pharmaceuticals suggest that new aspects and eventually new endpoints need to be considered in future. The test systems presently used are simple model systems for the ecosystem which proved to be useful for the issues and chemicals dealt with so far, but it is necessary to keep in mind that it does not correspond to the real world. The discussion about the infochemical effect helps to relearn this lesson.
2 Results
2.1 Natural infochemicals
2.1.1 Senders emit infochemicals, receivers can perceive them
It is important to note that the definition of infochemicals does not use structural features of the chemical, but it is based on the capacity of a receiver to recognize the substance. The perception renders a substance an infochemical. A substance may be an infochemical for only one or a few species not for others. Infochemicals can be intraspecific if the sender and receiver belong to the same species (i.e., pheromones) or interspecific if the sender and receiver belong to different species (i.e., semiochemicals). Infochemicals are present in the surrounding of the receiver and are therefore recognized by chemoperception of distant (long- to medium-range) signals that is in most organisms the sense of smell.
The consecutive steps in the reaction chain between sender and receiver, respectively odor recognition, were analyzed in the last years in more detail (e.g., Buck and Axel 1991; Freitag et al. 1998; Krieger and Breer 1999; Young and Trask 2002; Zarzo 2007; Zhao and Firestein 1999): The sender emits specific infochemicals under specific conditions, for example, an adult female emits sexual pheromones. The infochemicals diffuse in the air or water and some molecules come eventually into contact with the olfactory epithelium of the receiver where they bind to specific odorant binding proteins on the cell surface of the olfactory neuron. Binding to the odorant receptor leads to a conformational change of the receptor protein which induces an electrophysiological response that is directly conducted to the brain, usually leading to the perception of an odor and eventually to a reaction of the whole organism. The olfactory cells are primary sensory cells, i.e., olfactory neurons are the only cells that are in direct contact with the outside world and at the same time in immediate contact with the brain as signal processing organ. Specific odorant degrading enzymes can lead to the disappearance of the stimulus (Ferrari et al. 2007). In insects, the odorant forms a complex with a specific odorant binding protein in the antenna that is transported to the specific receptor proteins in the sensory neurons. As described here, three different specific protein species can be involved in the signal transduction: odorant binding proteins, odorant receptor proteins, and odorant degrading enzymes. In insects, the odorant binding proteins are encoded by many genes, e.g., 57 genes were identified for putative odorant binding proteins in Drosophila melanogaster (Field et al. 2007). Vertebrate odorant receptors detect a multitude of diverse odorant molecules. They are encoded by a large gene family belonging to the G-protein coupled transmembrane receptors. In mammals, this is the largest gene family with up to 1,000 genes (Buck and Axel 1991; Zhao and Firestein 1999). They show a relatively low sequence identity but seem to have conserved structural features (Khafizov et al. 2007). The fish receptor gene family is much smaller containing around a hundred genes (Freitag et al. 1998). These large numbers of genes can explain the high specificity of the chemical sensing. Plants are also able to perceive volatile substances with a surprising specificity (Boller 1995; Larcher 1995; Polya 2003).
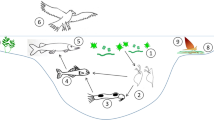
The chemical communication is like a complicated dynamic web. A sender emits usually more than one infochemical at various concentrations and at various times and a receiver perceives many infochemicals simultaneously from many sources. Imagine that environmental organisms are confronted with infochemicals from conspecifics (mates, competitors, kins, offspring), from food/prey, from predators, from other heterospecifics, or from other sources (Fig. 1).

Schematic simplified illustration of the complexity of the infochemical web. Senders can emit various substances that can be infochemicals for various receivers. The receivers can react to a certain infochemical by various reactions. The arrows in the figure give some examples. In nature, there will of course be more than three senders, infochemicals, or receivers each. These interrelationships are much more complex in nature: there can be tritrophic interactions, as explained in the text, or the number of infochemicals can vary. Anthropogenic discharges of substances might interfere in this network. The internal factors can be for example age, size, or feeding status, the external factors can be light, temperature, food concentration, other organisms, or anthropogenic substances
2.1.2 Chemical structures of infochemicals are diverse
The knowledge about the chemical structures of natural infochemicals is still very limited. The infochemicals relevant for animals and unicellular organisms belong to many chemical groups and were identified for example as proteins, peptides or amino acids, carbohydrates, hydrocarbons, aldehydes, carbonic acids, lipids, lactones, steroids, phenolics, terpenes, phosphatidylcholine, or nitrogen-containing compounds (see Klaschka 2008). There is no obvious systematic correlation of a group of organisms and a chemical structure. The knowledge obtained about infochemicals of one group of organisms cannot be transferred to other organisms not even of the same genus. It is interesting that all infochemicals identified so far for ciliates were proteins, but this does not mean that other substances might not be found to be infochemicals for ciliates in future. Some substances are very specifically used only by one species; others are more common and ‘understood’ by various unrelated species. For example, the sexual pheromone of the Asian elephant is the same used by 140 species of moth (Rasmussen et al. 1996). Some of the infochemicals have chiral centers and the stereoisomers are physiologically very different: S-carvone smells aromatic, whereas R-carvone has a minty odor. R-limonene smells like orange and sweet, whereas S-limonene smells like herbs and peppermint (Lahlou 2004). Chemical cues used by terrestrial higher plants and algae are for example oligosaccharides, glycopeptides, peptides, proteins, and other lipophilic substances (Larcher 1995; Boller 1995). Many examples for natural infochemicals are described for terrestrial insects (adults and their reproductive states) stimulated by research on pest control; much less information exists about aquatic organisms.
