
Abstract
Nicotinamide adenine dinucleotide (NAD) belongs to the family of naturally occurring adenine dinucleotides, best known for their various intracellular roles. However, there is evidence that they can also be released from cells to act as novel extracellular signalling molecules. Relatively little is known about the extracellular actions of NAD, especially in the cardiovascular system. The present study investigated the actions of NAD in the rat thoracic aorta, porcine coronary artery and porcine mesenteric arteries, mounted in organ baths for isometric tension recording. In the rat thoracic aorta and porcine coronary artery, NAD caused endothelium-independent concentration-dependent vasorelaxations which were unaffected by palmitoylCoA, a P2Y1 receptor antagonist, but which were blocked by CGS15943, a non-selective adenosine receptor antagonist. In the porcine coronary artery, NAD-evoked relaxations were abolished by SCH58261, a selective A2A receptor antagonist. In the rat thoracic aorta, NAD-evoked relaxations were attenuated by A2A receptor antagonism with SCH58261 but were unaffected by an A2B receptor antagonist, MRS1754. In contrast, in the porcine mesenteric artery, NAD-evoked endothelium-independent contractions, which were unaffected by a P2 receptor antagonist, suramin, or by NF449, a P2X1 receptor antagonist, but were attenuated following P2X receptor desensitisation with αβ-meATP. In conclusion, the present results show that NAD can alter vascular tone through actions at purine receptors in three different arteries from two species; its molecular targets differ according to the type of blood vessel.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
P1 and P2 receptors for purine and pyrimidine nucleosides and nucleotides are widely distributed in the cardiovascular system and are involved in diverse functions, including regulation of vascular contractility, growth and inflammation [1–4]. P1 receptors mediate the actions of adenosine, and P2X and P2Y receptors mediate the actions of ATP, ADP, UTP, UDP and UDP sugars. There are four adenosine receptors (A1, A2A, A2B and A3), seven P2X receptors (P2X1-7) and eight P2Y receptors (P2Y1, 2, 4, 6, 11, 12, 13, 14). Nicotinamide adenine dinucleotide (NAD) and NAD phosphate (NADP), and their reduced forms NADH and NADPH, belong to the family of naturally occurring adenine dinucleotides. These molecules are best known for their various intracellular roles, including actions as coenzymes, involvement in post-translational modification of proteins and as substrates for NADPH oxidases [5–7]. However, there is evidence that they can also be released from cells to act as novel extracellular signalling molecules. NAD (Fig. 1) is released in a variety of smooth muscle tissues during stimulation of nerves, including those in canine mesenteric artery and urinary bladder of human, mouse and dog [8–11]. Moreover, transporters for NAD, including connexin 43, have been identified that mediate both intercellular and intracellular transport of NAD through membranes [12–14], although this is controversial [15].

