
Abstract
We surveyed plant community development at the abandoned Ogushi sulfur mine. We found seven communities dominated by the following respective species: Deschampsia flexuosa, Miscanthus sinensis, shrub willow, Gaultheria miqueliana–Betula ermanii, Sasa senanensis–Betula ermanii, willow–Betula ermanii, and Sasa kurilensis–Abies veitchii. We examined the succession of these communities, in which younger communities of low height and ground cover contained seedlings of the successive communities that were taller and had higher ground cover. To understand the development of these different communities, we surveyed damage from mining pollution and effects of immature soils formed by landslides. The average pH (H2O) was 4.12, and aluminum concentrations were not sufficiently high to damage plant growth, except in areas where sulfur had been mined. The organic carbon and nitrogen content in soil samples were very low because of a delay in soil development caused by a large landslide in 1937. Hence, succession was positively correlated with the soil development stage. The delay in soil development after a large landslide influenced the seven successional steps of the plant communities, but mineral poisons at the abandoned Ogushi sulfur mine had no effect on succession.
Similar content being viewed by others


Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
Vegetation restoration is generally slow at abandoned sulfur mines. For example, vegetation has not yet recovered at the Matsuo sulfur mine site and after strip-mining in Iwate Prefecture, northern Japan, although the mine was abandoned in 1969 (Yuasa et al. 1995a). At the latter site, Iwate Prefecture and the Ministry of Construction covered the mine workings with volcanic ash from surrounding mountains in an attempt to recover natural vegetation, but foreign weeds became established and vegetation diversity declined (Yuasa et al. 1995b). At the Ishizu sulfur mine in Gunma Prefecture, which was abandoned in 1969, the soil is still bare, trees have withered, and grass is the main vegetation over wide areas, suggesting that recovery of vegetation is slow (Shimai 1984). At the Agatsuma sulfur mine in Gunma Prefecture, which operated from 1908 to 1971, deforestation is widespread because of sulfur dioxide from mining operations and tree cutting for fuel. There, original deciduous broadleaf forest is now recovering at the outer edge of the mine, but an open site with less vegetation still remains at the center of the mine site (Oguro et al. 1990).
Delays in vegetation recovery are caused by various factors; for example, smoke, sulfur dioxide, and ferric sulfide from mining operations pollutes the air and water and acidifies the soil. Furthermore, dissolved Al3+ in low-pH soil prevents plant roots from absorbing minerals.
The risk of landslides is also high at abandoned mine workings because the soil is unstable (Yuasa et al. 1995a). The Matsuo mine site, for example, is mostly covered by impermeable tuff that becomes clay that rainwater cannot penetrate, leading to a high risk of erosion and landslides (Yuasa et al. 1995a).
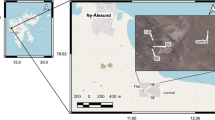
The Ogushi sulfur mine (Fig. 1) was also located on an unstable landform; it suffered a massive landslide in 1937 that covered an area of 84,000 m2 (Institute of Ogushi Mine History 1981a). The debris covered almost the entire mine site. The Ogushi sulfur mine was abandoned in 1971, and there is now a vast area of bare soil where vegetation recovery is slow. We do not know why the vegetation recovery is slow, but there are two possible explanations. One is past smoke, sulfur dioxide, and ferric sulfide from mining operations, and the other is the disturbance caused by the landslide in 1937. It is therefore necessary to understand why vegetation recovery is slow. This involves the examination of changes in the soil and vegetation. The purpose of this study was to understand why vegetation recovery is slow at this site. We examined the successional stages in vegetation recovery with changes in the soil at the abandoned Ogushi sulfur mine.

Location of the Ogushi sulfur mine, Gunma Prefecture, central Japan
Study area
The abandoned Ogushi sulfur mine is located at 36°36′35″N, 138°27′35″E, in Tsumagoi village, Gunma Prefecture (Fig. 1). The mine was operated from 1929 to 1971, and was located on the southeastern slope of Mt. Omeshi at 1,600 m asl, in the southern Nasu volcano belt. Blueprints of the locations of old mining holes are available (Institute of Ogushi Mine History 1981b). The mean annual temperature is 5.0°C, the highest annual temperature is 23.0°C, the lowest annual temperature is −11.6°C, the annual precipitation is 1,524.4 mm, and the maximum snow depth is 0.9 m (from 1971 to 2000; Japan Meteorological Business Support Center 2002). In this region, the warmth index (WI, Kira 1949) is 45.6°C month at 1,600 m asl. Kira’s WI is the sum of mean monthly temperatures above 5°C [WI = ∑(mean monthly temperature − 5°C) only for months warmer than 5°C, in units of °C month]. Kira (1949) noted that cold areas, where WI is lower than 45°C month, are usually covered with subalpine evergreen conifers, whereas warmer areas above 45°C month are covered by montane deciduous broadleaf forests. This suggests that the Ogushi abandoned mine site is in a transition zone from the cool-temperate deciduous broadleaf forest zone to the subalpine evergreen coniferous forest zone. The natural vegetation at this elevation is subalpine evergreen conifer forest with Abies veitchii, or broadleaf deciduous forest with Fagus crenata and Quercus mongolica ssp. crispula. These natural vegetation types are not established at the study site. Outside the mine, there is natural climax vegetation of evergreen conifers such as A. veitchii and A. mariesii, with ground cover of Sasa kurilensis.