In most cases, the chemical stimulus is the result of a complex mixture of many substances. For example, the queen pheromone of bumble bees Bombus terrestris has 300 different constituents with 16 physiologically active and identified compounds (Ayasse, personal communication). It was shown in some occasions that the senders can produce such a complex blend of odorants at low expense by a minimum number of enzymes by combinatorial biochemistry. In most studies on natural chemical cues, it is already a big success if at least a few single substances could be identified, knowing that they might be components of a complex mixture. The perception of infochemicals is not simple: Mixtures of several odorants can activate additional neurons compared to the single odorants separately and evoke qualitatively different smells (Zou and Buck 2006).
Figure 2 illustrates in a simplified model the potential origins of infochemicals which explains the multitude of chemical structures listed above. The substances can be originally food ingredients or plant material that might be transformed or used as such (as shown for many insects, Eltz et al. 2005). The chemical cues of predators depend on the prey species they ingested previously (Ferrari et al. 2006a, b). Infochemicals can be tissue components set free after injury. Alarm cues that are released by tissue damage were described for protozoans, flatworms, annelids, arthropods, mollusks, fish, and amphibians (Chivers et al. 1996). In several examples, the chemical cues assigned to a eukaryotic organism turned out to be the emissions of bacteria growing on or in the respective organisms. Degradation products emitted by injured or dead organisms can serve as food attractants. Diet released metabolites and waste are, for example, constituents in feces or urine in dominant male fish or crayfish (Breithaupt and Eger 2002). An induced synthesis is assumed for specific compounds such as specific sexual pheromones. Reactions in the extracellular space, eventually catalyzed by enzymes yielding transformation products, can be assumed for all routes mentioned. The release from the sender was observed as excretion, exocytosis, released cell surface signals, or simple leakage or diffusion in case of damaged or dead tissue. The release of alarm substance from club cells in the skin of fish has been studied by many laboratories.

An overview of some potential origins of infochemicals. The scheme illustrates the fact that infochemicals show various structural features (a.a. amino acids, OBP odorant binding protein, ORP odorant receptor protein, transf. prod. transformation products)
Infochemicals are by definition extracellular substances and mostly, they are odorants. Intracellular toxins, which are detrimental for the predator after ingestion, or antifeedants, which are effective after ingestion, are not considered as infochemicals. Some toxins which are effective also in the extracellular space as shown by bioassays, e.g., for okadaic acid from the red tide dinoflagellate or domoic acid from diatoms (Pohnert 2004), might, however, function also as infochemicals. A substance may be an infochemical for one species, a nutrient for another, waste for a third, and a toxin for a further species. In some cases, it is not clear whether a grazing inhibitory substance is an intracellular toxin or an extracellular infochemical (Turner and Tester 1997). The functions may also be different at various concentrations of a compound. The classification into separate groups is very artificial and does not correspond to the natural world. There is a complicated interplay between nutrients, toxins, and infochemicals, and it does not seem to be adequate to regard these groups as strictly disjunctive (see also Pohnert et al. 2007).
2.1.3 Effective concentrations are low
The natural environment of most aquatic organisms consists of a very dilute watery solution containing chemicals at very low concentrations that inform the receiver about mates, food, prey, predators, and so on. The sender emits very small amounts of infochemicals to minimize the cost of production, to avoid eavesdropping by potential enemies, and because the sender is usually very small compared to the surrounding volume. Environmental organisms are very sensitive and can interpret chemical signals at concentrations in the nanomolar and even lower range (Wolfe 2000). The high sensitivity was, for example, shown for the bile acid taurolithocholic acid where concentrations as low as 10–12 M proved to be recognized by rainbow trout in the facial nerve of the taste buds (Yamashita et al. 2006). Concentrations controlling behavior may be 0.1–5.0% of the LC50 found in standard tests (survival of rainbow trout; Little et al. 1993). The recent improvements of analytical methods make it possible to detect the very low concentrations of infochemicals. Biological responses can even be observed with fractions of the infochemicals obtained by gas chromatography where no peak is to be seen (W.S. Leal, personal communication). A substance can convey qualitatively different messages, depending on the concentration present, as everybody knows the odor quality may change with concentration, e.g., high concentrations of methyl sulfide smell like spoilt eggs, very dilute concentrations like asparagus. Some infochemicals are identical to substances which are present at high concentrations in tissues, such as amino acids. They can still be very specific infochemicals in a concentration window between high tissue concentrations and low background concentrations (Carr 1988; Klaschka 2008).