Chemical structure of nicotinamide adenine dinucleotide
Relatively little is known about the extracellular actions of NAD, especially in the cardiovascular system, but evidence obtained primarily in tissues outside the cardiovascular system indicates that NAD can act at cell surface purine receptors. Many of the actions of NAD can be explained through activation of adenosine receptors [16–18]. NAD can be cleaved by ectoenzymes; nucleotide phosphodiesterase/pyrophosphatase I (E-NPP, CD203 family) allows the production of AMP, which can be hydrolyzed to adenosine by ecto-5′-nucleotidase (CD73). The ectoenzyme CD38 degrades NAD to yield ADP ribose (ADPR), cyclic ADP ribose (cADPR) and nicotinic acid adenine dinucleotide phosphate (NAADP), and its cell surface location suggests an important role in recycling of extracellular nucleotides [12, 19, 20]. NAD has also been identified as an agonist at P2Y1 receptors in HEK cells and mouse colonic muscle [10], P2Y11 receptors in human granulocytes [21] and P2X receptors in human monocytes (P2X1 and possibly also P2X4 and P2X7) [22]. Multiple purine receptors are co-expressed in blood vessels; vasocontraction is typically mediated by P2X1, P2Y2, P2Y4 and P2Y6 receptors expressed on the smooth muscle, while vasorelaxation is mediated by endothelial P2Y1, P2Y2 and P2Y6 receptors and by A2A and A2B receptors expressed on the endothelium and smooth muscle [3]. Hence the vascular actions of NAD cannot be predicted but must be determined empirically.
In this study, responses to NAD in rat thoracic aorta, porcine mesenteric artery and porcine coronary artery were investigated using selective P1 and P2 receptor antagonists. We have recently shown that palmitoyl CoA (PaCoA) is an antagonist at P2Y1 receptors in the rat thoracic aorta and porcine mesenteric artery [23]. Hence, PaCoA was used to characterise the responses to NAD in different blood vessels, specifically to investigate the possible involvement of P2Y1 receptors. It has previously been shown that vasorelaxant P2Y1, P2Y2 and A2 receptors are expressed on the endothelium of the rat thoracic aorta [24, 25]. Porcine coronary arteries are reported to express vasorelaxant P2Y1, A1, A2A and A2B receptors [26–31]. Little is known about purine receptor expression in the porcine mesenteric artery, however, ADP-mediated relaxations through P2Y1 receptors have been reported [23]. The present results show that NAD can alter vascular tone through actions at purine receptors in three different arteries from two species; its actions differ according to the type of blood vessel, since NAD-evoked P2X-like receptor mediated contraction in the porcine mesenteric artery, but A2A receptor mediated relaxation in the porcine coronary artery and rat thoracic aorta.
Materials and methods
Porcine mesenteries and hearts, obtained from a local abattoir (Woods abattoir, Clipstone, Mansfield, Nottinghamshire) and male Wistar rats (200–250 g), obtained from Charles River (England, UK), were used in this study. Porcine first-order mesenteric arteries and coronary arteries were dissected out and stored overnight in Krebs-Henseleit solution (at 4 °C) for use the following day. Rat thoracic aorta were dissected out, and rings (3–4 mm in length) from these and the porcine mesenteric and coronary arteries were mounted for isometric recording in Krebs-Henseleit solution, oxygenated (95 % O2, 5 % CO2) and incubated at 37 °C as described previously [23]. Rat thoracic aortae were initially tensioned to 1 g and allowed to equilibrate for 30 min after which a further 1 g of tension was applied and the rings left to equilibrate for a further 30 min. Porcine coronary and mesenteric arteries were tensioned to 10 g and allowed to equilibrate for 1 h. After equilibration, arteries were contracted twice to KCl (60 mM), with washout after each addition, to act as an internal standard. Subsequently, arteries were preconstricted using methoxamine for the rat thoracic aorta and U46619, a thromboxane A2 mimetic, for porcine mesenteric and coronary arteries. Once a stable precontracted tone had been achieved, stepwise cumulative addition of NAD (0.1 μM–1 mM) to the preparations was carried out, in the absence and presence of selective antagonists. The antagonists/blockers used were: PaCoA (10 μM; a putative endogenous antagonist of P2Y1 receptors), CGS15943 (10 μM; an adenosine receptor antagonist), SCH58261 (100 nM; a selective A2A receptor antagonist), MRS1754 (1 μM; a selective A2B receptor antagonist), suramin (100 μM; a P2 receptor antagonist), αβ-meATP (10 μM; a P2X receptor agonist and desensitising agent) and NF449 (10 μM; a P2X1 receptor antagonist). These compounds were added 10 min before methoxamine or U46619 addition and so were in contact with the tissue for at least 30 min before addition of NAD. In some preparations, the endothelium was removed by gentle rubbing with the success of the treatment evaluated by testing responses to acetylcholine in the rat aorta and substance P in the porcine arteries as described previously [23]. U46619 and substance P were used in the pig arteries, because they respond poorly to methoxamine and acetylcholine (used in the rat aorta).
Materials
Krebs-Henseleit buffer was composed of the following (mM); NaCl 118, KCl 4.8, CaCl2·H2O 1.3, NaHCO3 25.0, KH2PO4 1.2, MgSO4·7H2O 1.2 and glucose 11.1. The supplier for chemicals in this work was Sigma Chemical Company, except for SCH58261 which was a gift from Schering Plough, Milan, Italy, and NF449 which was from Tocris Bioscience. All drugs were dissolved in water except for CGS15943 and SCH58261, which were dissolved in dimethyl sulfoxide.
Statistical analysis
Responses are expressed as a percentage of the methoxamine- or U46619-induced tone. Data are expressed as mean ± standard error of the mean. Two-way analysis of variance (ANOVA) with a Bonferroni post-hoc test was used for statistical comparisons. P < 0.05 was taken as statistically significant.
Results
KCl (60 mM) produced sustained contractions in the segments of rat thoracic aorta, porcine mesenteric artery and porcine coronary artery with mean responses of 0.91 ± 0.08 g (n = 35), 9.21 ± 0.43 g (n = 20) and 9.96 ± 0.20 g (n = 32), respectively.
Effect of NAD in precontracted rat thoracic aorta, porcine mesenteric artery and porcine coronary artery
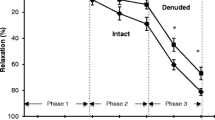
In the rat thoracic aorta, NAD evoked concentration-dependent relaxations (Figs. 2a and 4). The response to NAD did not reach a maximum response at concentrations up to 1 mM but almost fully reversed the methoxamine-induced precontraction allowing the calculation of an approximate pEC50 value of 4.24 ± 0.19 (n = 9). Endothelium removal had no significant effect on the NAD-induced relaxations (two-way ANOVA; P > 0.05) (Fig. 2a).