The landslide in 1937 washed out organic soil of the mine site, and the center of the mine site was flattened and prepared for mining operations. The flat area would have been trampled heavily until the end of mining in 1971. There are many signs of erosion, especially on slopes (Fig. 2). Most of the abandoned mine site is covered with bare land, and the margin of the mine is covered with some plant communities composed of herbs and/or low-growing young trees.

Landscape of the abandoned Ogushi sulfur mine site. Slopes showed many signs of erosion
Study methods
Species composition and community structure
We surveyed species composition in 84 plots using the Braun-Blanquet approach (Braun-Blanquet 1964) in the summer of 2005. The degree of coverage of each species was recorded by this approach. The sizes of the plots ranged from 25 (5 m × 5 m) to 225 (15 m × 15 m) m2 depending on the community sizes. For trees, the species, height, and diameter at breast height (DBH) of trees taller than 1.3 m were recorded. Basal area (BA, m2/ha) of each plot using the DBHs of trees was calculated.
Raunkiaer’s life form
All species in each plot were classified using Raunkiaer’s life forms (Miyawaki et al. 1994) to understand the successional stages of the communities (Shimano 2007). Early-successional plants have buds on low parts close to the ground, whereas late-successional plants have buds that are higher from the ground, e.g., buds on tree branches. The relative cover of species of each life form in a community is a good indicator of its stage of succession (Shimano 2007). Raunkiaer’s life forms of plants in Japan were obtained from Miyawaki et al. (1994), and Raunkiaer’s life form succession values (RLSV) were obtained from Shimano (2007). This method illustrates both quantitative and qualitative differences in plant communities; e.g., the life form of the Miscanthus community is Hemicryptophyte (H, bud height is 0 m from the ground), whereas that of the Rhododendron community is Nanophanerophyte (N, representative bud height is 1.15 m, see below), even though they have the same vegetation height (1.5 m). To calculate the RLSV, we used the median bud height. The median bud heights of Raunkiaer’s life forms are as follows:
Geophytes | 0 m (buds underground) |
Hemicryptophytes | 0 m (buds on the ground) |
Chamaephytes | 0.15 m (buds from 0 to 0.3 m) |
Nanophanerophytes | 1.15 m (buds from 0.3 to 2 m) |
Mesophanerophytes | 5.0 m (buds from 2 to 8 m) |
Macrophanerophytes | 19.0 m (buds from 8 to 30 m) |
We examined the cover of each species in each plot and calculated the percentage cover; for example, if a plot has 50% Hemicryptophytes, 30% Nanophanerophytes, and 20% Macrophanerophytes, the RLSV is:
Usually, a higher RLSV indicates a later successional stage (Shimano 2007). The plant nomenclature followed that of Satake et al. (1982a, b, c, 1989a, b).
Soil analysis
We analyzed soil pH, extractable aluminum (Ex-Al), soil organic carbon, and nitrogen content. The 84 vegetation plots at the Ogushi sulfur mine were classified into seven communities. We examined the surface soil (0–5 cm) of five to six plots for each community from August to October 2005 (Fig. 3). At the same time, we took soil samples in a sulfur collection area (three samples) and in a bare soil area (three samples). The soil was air-dried, passed through a 2-mm sieve, and analyzed.

Vegetation map and sites for soil collection (filled circle). S in the map shows soil collection sites where sulfur was mined in the past. Bold lines show the boundary of each community. Each number represents each community and site as follows. 0 bare soil, 1 patchy Deschampsia flexuosa community, 2 Miscanthus sinensis community, 3 shrub willow community, 4 Gaultheria miqueliana–Betula ermanii community, 5 Sasa senanensis–Betula ermanii community, 5′ Sasa senanensis–Betula ermanii community with few Betula ermanii, 6 willow–Betula ermanii community. Outside of the area shown in this map were areas covered with a natural Sasa kurilensis–Abies veitchii community (community number 7)
Soil pH
A 10-g sample of air-dried soil was added to 25 ml distilled water. After occasional agitation for 30 min, pH was measured with a glass electrode. After this, 1.86 g KCl was added to the distilled water and soil solution, and the pH was determined in the same way. The addition of KCl allows the measurement of exchangeable acidity associated with exchangeable Al+ and H+ in the clay and humified material in the soil (Nishio et al. 2000).