2.1.4 Receivers can respond in many different ways
It is an enigma how accurately the organisms use the information of the chemical surrounding for appropriate responses. Very many different reaction patterns were described: Examples for developmental reactions were shown in all taxa, such as cell division, growth, mating/conjugation, growth of ovaries, laying eggs, encystment, and diapause induction, hatching of resting stages, aggregation, colony formation, quorum sensing (of bacteria). Morphological changes such as formation of spines, helmets, neckteeth, keels, ridges, increased/decreased size, and change of body proportions were shown for protozoans, cladocerans, rotifers, bryozoans, gastropods, insects, fish, and amphibians. Behavioral reactions were also observed in all taxa, for example, increased/decreased food uptake, enhanced/reduced or directed/undirected mobility, dial vertical migration, phototactic behavior, looking after, and feeding the offspring. Also the production or release of toxins or infochemicals can be the response to an infochemical stimulus. An appropriate response of an organism to an infochemical stimulus can ask for a combination of several single reactions: The comprehensive defense strategy of Daphnia can comprehend diel vertical migration as well as suppressed growth, delayed hatching, or the formation of helmets (e.g., Boersma et al. 1998; Burks et al. 2000; Stibor and Lampert 2000).
The responses to infochemicals can be intraspecific, for example, attract a mate, copulate, recognize, or feed the offspring, inform the conspecifics about the presence of food, about their mood, mark territories, or warn conspecifics. The responses can also be interspecific, for example, recognize a suitable prey, identify and avoid a predator, camouflage invaders (for example parasites of ants), or recognize the correct plant for egg disposal. Infochemicals can be used simultaneously intra- and interspecifically: The alarm substances produced by a prey can be used by conspecifics and also by other prey species if it is hunted by the same predator.
Even for plants, the chemical communication plays important roles: Pollinators are attracted by fragrances; plants that are attacked by caterpillars can attract the exact species of ichneumons that are parasites of the respective aggressors (tritrophic interaction). Allelopathy (plants release substances that reduce or inhibit growth of other plant species) or phytoalexines (substances produced as defence against aggressors in plants) are widespread examples of infochemicals in the plant kingdom (Boller 1995; Larcher 1995; Pichersky 2004).
Each organism is confronted with conflicting environmental visual, acoustic, and chemical information, and it has various options of suitable reactions (e.g., Tomba et al. 2001; Shiojiri et al. 2006). Some reactions are only favorable at certain situations and not at others, e.g., if a prey increases its morphological defenses by formation of spines or helmets as defense against one predator, it increases at the same time its visibility and becomes more vulnerable in the presence of other visually hunting predators (Tollrian and Harvell 1999). Swarming can be a defense reaction against visually hunting predators, e.g., fish predators, as they are confused by a swarm, but swarming can also be a disadvantage for the prey as a swarm is detected more easily and can attract other visually hunting predators, e.g., fish-eating seabirds. The responses to infochemicals are not stereotype but depend on internal factors such as age, sex, conspecifics, feeding state, mood, injuries, learning, or activity state/sleeping/awake. This is not surprising: Humans also react differently to the smell of food if they are hungry or not. Also external factors influence the response pattern such as light, pH, temperature, time of the day, conspecifics, and other organisms, water, or air current (Klaschka 2008; Pollock et al. 2006; Prince et al. 2006). Chemicals in the surrounding medium can influence the perception of odors, as was shown for neuromodulators (Czesnik et al. 2007).
The benefit for the sender does not always seem to outweigh the costly production of infochemicals. Warning by alarm cues does not benefit the sender itself but its conspecifics (Chivers et al. 1996). The question must be posed whether the real functions of these substances were understood sufficiently. For example, fish of the order Ostariophysi emit alarm cues from their epidermal club cells after attack by a predator. These substances could benefit the sender by attracting attention of other predators which would interfere in the predation process and give the prey a chance to escape or by attraction of conspecifics that might support the attacked prey. Surprisingly, substances that were considered as alarm substances were shown to have a predominant role as skin protecting substances against pathogens, parasites, and ultraviolet B radiation whereas the alarm function does not seem to be of first priority (Chivers et al. 2007). The same is true for fragrances and essential oils which have a function in the chemical communication and additional properties, e.g., as defense against herbivores: They can be molluscicidal, induce muscle contractions or are spasmolytic, can affect the central nervous system, interact with lipids in the cell membranes of the cortex, can modify ion permeability of membranes, act on the blood pressure, or act as antioxidants (Lahlou 2004).
Irrespective of the multitude of natural infochemicals and their roles, there are striking similarities in the organization of chemical sensing in the various taxa (Dryer 2000; Krieger and Breer 1999). Infochemicals played comprehensive roles in evolution as illustrated by three examples: Infochemicals were important for sexual isolation leading to the formation of new species. Chemical-induced selection pressure by predators was shown to be transferred to the next generation (Agrawal et al. 1999). Infochemicals were involved in the fine tuning across species like in the coevolution of pollinators and plants.
Synoptical tables with examples of senders/receivers/infochemicals can be found in, e.g., Dodson et al. (1994), Klaschka (2008), Klaschka and Kolossa-Gehring (2007), Larsson and Dodson (1993), or Polya (2003). The examples in this article were selected to show the complexity of the chemical communication by infochemicals. Simple systematic categories do not seem to cover all aspects. Table 1 gives an overview of the multiplicity of the findings about infochemicals described here.