The effect of nicotinamide adenine dinucleotide (NAD) in segments from: a rat thoracic aorta (RTA), b porcine mesenteric artery (PMA) and c porcine coronary artery (PCA). Arteries were precontracted with methoxamine a and U46619 (b, c). Responses were evaluated in endothelium intact vessels (Control) and in those in which the endothelium had been removed (Denuded) in each of the RTA, PMA and PCA (n = 7–10, n = 6–11, n = 5–6, respectively). Results are mean ± SEM
In porcine mesenteric artery segments precontracted with U46619, NAD evoked concentration-dependent contractions with a maximal contraction at 300 μM (Figs. 2b and 6). The highest concentration of NAD used (1 mM) caused a reduced contraction of the porcine mesenteric artery. Non-linear analysis allowed computation of an R max value of 46 ± 8 %, pEC50 value of 4.54 ± 0.22 and a Hill slope of 1.05 ± 0.12 (n = 12). Endothelium removal had no significant effect on the NAD-induced responses (Fig. 2b).
In porcine coronary artery segments precontracted with U46619, NAD evoked concentration-dependent relaxations (Fig. 2c). The response to NAD did not reach a maximum response, so R max, pEC50 or Hill slope could not be calculated. Endothelium removal produced no significant effect on the NAD-induced relaxations (Fig. 2c).
In porcine mesenteric artery and porcine coronary artery segments, endothelium removal was assessed using substance P (10 nM), while acetylcholine (100 nM) was used to assess endothelium removal in the rat thoracic aorta. Any relaxation in response to substance P/acetylcholine of less than 10 % of the U46619/methoxamine contraction was considered an indication of successful removal of the endothelium. In endothelium-denuded porcine mesenteric artery and porcine coronary artery, there was an absence of the transient relaxation in response to substance P (6 ± 1 % (n = 6) and 2 ± 1 % (n = 5), respectively) that was seen in control porcine mesenteric and porcine coronary artery segments (60 ± 1 % (n = 11) and 70 ± 1 % (n = 6), respectively). In rat thoracic aorta, there was only a very slight relaxation (2 ± 0.4 %, n = 7) in response to acetylcholine in endothelium-denuded vessels compared with the profound relaxation evoked in controls (65 ± 0.4 %, n = 10).
Effect of P1 and P2 receptor antagonists on responses to NAD in the rat thoracic aorta
Since NAD is able to act as an agonist at the P2Y1 receptor, PaCoA was used to investigate whether NAD activates P2Y1 receptors in the rat thoracic aorta. PaCoA had no significant effect on the vasorelaxant response to NAD (P > 0.05, two-way ANOVA) (Fig. 3a).