Ex-Al content
To measure the Ex-Al content, we used KCl extraction (Bertsch and Bloom 1996). A 5-g sample of air-dried soil was added to 25 ml 1 M KCl, agitated for 30 min at 20°C, and centrifuged at 1,100×g for 10 min at 24°C. The supernatant was removed and diluted 100 times to determine the Ex-Al content using furnace atomic absorption spectrophotometry (Perkin-Elmer 5100PC).
Soil organic carbon, nitrogen, sulfur, and moisture content
Soil was crushed in a mortar and the specific gravity was measured. A CHNS analyzer (Thermo Finnigan EA1112) was used to determine the carbon, nitrogen, and sulfur content. Soil moisture was obtained by drying soil in a tumbler at 105°C for 12 h; the soil moisture content was obtained using formula (1).
Results
Plant communities
We classified seven plant communities of different physiognomy with both common and discriminative species: Deschampsia flexuosa, Miscanthus sinensis, shrub willow, Gaultheria miqueliana–Betula ermanii, Sasa senanensis–Betula ermanii, willow–Betula ermanii, and Sasa kurilensis–Abies veitchii (Table 1). Polytrichum commune and Miscanthus sinensis were common except in the Sasa kurilensis–Abies veitchii community. The Deschampsia flexuosa community had only herb or dwarf shrub species, and B. ermanii was common except in the D. flexuosa community. Each community had not only physiognomical differences, but also different species composition.
Each community showed a characteristic distribution in the abandoned mine site (Fig. 3). Bare land was widespread on slopes in the abandoned mine site, where there was no soil, and where small rocks and sand were easily moved by erosion. Most of the bare land showed many signs of erosion (Fig. 2). Deschampsia flexuosa dominated many patches on the flat plain, which showed fewer signs of erosion. The M. sinensis community surrounded the outside of the D. flexuosa patches. Other tree communities of willow and Betula were on the mountainside or on slopes that showed less evidence of erosion. The Sasa kurilensis–Abies veitchii community was outside of the abandoned mine site.
Changes during community development
The number of species, community height, and proportion of ground cover were low in the D. flexuosa community, but high in woody plots such as the S. kurilensis–A. veitchii community (Table 2). BA was significantly correlated with tree height growth in each community (Spearman rank correlation, RS = 0.885, P < 0.001; Table 2, Fig. 4).

Basal area of trees, determined using the diameter measured at breast height (1.3 m). The ends of the bars indicate the maximum and minimum values. The D. flexuosa community (community no. 1) and the M. sinensis community (community no. 2) were omitted because there were no trees. Community codes are provided in Table 1. Mean values are shown in Table 1
Figure 5 shows the Raunkiaer’s life forms of all seven community types in order of tree height and proportion of ground cover. Higher RLSV communities had few Hemicryptophytes and many Nanophanerophytes, Mesophanerophytes, and Macrophanerophytes (RS = 0.575, P < 0.001). In the D. flexuosa community (no. 1), Hemicryptophytes dominated (90%), and most Hemicryptophytes were D. flexuosa. In the M. sinensis community (no. 2), M. sinensis had 50% ground cover. Hemicryptophytes accounted for 70% of plants, including M. sinensis, while Macrophanerophytes accounted for fewer than 10% of plants. In the S. kurilensis–A. veitchii community (no. 7), Hemicryptophytes accounted for fewer than 10% of plants, and Mesophanerophytes and Macrophanerophytes dominated (90%).

Raunkiaer’s life forms and Raunkiaer’s life form succession values (RLSV) with standard deviation. Each community was ordered in a successional series using the life forms of all plants (RS = 0.64, P < 0.01). Community codes are provided in Table 1. G geophyte, H hemicryptophyte, Ch chamaephyte, N nanophanerophyte, M mesophanerophyte, MM macrophanerophyte
The Shannon–Wiener diversity index (H′) for the D. flexuosa community was significantly lower than that for the other communities (Fig. 6a).

a Shannon–Wiener index of diversity H′, b soil pH, c soil organic carbon content, d soil nitrogen content, e carbon/nitrogen ratio, f sulfur content, g soil moisture content, h Ex-Al, Different letters indicate significant differences according to Fisher’s multiple comparisons. In a, H is not shown for sites 01 and 02, because there were no plants. Site no. 01 sulfur collection location, site no. 02 bare soil area
Soil characteristics of the vegetation communities
The D. flexuosa community (no. 1) had no A0 layer and was composed of mineral soil with patchy D. flexuosa (no A0 + B layer). The M. sinensis community (no. 2) had a thin A0 layer with mixed mineral and humic soil in the A layer (thin A0 + thin A + B layers). The shrub willow community (no. 3) had both a thin A0 and a B layer, but no A layer (thin A0 + B layers). The G. miqueliana–B. ermanii community (no. 4) on unstable slopes had an A0 layer, and the B layer was deeper than 5 cm below ground (thin A0 + thin A + B layers). Sasa senanensis blanketed the forest floor in the S. senanensis–B. ermanii community (no. 5), which had a thick A0 layer (thick A0 + thick A + B layers). Here, the B layer was also deeper than 5 cm. The willow–B. ermanii community (no. 6) had a thicker A0 layer (thick A0 + thick A + B layers) than did the S. senanensis–B. ermanii community (no. 5, thick A0 + thick A + B layers). At both sites, B layers were deeper than 5 cm below ground. In the S. kurilensis–A. veitchii community (no. 7), soil in the A layer had an aggregated structure. According to the Geographical Survey Institute (1974) classification, the soil type was a moist podzol.