2.2 Anthropogenic infochemicals and the infochemical effect
Some of the man-made chemicals discharged into the environment can interfere with the delicate natural chemical communication system. This is called infochemical effect (Klaschka 2008). Some anthropogenic compounds are identical to substances used in the natural communication systems. This is, for example, the case for many fragrance ingredients, many of which are pollinator attractants in plants or pheromones for animals in nature. Substances released from odoriferous anthropogenic products are perceived by animals just like naturally emitted compounds. For example, Japanese scientists found out that hornets perceive 2-pentanol, used as fragrance ingredients in cosmetics, as alarm pheromone and attack specifically people that applied products containing this compound (Ono et al. 2003). Benzaldehyde, benzoic acid, citral, isopentenylacetate, alpha and beta pinene, or salicylaldehyde are used as fragrance ingredients and were described as infochemicals in natural systems (Klaschka and Kolossa-Gehring 2007). Anthropogenic substances do not have to be identical to natural infochemicals to have an effect. Also new synthetic fragrance molecules are bioactive compounds as they invoke the odor sensation in the human nose. It cannot be excluded that odoriferous molecules that induce a sensation in man induce also a sensation in other organisms. Also pesticides, heavy metals, or other contaminants were shown to interfere with the natural chemical communication system at very low sublethal concentrations (Barry 2000; Hanazato 1999; Lüring and Beckmann 2002; Michels et al. 2000; Moore and Lower 2001; Preston et al. 1999a, b). For example, it has been known for a long time that exposure to cadmium is responsible for an olfactory dysfunction in man. Cadmium is taken up specifically by olfactory sensory neurons in fish and transported in the axon towards the central nervous system. At environmentally relevant concentrations of cadmium fish species are no longer able to produce alarm substances to warn their conspecifics (Blechinger et al. 2007).
On the basis of the compiled information, the main questions arise: Which anthropogenic substances are responsible for the infochemical effect? To what extent do anthropogenic emissions influence the natural communication system? Which organisms are affected? Many facts indicate that the effect might be grave. Brown and Smith (1998) describe an example where infochemicals play a role for the heavy decrease of a population: It is a common observation that hatchery reared trout undergo high mortality when released in the wild. As they could not learn the smell of their predators, they do not show antipredator flight reactions. In other cases, it is not possible at the moment to address the infochemical effect as major cause responsible for a reduced vigor of a population in the environment, because infochemicals can play so many different roles and because the perception and the resulting reactions of organisms are influenced by many external and internal factors (Klaschka 2008). More needs to be known before it might be possible to observe the infochemical effect in nature.
Organisms are flexible. One can imagine that they can use several strategies to cope with disturbances by infochemicals, like the following three possibilities: The mixture of natural infochemicals might be so specific that the organisms will not be confused if a few components are not correct due to anthropogenic influences. Populations might have a surplus of individuals to survive and the losses due to the malfunction of chemical communication might be buffered. The organisms might be able to learn and interpret the changed composition of the chemical environment. A better understanding of infochemicals and of the infochemical effect is needed as prerequirement for finding out whether these strategies are relevant in nature.
3 Recommendations
3.1 Testing the infochemical effect
A systematic analysis of the infochemical effect would be needed to answer the questions posed in the previous paragraph. It will not be easy to define what a systematic analysis would look like. The variety of effects and interactions seems to be unlimited. The development of a standard test protocol for ecotoxicology should be based on the experience gained in chemical ecology. It should be performed with a representative organism, yield reliable dose–effect relationships and should show the relevance for the ecosystem. In principle, the interference of anthropogenic substances on the chemical communication could be measured at each step of the reaction chain described in Section 2.1.1. The following list gives examples for the approaches that are conceivable to identify the role of an infochemical: binding studies to the odorant binding protein or to the odorant receptor binding protein, in situ hybridization and immunohistochemical studies, measurement of electrical signals of the receptor cells in the tissue (electroolfactograms, electroantennograms), measurement of growth of populations by cell density or turbidity, registration of phenotypic changes, behavioral tests (in situ online biomonitoring, use of T-shaped olfactometers, test of avoidance responses), and multispecies tests with observation of community structure and community function. The development of a suitable test design will not be easy as there are a multitude of relevant chemicals, a multitude of relevant receptors, and a multitude of relevant reactions, and it must be expected that the effective concentrations are extremely low. The chemical communication is of utmost importance for the ecosystem and justifies great endeavors to find solutions to these problems. The ideas listed above can be a point of depart for further experimental research.
3.2 Significance for standard tests
Infochemicals present in standard ecotoxicological test systems could play roles which were not considered so far: Contaminating substances unintentionally present at minor concentrations could affect the test organisms by the infochemical effect with the subsequent morphological, physiological, or behavioral responses (e.g., Weber 2001). The phenotypic plasticity of organisms in a test system might be due to infochemicals which had not been considered so far. The symptoms regarded as ‘unspecific’ and attributed to so-called ‘environmental stress’ might be specific reactions to the disturbed chemical communication system. It was described above that the reactions of a certain organism to infochemicals are flexible depending on external and internal factors; this flexibility might explain part of the variability in standard test results.