Relaxatory responses in the rat thoracic aorta to nicotinamide adenine dinucleotide (NAD) in the presence of a PaCoA, b CGS15943, c SCH58261 and d MRS1754. Dimethyl sulfoxide (DMSO) is the vehicle control for CGS15943 and SCH58261. Vessels were precontracted with methoxamine. Results are shown as mean ± SEM (n = 5–6, n = 4–5, n = 12–15 and n = 4–8, respectively). *P < 0.05; **P < 0.01; ***P < 0.001
Since NAD is able to act as an adenosine receptor agonist, CGS15943 (1 μM; a non-selective adenosine receptor antagonist) was also investigated, which significantly inhibited the NAD-evoked relaxations (Fig. 3b). SCH58261 (100 nM; an A2A receptor-selective antagonist) was used, which also significantly decreased the NAD-evoked relaxations with a calculated pK B value of 7.25 ± 0.24 (Fig. 3c). A representative trace showing the inhibitory effect of SCH58261 on responses to NAD is shown in Fig. 4. The possible involvement of A2B receptors was also investigated using MRS1754 (1 μM; a selective A2B receptor antagonist), which had no significant effect on NAD-evoked relaxations (Fig. 3d).

A representative trace for the relaxatory responses to cumulative addition of increasing concentrations of nicotinamide adenine dinucleotide (NAD) in the rat thoracic aorta in the absence (Control, upper trace) and the presence of SCH58261 (SCH58261, lower trace). Vessels were precontracted with methoxamine
These experiments in rat thoracic aorta were carried out in methoxamine-precontracted tissues. In the absence of antagonists, methoxamine caused a sustained contraction to 68 ± 6 % (n = 13) of the KCl response. The bath concentration of methoxamine required to produce this level of contraction was 1.4 ± 0.2 μM (n = 13). In the presence of PaCoA, SCH58261 and CGS15943, the level of tone was 70 ± 6 % (n = 6), 74 ± 5 % (n = 15) and 59 ± 4 % (n = 5) of the KCl contraction, respectively. There was no significant difference between the percentage contractions obtained to methoxamine in the presence of the antagonists and the control (one-way ANOVA, P > 0.05). In the presence of PaCoA, SCH58261 and CGS15943, the mean bath concentration of methoxamine required to precontract the arteries was also unchanged when compared with the control (one-way ANOVA, P > 0.05).
Effect of P1 and P2 receptor antagonists on responses to NAD in porcine mesenteric artery
To test for any involvement of P2 receptors in the NAD-mediated contractile response of the porcine mesenteric artery, suramin (100 μM; a P2 receptor antagonist) was employed; it had no significant effect except at the highest NAD concentration where suramin prevented the reduction in NAD-evoked contraction (Fig. 5a). By contrast, αβ-meATP (10 μM; a P2X receptor desensitising agonist) caused a significant inhibition of the NAD-evoked responses (Figs. 5b and 6). αβ-meATP markedly inhibited or abolished the NAD responses of the mesenteric arterial preparations (thus an EC50 value could not be calculated). A mean contraction of 45 ± 1 and 17 ± 7 % (n = 7 and n = 8, respectively) at 300 μM NAD was found in the absence and presence of αβ-meATP, respectively. NF449 (10 μM; P2X1 receptor antagonist) had no significant effect on the contractile response to NAD; mean contraction at 300 μM NAD was 36 ± 5 and 37 ± 7 % (n = 9) in the absence and presence of NF449, respectively.

The contractile response of porcine mesenteric artery to nicotinamide adenine dinucleotide (NAD) in the absence and presence of a suramin and b αβ-meATP. The arteries had been precontracted with U46619. Results are shown as mean ± SEM (n = 4 and n = 7–8, respectively). **P < 0.01; ***P < 0.001

A representative trace for the contractile response to nicotinamide adenine dinucleotide (NAD) of porcine mesenteric artery in the absence (Control, upper trace) and presence of αβ-meATP (αβ-meATP, lower trace). The arteries had been precontracted with U46619
Since multiple contractile and relaxant purine receptors are typically expressed on blood vessels, we hypothesised that the contractile actions of NAD could be augmented following antagonism of NAD-mediated vasorelaxation. Since NAD can act as an agonist at the P2Y1 receptor, the effect of PaCoA was investigated. PaCoA at 10 μM had no significant effect on the response to NAD (two-way ANOVA, P > 0.05) (Fig. 7a). NAD can also act at P1 receptors, hence the effects of CGS15943 (10 μM; a non-selective adenosine receptor antagonist) and SCH58261 (100 nM; a selective A2A receptor antagonist) were investigated; neither had any significant effect on the response to NAD (Fig. 7b, c). In separate experiments, adenosine produced concentration-dependent relaxation of the porcine mesenteric artery with an R max of 87 ± 4 % and a pEC50 of 3.87 ± 0.07 (n = 6).