Soil analysis
The soil pH increased significantly toward neutrality with the development of plant community height (Fig. 6b, RS = 0.669, P < 0.001), although the soil pH was low in the S. kurilensis–A. veitchii and D. flexuosa communities.
The soil organic carbon and nitrogen contents increased significantly with vegetation height (Fig. 6c, d), with higher values in the S. kurilensis–A. veitchii community than in the other communities (RS = 0.950, P < 0.001 for soil organic carbon, and RS = 0.950, P < 0.001 for nitrogen content). The carbon/nitrogen ratio ranged from 10 to 30, and did not differ significantly among the communities (Fig. 6e). The S. kurilensis–A. veitchii community had the highest sulfur content among the communities (Fig. 6f). Bare area (site 01) had higher sulfur content, but was not significantly different to the others.
Soil moisture content was low in the bare soil area and in all communities, except in the S. kurilensis–A. veitchii community (Fig. 6g).
The Ex-Al content differed among the communities, and that in the abandoned sulfur collection area was the highest (Fig. 6h).
Discussion
Successional placement of the seven communities
We expect that community succession at the study site will proceed because ground cover, vegetation height, number of species (Table 2), and BA (Fig. 4) are all increasing as the soil thickens. As soil development proceeds, the D. flexuosa community will be followed by the M. sinensis community, shrub willow community, G. miqueliana–B. ermanii community, S. senanensis–B. ermanii community, willow–B. ermanii community, and finally, the S. kurilensis–A. veitchii community. The RLSVs (Shimano 2007) mostly support this assumption (Fig. 5). Oshima (1988) found that vegetation changes from desert to grassland and to forest, from low to tall plants, and to strongly shade-tolerant plants with general succession. This is accompanied by a gradual increase in the number of species, a small decrease, and then community stratification as succession progresses (Oshima 1988). At the abandoned Ogushi mine site, therefore, the number of species increased with vegetation development, suggesting ordered general succession. Nakamura (1986) found that the D. flexuosa community occurs in the early stages of succession on the bare soil of volcanoes and forms patchy communities on pyroclastic rocks. The patchy communities are then succeeded by the M. sinensis community and the willow–B. ermanii community Empetrum nigrum and Vaccinium uliginosum. The invasion of other plants becomes possible with soil development. If dwarf bamboo does not invade the willow community, it will be succeeded by the willow–B. ermanii community, but if dwarf bamboo invades the willow–B. ermanii community, this community will be succeeded by the S. kurilensis–A. veitchii community, which contains more shade-tolerant species, as the dwarf bamboo regenerates. Under the cover of evergreen dwarf bamboo, willows and B. ermanii, which are both light-demanding deciduous trees, will no longer be able to regenerate (Fig. 7).

Successional series, estimated from the species composition, increase in vegetation height, BA, pH, soil organic carbon and nitrogen contents, change in Raunkiaer’s life forms, and invasion of seedlings and trees
RLSVs also changed with succession. As succession proceeded, the Nano- and Macrophanerophytes increased and the Hemicryptophytes decreased (Fig. 5) as the winter bud height increased. The Chamaephytes increased in the G. miqueliana–B. ermanii community, and the ground surface was covered by G. miqueliana, G. adenothrix, and E. nigrum in this community. However, once soil forms, B. ermanii saplings grow rapidly, leading to a decrease in Chamaephytes.
Seven communities were identified according to species composition, increases in vegetation height, BA, pH, soil organic carbon and nitrogen contents, invasion of seedlings and trees, changes in Raunkiaer’s life forms, and the occurrence of tree seedlings (Fig. 7). The communities will succeed in the order in which they are listed above because small M. sinensis individuals were observed in the D. flexuosa community; B. ermanii seedlings were observed in the M. sinensis community; B. ermanii seedlings were frequent in the willow shrub community; and A. veitchii seedlings were found in the S. kurilensis–A. veitchii community (Fig. 7). No other communities developed at the Ogushi sulfur mine site, apart from A. veitchii forests, because the soil was undeveloped due to a landslide in 1937. The Ex-Al values were not high and the soil organic carbon and nitrogen contents were low, because the soil was undeveloped. The sites that had experienced the most disturbance had younger communities, whereas the less disturbed sites had more developed communities such as the A. veitchii community. Each community at each successional stage will develop as the soil develops.