The consideration of the importance of chemical communication leads to a new understanding of observations that were difficult to understand so far: The qualitative shift of infochemical perceptions at different concentrations with the subsequent consequences for the chemical communication might be regarded as an example for hormesis (BUA 2007). The consideration of infochemicals opens the eyes for a new quality in ecotoxicology and a new level of complexity in standard test systems. This knowledge will help to understand reactions of test organisms which were considered as equivocal before and it will help to beware of misinterpretations.
Computer-based models of ecotoxicological effects are widespread and very useful to save time and money. These models fit best for unspecific effects or for specific effects of substances with known structural features. The fact that some models do not fit with the experimental data might therefore be explained by effects such as the infochemical effect which have not been taken into account so far. The effects of infochemicals are very specific. New models are needed to reflect these effects (Browne et al. 1998; Rossiter 1996).
3.3 Comparison with other sublethal effects
Sublethal effects at low concentrations have received more attention in the last years. It became clear that they can play major roles on the level of populations, communities, or ecosystems. Several mechanisms were described to explain the deleterious sublethal effects observed: endocrine effects, chemosensitizers that impede the multidrug resistance transporters (Luckenbach and Epel 2005; Manzini and Schild 2003; Smital et al. 2004), or specific effects by pharmaceuticals at very low concentrations (Kümmerer 2001; Kümmerer 2004). The infochemical effect should be added in this list. These effects can be interconnected. For example, infochemicals can also be involved in endocrine processes (Moore and Lower 2001; Waring and Moore 1997; Stoka 1999). Infochemicals exhibit similarities with substances showing an endocrine effect, chemosensitizers, or pharmaceuticals in the environment: They act specifically, they are efficient at very low concentrations, and the number of compounds with these effects seems to be large.
There are several explanations why the sublethal effects due to the infochemical effect have not been detected so far: The responses to infochemicals might be observed at much smaller concentrations than the observation of lethality. Changes of the behavior might also be much faster than other endpoints, e.g., the swimming activity of rainbow trout was influenced after 96 h whereas the growth was only affected after 30 days (Little et al. 1990; Little et al. 1993). Behavioral effects, e.g., as response to infochemicals, are among the most sensitive responses to chemical stress (Gerhardt and de Bisthoven 1995). In addition, usually single species are tested, which does not allow to detect effects of interspecies communication.
3.4 Legal consequences
In the actual version of the European chemicals regulation Registration, Evaluation, and Authorisation of Chemicals (REACh; EC 2006), there are several procedures that stand against a standardized closer consideration of infochemicals. For example, according to Annex V of REACh, natural substances do not need to be registered if they did not undergo a chemical reaction or were not classified or labeled according to Directive 67/548/EWG (EC 1967). Many infochemicals, such as many fragrances, are identical to or are natural substances and for many of them, the data basis is not sufficient to decide whether they are to be classified and labeled or not. According to REACh, substances with a low exposure need not be tested further. Infochemicals are effective at very low concentrations. Fragrances are used at low concentrations because they are active at low concentrations. The ratio between effect and exposure—and not exposure alone—indicates whether a substance is environmentally relevant as elaborated in the European Technical Guidance Document for risk assessment (EC 2003). At due time and with sufficient knowledge, the infochemical effect should be integrated in the European environmental risk assessment scheme, like the endocrine or other specific effects. According to REACh, substances which are chemically related can be assessed in groups. As many infochemicals such as fragrance ingredients are closely related, the ecotoxicological data for some chemicals might be extrapolated to others. This could be critical for the effect as infochemicals, as molecules with very similar structures can induce very different odor sensations and play different roles in the chemical communication.
The knowledge about infochemicals might also affect the regulations in the European Water Framework, as biomonitoring might become a suitable method to observe the infochemical effect. The knowledge on infochemicals could also lead to an improvement of products and reduce the impact on the environment.
The discussion on the infochemical effect implies one main conclusion for environmental legislation: The discharge of chemicals should be minimized as the impacts they may have are far from being understood. If substances are so potent like infochemicals, more should be known about their effects before they are released into the environment. This is a very strong support of the precautionary principle.
4 Perspectives
Amazing facts have been elucidated about natural infochemicals in chemical ecology in the last years. Chemical communication plays a basic role for wildlife for all taxonomic groups for their reproductive and social behavior, their defense and orientation. Environmental organisms are simultaneously exposed to natural infochemicals from different sources and to anthropogenic compounds that can influence their chemical communication. The consideration of infochemicals in ecotoxicological tests opens up a new horizon. It leads to a new understanding of dose–effect relationships and sublethal effects. Some authors understand the relevance of these implications: As “... such chemicals can interfere with ... chemical communication at sublethal concentrations, there might be a large group of hitherto unknown compounds that could disrupt aquatic community structure and food webs in both lakes and oceans.” (Pohnert et al. 2007). Much more research is needed to elucidate systematically the role of infochemicals and transfer the knowledge to ecotoxicology. Appropriate test methods need to be developed for a realistic assessment in nature. Looking closer at the infochemical effect shows how complex the environment is and how little is known so far. The infochemical effect may be an important effect in ecotoxicology that has been ignored so far.