Soil conditions affecting vegetation development
The soil pHs of the communities were high in herbaceous communities and deciduous broadleaf woods (Fig. 6b). The pHs in S. kurilensis–A. veitchii community were low because humic acid from A. veitchii, an evergreen conifer, dominated in the soil of the S. kurilensis–A. veitchii community; Tsutsumi (1987) pointed out that the soils in conifer forests tend to be acidic because conifer litter is not easily decomposed. Site 01 was a sulfur collection location in past. There pHs were low because sulfur oxidizing bacteria would made SO 2−4 (Stanier et al. 1979). Site 02 was not a sulfur collection location, but it might be affected by 01, because site 02 was surrounded by site 01.
The soil of the communities developed along the successional gradient. The soil organic carbon and nitrogen contents slowly increased with succession to the willow–B. ermanii community (Fig. 6c, d). In all sites, aerobic free-living bacteria that fix nitrogen, such as the Azotobacteriaceae, would fix nitrogen after a landslide. If there were enough thick soil layers, the nitrogen-like nitric acid and ammonia would be retained. In the case of a thin or no soil layer, however, nitrogen would run off with rainwater. Sulfur did not change significantly (Fig. 6f). The sulfur content was higher in soil of the S. kurilensis–A. veitchii community than in the other communities, but did not cause any visible damage to the plants. The source of sulfur at all sites is base rock, and the sulfur would be retained well in soil of the S. kurilensis–A. veitchii community. In other communities, the sulfate ion would be flushed away with rainwater due to no or thin soil layers. The S. kurilensis–A. veitchii community that we surveyed was general climax forest and was located far from the influence of the mine. This community had the highest soil organic carbon and nitrogen contents (19.2 ± 11.0% and 1.09 ± 0.61%, respectively; Fig. 6c, d). At a non-mine site on Mt. Nishikoma, Nagano Prefecture, the soil organic carbon and nitrogen contents of a mixed coniferous forest were 30.7 ± 12.4% and 1.6 ± 0.7%, respectively (Isomura 2004). The values in the S. kurilensis–A. veitchii community that we surveyed were thus within the ranges of reported values.
High Ex-Al values have a negative effect on plant growth. In a burdock crop, which has high sensitivity to Al, root elongation was inhibited at soil Ex-Al >90 mg/kg and was significantly inhibited at >180 mg/kg (Saigusa 1994). In the abandoned sulfur collection location and the M. sinensis community that we examined, Ex-Al was >90 mg/kg, but M. sinensis in this community is an acid-resistant plant (Saigusa 1994; Yuasa et al. 1995b); S. reinii also occurs in highly acidic moors (Matsuda 1979) and was also found in the community we surveyed. At other sites in the Ogushi sulfur mine, the Ex-Al content and soil pH were low, except at the sulfur collection locations. In the sulfur collection locations, the soil had not developed and the soil organic carbon content was low, so the breakdown of organic matter was also low (Fig. 6c, d, f, g). Vegetation restoration is slow at the Ogushi sulfur mine, because the soil remains undeveloped following a large landslide in 1937. If the soil is undeveloped, the supply of nutrients, such as nitrogen and sulfur, for plants will be low. We found no relationship between the level of Al and the delay in vegetation succession.
The soil organic carbon and nitrogen contents were low close to the abandoned mine (Fig. 6c, d) because the soil was undeveloped. After the landslide in 1937, the center of the bare soil area was also the center of the Ogushi mine site; this area was continuously compacted by trampling until 1971, when the mine was abandoned. Under such conditions, vegetation recovery is slow and soil remains undeveloped. More frequent erosion would remove surface soil from the slopes (Fig. 2). Yuasa et al. (1995a) noted that the soil organic carbon content is associated with the standing stock of vegetation, and a high nitrogen content in organic soils will increase plant growth. In the case of the Azuma sulfur mine, located 4 km southeast of the Ogushi sulfur mine, vegetation is recovering because soil has developed due to the lack of landslides, although the vegetation at the former mine was previously devastated because of smoke (Oguro et al. 1990).
The landslide in 1937 widely destroyed vegetation at the Ogushi sulfur mine site. At that time, organic soil with nutrients such as nitrogen and phosphorus ran off, and frequent erosion occurred, which prevented plants from becoming established (Fig. 2). In some stable sites with less erosion, such as the base of a mountain, a plain, or a valley where earth and sand accumulate, succession is proceeding slowly, because the soils of each community were less affected by mining (e.g., low Ex-Al, sulfur content, high pH, Fig. 6c, d, f, h), excluding the D. flexuosa community in the center of the mining site. Woody communities including willows and Betula were found on such stable sites that had soil.