References
Agrawal AA, Laforsch C, Tollrian R (1999) Transgenerational induction of defenses in animals and plants. Nature 401:60–63
Atzmüller M, Grammer K (2000) Biologie des Geruchs: Die Bedeutung von Pheromonen für Verhalten und Reproduktion. Speculum 18(1):12–18
Barry MJ (2000) Effects of endosulfan on Chaoborus-induced life-history shifts and morphological defenses in Daphnia pulex. J Plankton Res 22:1705–1718
Blechinger SR, Matz C, Kusch RC, Chivers DP, Krone PH (2007) Brief embryonic cadmium exposure induces a stress response and cell death in the developing olfactory system followed by long-term olfactory deficits in juvenile zebrafish. Toxicol Appl Pharm 224:72–80
Boller T (1995) Chemoperception of microbial signals in plant cells. Annu Rev Plant Physiol Plant Mol Biol 46:189–214
Boersma M, Spaak P, de Meester L (1998) Predator-mediated plasticity in morphology, life history, and behavior in Daphnia—the uncoupling of responses. Am Nat 152:237–248
Breithaupt T, Eger P (2002) Urine makes the difference: chemical communication in fighting crayfish made visible. J Exp Biol 205:1221–1231
Brönmark C, Hansson LA (2000) Chemical communication in aquatic systems—an introduction. Oikos 88:103–109
Brown GE, Smith RJF (1998) Acquired predator recognition in juvenile rainbow trout (Oncorhynchus mykiss): conditioning hatchery-reared fish to recognize chemical cues of a predator. Can J Fish Aquat Sci 55:611–617
Browne KA, Tamburri MN, Zimmer-Faust RK (1998) Modelling quantitative structure-activity relationships between animal behaviour and environmental signal molecules. J Exp Biol 201:245–258
BUA (2007) Beratergremium für Altstoffe. Hormesis in der Ökotoxikologie. BUA Bericht 267, Hirzel, Stuttgart
Buck L, Axel R (1991) A novel multigene family may encode odorant receptors: a molecular basis for odour recognition. Cell 65:175–187
Burks RL, Lodge DM (2002) Cued in: advances and opportunities in freshwater chemical ecology. J Chem Ecol 28:1901–1917
Burks RL, Jeppesen E, Lodge DM (2000) Macrophyte and fish chemicals suppress Daphnia growth and alter life-history traits. Oikos 88:139–147
Carr WE (1988) The molecular nature of chemical stimuli in the aquatic environment. In: Atema J, Fay RR, Popper AN, Tavolga WN (eds) Sensory biology of aquatic animals. Springer, Berlin, pp 3–27
Chivers DP, Smith RJF (1998) Chemical alarm signalling in aquatic predator–prey systems—a review and prospectus. Écoscience 5:338–352
Chivers DP, Mirza RS (2001) Predator diet cues and the assessment of predation risk by aquatic vertebrates—a review and prospectus. In: Marchlewska-Koj A, Lepri JJ, Müller-Schwarze D (eds), Chemical signals in vertebrates, vol 9. Plenum, New York, pp 277–284
Chivers DP, Brown GE, Smith RJF (1996) Evolution of chemical alarm signals—attracting predators benefits alarm signal senders. Am Nat 148:649–659
Chivers DP, Wisenden BD, Hindman CJ, Michalak TA, Kusch RC, Kaminskyj SGW, Jack KL, Ferrari MCO, Pollock RJ, Halbgewachs CF, Pollock MS, Alemadi S, James CT, Savaloja RK, Goater CP, Corwin A, Mirza RS, Kiesecker JM, Brown GE, Adrian JC Jr, Krone PH, Blaustein AR, Mathis A (2007) Epidermal ‘alarm substance’ cells of fishes are maintained by non-alarm functions: possible defence against pathogens, parasites and UVB radiation. Proc R Soc Lond B 274:2611–2619
Czesnik D, Schild D, Kuduz J, Manzini I (2007) Cannabinoid action in the olfactory epithelium. PNAS 104(8):2967–2972
Dodson SI, Crowl TA, Peckarsky BL, Kats LB, Covich AP, Culp JM (1994) Non-visual communication in freshwater benthos—an overview. J North Am Benthol Soc 13:268–282
Dryer L (2000) Evolution of odorant receptors. Bioessays 22:803–810
EC (1967) Council Directive 67/548/EEC of 27 June 1967 on the approximation of laws, regulations and administrative provisions relating to the classification, packaging and labelling of dangerous substances. Official Journal P 196, 16/8/1967, as amended by Commission Directive 2001/59/EC of 6 August 2001 adapting to technical progress for the 28th time Council Directive 67/548/EEC on the approximation of the laws, regulations and administrative provisions relating to the classification, packaging and labelling of dangerous substances. Official Journal L 225, 12/8/2001
EC (2003) Technical guidance document on risk assessment in support of commission directive 93/67/EEC on risk assessment for new notified substances, Commission Regulation (EC) No 1488/94 on risk assessment for existing substances and directive 98/8/EC of the European Parliament and of the Council concerning the placing of biocidal products on the market, Parts I, II and IV. European Communities, EUR 20418 EN/1
EC (2006) Regulation (EC) No 1907/2006 of the European Parliament and of the Council concerning the Registration, Evaluation, Authorisation and Restriction of Chemicals (REACh)
Eltz T, Sager A, Lunau K (2005) Juggling with volatiles: exposure of perfumes by displaying male orchid bees. J Comp Physiol A 191:575–581
Ferrari MCO, Messier F, Chivers DP (2006a) The nose knows: minnows determine predator proximity and density through detection of predator odours. Anim Behav 72:927–932
Ferrari MCO, Kapitania-Kwok T, Chivers DP (2006b) The role of learning in the development of threat-sensitive predator avoidance: the use of predator cue concentration by fathead minnows. Behav Ecol Sociobiol 60:522–527
Ferrari MCO, Messier F, Chivers DP (2007) Degradation of alarm cues under natural conditions: risk assessment by larval amphibians. Chemoecology 17:263–266, DOI 10.10007/s00049-007-0381-0
Field LM, Birkett M, Dufour S, He X, Hooper A, Pickett J, Zhou J-J (2007) Prospects for coupling new molecular approaches to chemical analysis. Contribution at the 23rd ISCE Annual Meeting in Jena, Germany
Freitag J, Ludwig G, Andreini P, Rössler P, Breer H (1998) Olfactory receptors in aquatic and terrestrial vertebrates. J Comp Physiol A 183:635–650
Gerhardt A, de Bisthoven LJ (1995) Behavioural, developmental and morphological responses of Chironomus gr. thummi larvae (Diptera) to aquatic pollution. J Aquat Ecosyst Health 4:205–214
Hanazato T (1999) Anthropogenic chemicals (insecticides) disturb natural organic chemical communication in the plankton community. Environ Pollut 105:137–142
Khafizof K, Anselmi C, Menini A, Carloni P (2007) Ligand specificity of odorant receptors. J Mol Model 13(3):401–409
Klaschka U (2008) Odorants—potent substances at minor concentrations. The ecological role of infochemicals. In: Kümmerer K (ed), Pharmaceuticals in the environment. Sources, fate, effect and risks, 3rd edn. Springer (in press)
Klaschka U, Kolossa-Gehring M (2007) Fragrances in the environment—pleasant odours for nature? Environ Sci Pollut Res 14Special Issue (1):44–52
Krieger J, Breer H (1999) Olfactory reception in invertebrates. Science 286:720–723
Kümmerer K (ed) (2001) Pharmaceuticals in the environment. Sources, fate, effect and risks, 1st edn. Springer
Kümmerer K (ed) (2004) Pharmaceuticals in the environment. Sources, fate, effect and risks, 2nd edn. Springer
Kusch RC, Mirza RS, Chivers DP (2004) Making sense of predator scents: investigating the sophistication of predator assessment abilities of fathead minnows. Behav Ecol Sociobiol 55:551–555
Lahlou M (2004) Essential oils and fragrance compounds—bioactivity and mechanisms of action. Flavour Fragr J 19:159–165
Larsson P, Dodson S (1993) Chemical communication in planktonic animals. Arch Hydrobiol 129:129–155
Larcher W (1995) Physiological plant ecology—ecophysiology and stress physiology of functional groups. Springer, Berlin, Heidelberg, New York
Little EE, Archeski RD, Flerov BA, Kozlovskaya VI (1990) Behavioral indicators of sublethal toxicity in rainbow trout. Arch Environ Contam Toxicol 19:380–389
Little EE, Fairchild JF, DeLonay AJ (1993) Behavioral methods for assessing impacts of contaminants on early life stage fishes. Am Fish Soc Symp 14:67–76
Luckenbach T, Epel D (2005) Nitromusk and polycyclic musk compounds as long-term inhibitors of cellular xenobiotic defense systems mediated by multidrug transporters. Environ Health Perspect 113:17–24
Lüring M, Beckmann W (2002) Extractable substances (anionic surfactants) from membrane filters induce morphological changes in the green alga Scenedesmus obliquus (Chlorophyceae). Environ Toxicol Chem 21:1213–1218
Manzini I, Schild D (2003) Multidrug resistance transporters in the olfactory receptor neurons of Xenopus laevis. J Physiol 546(2):375–385
Michels E, Semsari S, Bin C, de Meester L (2000) Effect of sublethal doses of cadmium on phototactic behaviour of Daphnia magna. Ecotoxicol Environ Saf 47:261–265
Moore A, Lower N (2001) The impact of two pesticides on olfactory-mediated endocrine function in mature male Atlantic salmon (Salmo salar L.) parr. Comp Biochem Physiol B 129(2–3):269–276
Ono M, Terabe H, Hori H, Sasaki M (2003) Components of giant hornet alarm pheromone. Nature 424:637–638
Pichersky E (2004) Plant scents. What we perceive as a fragrant perfume is actually a sophisticated tool used to entice pollinators, discourage microbes and fend off predators. Am Sci 92:514–521
Pohnert G (2004) Chemical defense strategies of marine organisms. Top Curr Chem 239:179–219
Pohnert G, Steinke M, Tollrian R (2007) Chemical cues, defence metabolites and the shaping of pelagic interspecific interactions. Trends Ecol Evol 22:198–204
Pollock MS, Pollock RJ, Chivers DP (2006) The effects of body size, body condition and reproductive state on the responses of fathead minnows to alarm cues. Can J Zool 84:1351–1357