References
Bertsch PM, Bloom PR (1996) Aluminum. In: Sparks DL (ed) Methods of soil analysis. Soil Science Society of America, Madison, pp 517–550
Braun-Blanquet J (1964) Pflanzensoziologie: Grundzüge der Vegetationskunde, 3rd edn. Springer, Wien, p 865
Harada H (1979) Larix lptolepis Gesellshaft. Vegetation of Nagano Prefecture. In: Arbeitsgemeinschaft für die Vegetationskartierung der Präfektur Nagano. Reale Vegetation der Präfektur Nagano: Eine pflanzensoziologishe Studie für den Natur—und Umweltschutz, sowie die Schaffung eines vegetationsreichen Landes der Präfektur Nagano, Präfektur Nagano, pp 77–81 (in Japanese with German summary)
Hirabayashi K (1979) Vegetation of Nagano Prefecture. Vegetation of Nagano Prefecture. In: Arbeitsgemeinschaft für die Vegetationskartierung der Präfektur Nagano. Reale Vegetation der Präfektur Nagano: Eine pflanzensoziologishe Studie für den Natur - und Umweltschutz, sowie die Schaffung eines vegetationsreichen Landes der Präfektur Nagano, Präfektur Nagano, pp 50–51 (in Japanese with German summary)
Institute of Ogushi mine History (1981a) Sight of Ogushi mine (in Japanese). In: Institute of Ogushi mine History. History of Ogushi mine assembly of Ogushi mine, Nagano, pp 65–81 (in Japanese)
Institute of Ogushi mine History (1981b) History of Ogushi mine. In: Institute of Ogushi mine history. History of Ogushi mine assembly of Ogushi mine, Nagano, pp 23–64 (in Japanese)
Isomura I (2004) Relationship between soil properties and chemical forms of Al in forest soils (in Japanese). Graduation thesis of Department of Environmental Sciences, Faculty of Science, Shinshu University, Japan. School year in 2004, pp 1–48 (in Japanese)
Ito S (1979) Menziesia pentandra-Tsuga diversiflia-Gesellschaft. In: Arbeitsgemeinschaft für die Vegetationskartierung der Präfektur Nagano. Reale Vegetation der Präfektur Nagano: Eine pflanzensoziologishe Studie für den Natur - und Umweltschutz, sowie die Schaffung eines vegetationsreichen Landes der Präfektur Nagano, Präfektur Nagano, pp 74–77 (in Japanese with German summary)
Japan Meteorological Business Support Center (2002) Meshed climate data 2000. CD-ROM. Japan Meteorological Business Support Center, Tokyo in Japanese
Kira T (1949) Forest zone in Japan. Forestry explanation series 17, Nippon-Ringyo-Gijutsu- Kyokai, Tokyo (in Japanese)
Kobayashi M (1979) Alnus maximowiczii-Betuletum ermanii. In: Arbeitsgemeinschaft für die Vegetationskartierung der Präfektur Nagano. Reale Vegetation der Präfektur Nagano: Eine pflanzensoziologishe Studie für den Natur - und Umweltschutz, sowie die Schaffung eines vegetationsreichen Landes der Präfektur Nagano, Präfektur Nagano, pp 81–83 (in Japanese with German summary)
Matsuda Y (1979) Empetrum nigrum-Sphagnum papillosum-Gesellschaft. In: Arbeitsgemeinschaft für die Vegetationskartierung der Präfektur Nagano. Reale Vegetation der Präfektur Nagano: Eine pflanzensoziologishe Studie für den Natur - und Umweltschutz, sowie die Schaffung eines vegetationsreichen Landes der Präfektur Nagano, Präfektur Nagano, pp 107–111 (in Japanese with German summary)
Miyawaki A, Okuda S, Fujiwara R (1994) Handbook of Japanese vegetation (new revised edition). Shibundo, Tokyo, p 910 (in Japanese)
Nakamura Y (1986) Alpine wüstenvegetation. In: Miyawaki A (ed) Vegetation of Japan: Kanto. Shibundo, Tokyo, pp 382–385 (in Japanese with German title)
Nishio M, Kozai T, Oku H, Nakasuji F, Oki Y (2000) Soil factor. In: Growth and the environment of crops. Nousan-gyoson-bunkakyoukai, Tokyo (in Japanese)
Oguro T, Takeuchi K, Imagawa T, Takaoka S (1990) Vegetation changes caused by smoke damage in the Agatsuma sulfur mine, Gunma prefecture, central Japan. J Japanese Inst Landsc Archit 53:151–156 (in Japanese with English abstract)
Okuda S (1979) Salix sachalinensis-Gesellschaft. In: Arbeitsgemeinschaft für die Vegetationskartierung der Präfektur Nagano. Reale Vegetation der Präfektur Nagano: Eine pflanzensoziologishe Studie für den Natur - und Umweltschutz, sowie die Schaffung eines vegetationsreichen Landes der Präfektur Nagano, Präfektur Nagano, pp 198–200 (in Japanese with German summary)
Okuda S (1986) Montane Weiden-Vegetation. In: Miyawaki A (ed) Vegetation of Japan: Kanto. Shibundo, Tokyo, pp 307–310 (In Japanese with German title)
Okuhara H (1990) Wildflower of Shinshu. Shinano Mainichi Shimbun, Nagano, p 19
Oshima Y (1988) Succession of biological community. In: Oshima Y, Okuma M, Sato M, Takahashi K, Mizuno T, Yamada A (eds) Biology. Asakura, Tokyo, pp 253–258
Saigusa M (1994) Acid soils and plants. In: Japanese society of soil science and plant nutrition (ed.) Japanese J soil Sci plant nutr. Hakuyu, Tokyo, pp 43–57 (in Japanese)
Sasaki Y (1979) Boehmerio-Artemisitum montanae. In: Arbeitsgemeinschaft für die Vegetationskartierung der Präfektur Nagano. Reale Vegetation der Präfektur Nagano: Eine pflanzensoziologishe Studie für den Natur - und Umweltschutz, sowie die Schaffung eines vegetationsreichen Landes der Präfektur Nagano, Präfektur Nagano, pp 204–205 (in Japanese with German summary)
Satake Y, Ohwi J, Kitamura S, Watari S, Tominari T (eds) (1982a) Wild flowers of Japan: herbaceous plants 1. Heibonsha, Tokyo, p 305 in Japanese
Satake Y, Ohwi J, Kitamura S, Watari S, Tominari T (eds) (1982b) Wild flowers of Japan: herbaceous plants 2. Heibonsha, Tokyo, p 318 (in Japanese)
Satake Y, Ohwi J, Kitamura S, Watari S, Tominari T (eds) (1982c) Wild flowers of Japan: herbaceous plants 3. Heibonsha, Tokyo, p 259 (in Japanese)
Satake Y, Hara H, Watari S, Tominari T (eds) (1989a) Wild flowers of Japan: woody plants 1. Heibonsha, Tokyo, p 321 (in Japanese)
Satake Y, Ohwi J, Kitamura S, Watari S, Tominari T (eds) (1989b) Wild flowers of Japan: woody plants 2. Heibonsha, Tokyo, p 305 (in Japanese)
Shimai S (1984) Vegetation of destruction area in sulfur mines. Bulletin of Daito Bunka University 22:197–204 (in Japanese with English abstract)
Shimano K (2007) Key points of regeneration-dynamics study in forests and fields. In: Minami Y, Okitsu S (eds) Basic master: ecology. Ohmsha, Tokyo, pp 30–56 (in Japanese)
Shimizu T (1979) Betuletum ermanii-Gesellschaft. In: Arbeitsgemeinschaft für die Vegetationskartierung der Präfektur Nagano. Reale Vegetation der Präfektur Nagano: Eine pflanzensoziologishe Studie für den Natur - und Umweltschutz, sowie die Schaffung eines vegetationsreichen Landes der Präfektur Nagano, Präfektur Nagano, pp 273–274 (in Japanese with German summary)
Shimizu T (2002) Alpine plants. Yama-Kei, Tokyo, p 495 (in Japanese)
Stanier RY, Adelberg EA, Ingraham JL, Wheelis ML (1979) Introduction to the microbial world. Prentice-Hall, Englewood Cliffs, p 468
Suzuki K (1979) Gentiana triflora var. japonica - Miscanthus sinensis Gesellschaft. In: Arbeitsgemeinschaft für die Vegetationskartierung der Präfektur Nagano. Reale Vegetation der Präfektur Nagano: Eine pflanzensoziologishe Studie für den Natur - und Umweltschutz, sowie die Schaffung eines vegetationsreichen Landes der Präfektur Nagano, Präfektur Nagano, p 300 (in Japanese with German summary)
Tsutsumi T (1987) Physical cycles and soils in forests. In: Physical cycles in forests. Tokyo University Publishers, Tokyo, pp 1–11 (in Japanese)
Yuasa Y, Murai H, Hamaura K, Inoue K (1995a) Soil properties of revegetated open-cut mining lands in the past Matsuo sulfur mine, Iwate Prefecture. J Sci Soil Manure Jpn 66:520–526 (in Japanese with English abstract)
Yuasa Y, Sawada K, Murai H, Inoue K (1995b) Vegetational changes in revegetated open-cut mining lands of the former Matsuo sulfur mine, Iwate Prefecture. J Sci Soil Manure Jpn 66:646–654 (in Japanese with English abstract)
Acknowledgments
We thank Professor T. Kunito and Professor F. Kumon, Faculty of Science, Shinshu University, for their helpful suggestions. We acknowledge the anonymous referees and the corresponding editor.
Author information
Authors and Affiliations
Corresponding author
Appendix
Appendix
Characteristics of the communities
Deschampsia flexuosa community
Deschampsia flexuosa formed patchy communities on bare mineral soil. Typical species of the community were Reynoutria japonica, Calamagrostis arundinacea, and E. nigrum (Table 1). D. flexuosa is the characteristic plant of the phytosociological association Deschampsio–Stelarietum nipponicas, which dominates poor communities in the most unstable habitats of Mt. Norikura and Mt. Ontake (Hirabayashi 1979) and is a pioneer community on dry collapsed slopes. R. japonica was also observed by Hirabayashi (1979) on volcanic dust on Mt. Asama, where volcanic activity continues. The D. flexuosa community is the pioneer vegetation at the abandoned Ogushi sulfur mine.