Polya G (2003) Biochemical targets of plant bioactive compounds. A pharmacological reference guide to sites of action and biological effectsTaylor and FrancisLondon and New York
Preston BL, Cecchine G, Snell TW (1999a) Effects of pentachlorophenol on predator avoidance behaviour of the rotifer Brachionus calyciflorus. Aquat Toxicol 44:201–212
Preston BL, Snell TW, Dusenbery D (1999b) The effects of sublethal pentachlorophenol exposure on predation risk in freshwater rotifer species. Aquat Toxicol 47:93–105
Prince EK, Lettieri L, McCurdy KJ, Kubanek J (2006) Fitness consequences for copepods feeding on a red tide dinoflagellate: deciphering the effects of nutritional value, toxicity, and feeding behaviour. Oecologia 147:479–488
Rasmussen LEL, Lee TD, Roelofs EL, Zhang AJ, Daves GD (1996) Insect pheromone in elephants. Nature 379:684
Rossiter KJ (1996) Structure–odor relationships. Chem Rev 96:3201–3240
Shiojiri K, Ozawa R, Takabayashi J (2006) Plant volatiles, rather than light, determine the nocturnal behavior of a caterpillar. PloS Biol 4(6):e164
Smital T, Luckenbach T, Sauerborn R, Hamdoun AM, Vega RL, Epel D (2004) Emerging contaminants—pesticides, PPCPs, microbial degradation products and natural substances as inhibitors of multixenobiotic defense in aquatic organisms. Mutat Res 552:101–117
Stibor H, Lampert W (2000) Components of additive variance in life-history traits of Daphnia hyalina: seasonal differences in the response to predator signals. Oikos 88:129–138
Stoka AM (1999) Phylogeny and evolution of chemical communication—an endocrine approach. J Mol Endocrinol 22:207–225
Tollrian R, Harvell CD (1999) The ecology and evolution of inducible defences. Princeton University Press, Princeton, NJ
Tomba AM, Keller TA, Moore PA (2001) Foraging in complex odour landscapes: chemical orientation strategies during stimulation by conflicting chemical cues. J North Am Benthol Soc 20:211–222
Turner JT, Tester PA (1997) Toxic marine phytoplankton, zooplankton grazers, and pelagic food webs. Limnol Oceanogr 42(5):1203–1214
Waring CP, Moore A (1997) Sublethal effects of a carbamate pesticide on pheromonal mediated endocrine function in mature Atlantic salmon (Salmo salar L.) parr. Fish Physiol Biochem 17:203–211
Weber A (2001) Interactions between predator kairomone and food level complicate the ecological interpretation of daphnia laboratory results. J Plankton Res 23(1):41–46
Wolfe GV (2000) The chemical defence ecology of marine unicellular plankton—constraints, mechanisms and impacts. Biol Bull 198:225–244
Wyatt TD (2003) Pheromones and animal behaviour. Cambridge University Press, Cambridge
Yamashita S, Yamada T, Hara TJ (2006) Gustatory responses to feeding- and non-feeding-stimulant chemicals, with an emphasis on amino acids, in rainbow trout. J Fish Biol 68(3):783–800
Young JM, Trask BJ (2002) The sense of smell: genomics of vertebrate odorant receptors. Hum Mol Genet 11:1153–11607
Zarzo M (2007) The sense of smell: molecular basis of odorant recognition. Biol Rev 82(3):455–479
Zhao H, Firestein S (1999) Vertebrate odorant receptors. Cell Mol Life Sci 56:647–659
Zimmer RK, Butman CA (2000) Chemical signaling processes in the marine environment. Biol Bull 198:168–187
Zou Z, Buck L (2006) Combinatorial effects of odorant mixes in olfactory cortex. Science 311:1477
Electronic supplementary material
The Online Edition of this article is accompanied by a series of slides (texts and diagrams) that you can easily access via this paper. The series titled “Fragrances as infochemicals” responds to the following questions:
-
1.
What are infochemicals?
-
2.
Can anthropogenic substances be infochemicals?
-
3.
Are fragrance ingredients infochemicals?
Each question represents a section. The concluding section provides, as far as is currently possible, information on the following considerations:
-
1.
Will the infochemical effect be a new chapter in ecotoxicology?
-
2.
Which anthropogenic substances are infochemicals?
-
3.
How do the anthropogenic discharges influence natural emissions?
-
4.
Which organisms are affected?
-
5.
Is the ecosystem affected?
The series “Fragrances as infochemicals” makes an ideal co-reading, facilitates the comprehension of the paper and gives new insights in an emerging area of ecotoxicology.
Author information
Authors and Affiliations
Corresponding author
Electronic Supplementary Material
Below is the link to the electronic supplementary material.
ESM
(PDF 1.41 MB)
Rights and permissions
About this article
Cite this article
Klaschka, U. The infochemical effect—a new chapter in ecotoxicology. Environ Sci Pollut Res 15, 452–462 (2008). https://doi.org/10.1007/s11356-008-0019-y
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11356-008-0019-y