M. sinensis community
The M. sinensis community is dominated by M. sinensis and contains the typical herbaceous species Seseli libanotis and Anaphalis margaritacea (Table 1). The M. sinensis community at the abandoned Ogushi sulfur mine is similar to the Gentina truflora var. japonica–M. sinensis community reported by Suzuki (1979) as pioneer vegetation in secondary grassland following burning or mowing. Because there were no human impacts at Ogushi after the mine was abandoned and the vegetation height was taller than the D. flexuosa community (Table 2), shrubs invaded (Fig. 4). Harada (1979) found A. margaritacea in an E. nigrum var. japonicum–Larix leptolepis community on sunny bare soils. At Younotaira on Mt. Asama (2,140 m) and at Takamine spa (2,040 m), A. margaritacea grows with pioneer plants in sunny habitats where the soil is unstable (Okuhara 1990), e.g., bare soil in landslide areas or sites of volcanic eruption. Hypericum kamtschaticum and Seseli libanotis in this community grow in open or dry grassland and gravel areas (Shimizu 2002). Although the development of soil was poor in this community, it was more developed compared with the soil of the D. flexuosa community (Fig. 6), because the M. sinensis site would have developed over a longer time than D. flexuosa sites. This community will be succeeded by B. ermanii forest because it contained B. ermanii seedlings.
Shrub willow community
Salicaceae dominated the shrub willow community; this community usually occurs within the Saxifraga cherlerioides var. rebunshirensis–Stellaria ruscifolia community, which is mainly distributed on sunny rocks or coarse fragmented areas without soil (Okuda 1979). Artemisia montana is a tall perennial herb in the Boehmerio–Artemisitum montanae association described by Sasaki (1979) and develops in mountainous areas in higher regions of the Fagetea–crenatae association, which grows as coarse fragments in poorly developed forests. This community is part of the primary succession at abandoned mine sites because it has many species in common with the M. sinensis community. The soil is better developed and Raunkiaer’s life form indices are higher than in the M. sinensis community.
Gaultheria miqueliana–B. ermanii community
Betula ermanii is approximately 1.7 m tall and dominated the G. miqueliana–B. ermanii community. The dwarf shrubs G. miqueliana and Vaccinium vitis-idaea covered the ground (Table 2). Ito (1979) noted that E. nigrum and G. miqueliana in this community occur on habitats with exposed parent rock or shallow soil with exposed tree roots. Shimizu (1979) observed the B. ermanii community in sites with thick soil and shallow moving soil on steep slopes. This community occurs on unstable soils. B. ermanii is the pioneer tree, and the succession is more advanced than in the above described herbaceous communities.
S. senanensis–B. ermanii community
Sasa senanensis dominated under the open canopy of B. ermanii. S. senanensis grows on the floor of natural forests that contain Fagus crenata and Abies mariesii (Ito 1979). Kobayashi (1979) found that S. senanensis usually grows under strong artificial influences; he reported that Alnus maximowiczii–B. ermanii forests that had S. senanensis on the forest floor would succeed to coniferous forest because of the presence of seedlings of coniferous trees, stable soil, and the increasing vegetation height. Dwarf bamboo has underground rhizomes, which indicates that the soil in this community is more developed compared with that of the shrub willow and G. miqueliana–B. ermanii communities.
Willow–B. ermanii community
Betula ermanii and the willows Salix sachalinensis, S. integra, and S. reinii dominated the willow–B. ermanii community. S. sachalinensis usually occurs from montane to lower montane regions and dominates riversides (Okuda 1979). Okuda (1979) pointed out that the forest floor of a S. sachalinensis community is unstable, but that S. sachalinensis can tolerate unstable sites. Okuda (1986) found S. sachalinensis and S. integra in the Coriario–Elaeagnetum umbellatae with pioneer summer-green shrubs such as Weigelia hortensis and Elaeagnus umbellata. Seedlings of A. veitchii in the willow–B. ermanii community will grow and create climax vegetation.
S. kurilensis–A. veitchii community
Conifers such as A. veitchii and Tsuga diversifolia dominated the S. kurilensis–A. veitchii community, and S. kurilensis (1.5–2.0 m high) dominated the forest floor. Abies veitchii, the most dominant species, is part of the Abietetum veitchii–mariesii and is a climax species at 1,800–2,000 m (Ito 1979). Salix bakko and Menziesia multiflora in this community also occur in the Abietetum veitchii–mariesii (Ito 1979). In addition, there were A. veitchii seedlings on the forest floor. Therefore, the S. kurilensis–A. veitchii community is the climax vegetation in this area.
Rights and permissions
About this article
Cite this article
Takeuchi, K., Shimano, K. Vegetation succession at the abandoned Ogushi sulfur mine, central Japan. Landscape Ecol Eng 5, 33–44 (2009). https://doi.org/10.1007/s11355-008-0057-0
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11355-008-0057-0