The porcine mesenteric artery contractile response to nicotinamide adenine dinucleotide (NAD) in the presence of a PaCoA, b CGS15943 and c SCH58261 (n = 7, n = 8 and n = 4, respectively). The arteries had been precontracted with U46619. Results are shown as mean ± SEM
The above experiments in porcine mesenteric artery were carried out in vessels precontracted with U46619. In the absence of antagonists, U46619 caused a sustained contraction to 75 ± 4 % (n = 7) of the KCl response. The concentration of U46619 required to produce this level of contraction was 131 ± 18 nM (n = 7). In the presence of PaCoA, SCH58261, suramin and αβ-meATP, U46619 elicited 65 ± 9 % (n = 7), 63 ± 5 % (n = 4), 50 ± 10 % (n = 4) and 58 ± 3 % (n = 8) of the KCl contraction, respectively, which was not significantly different from the control (one-way ANOVA, P > 0.05). The concentration of U46619 required to produce these contractions was unchanged (one way ANOVA, P > 0.05). In the presence of CGS15943, a higher concentration of U46619 (763 ± 259 nM, n = 6) was required to precontract the preparations compared with that used in the absence of CGS15943 (531 ± 216 nM, n = 7). There was no significant difference in the level of tone elicited in the absence and presence of CGS15943 at 55 ± 6 % (n = 7) and 49 ± 10 % (n = 6) of the KCl contraction, respectively. There was no significant difference in the level of tone elicited in the absence and presence of NF449, at 61 ± 5 and 62 ± 6 % (n = 9, n = 9), respectively.
Effect of P1 and P2 receptor antagonists on responses to NAD in porcine coronary artery
Since NAD can act as an agonist at the P2Y1 receptor, PaCoA (10 μM) was used to investigate whether NAD activates P2Y1 receptors in the porcine coronary artery. PaCoA had no significant effect on the vasorelaxant response to NAD (two-way ANOVA, P > 0.05) (Fig. 8a). To characterise the NAD-evoked relaxations, we used CGS15943 (10 μM; adenosine receptor antagonist) which, with the exception of the response to the highest concentration of NAD (1 mM), abolished the NAD-evoked relaxations (Fig. 8b). In the presence of CGS15943, contractions were observed at 10–100 μM of NAD; these were unaffected by the P2X1 receptor antagonist NF449 (10 μM; data not shown). SCH58261 (100 nM; selective A2A receptor antagonist) abolished the NAD-evoked relaxations (Figs. 8c and 9).

The porcine coronary artery relaxation to nicotinamide adenine dinucleotide (NAD) in the presence of a PaCoA, b CGS15943 and c SCH58261. Preparations were precontracted with U46619. Results are shown as mean ± SEM (n = 5, n = 6 and n = 6 respectively). *P < 0.05; **P < 0.01; ***P < 0.001

A representative trace for the porcine coronary artery relaxation to nicotinamide adenine dinucleotide (NAD) in the absence (Control, upper trace) and presence of SCH58261 (SCH58261, lower trace). Vessels were precontracted with U46619
The above experiments in porcine coronary artery were carried out in U46619-precontracted tissues. In the absence of antagonists, U46619 caused a sustained contraction to 60 ± 6 % (n = 9) of the KCl response. The concentration of U46619 required to produce this level of contraction was 145 ± 62 nM (n = 9). In the presence of PaCoA and SCH58261, U46619 elicited 65 ± 5 % (n = 5) and 76 ± 2 % (n = 6) of the KCl contraction, respectively, which was not significantly different from the control (one-way ANOVA, P > 0.05). The mean bath concentration of U46619 required was unchanged (one-way, ANOVA P > 0.05).
In the presence of CGS15943, a higher concentration of U46619 was required to precontract the PCA preparations; for these experiments, 175 ± 79 nM (n = 7) of U46619 was needed to achieve 57 ± 7 % (n = 7) of the KCl contraction in controls, while 773 ± 204 nM (n = 6) of U46619 was needed to elicit 24 ± 4 % (n = 6) of the KCl contraction in the presence of CGS15943. Both the level of contraction to U46619 and the concentration of U46619 required to achieve that level were significantly different from the control (one-way ANOVA, P < 0.05).
Discussion
The effects of NAD on the rat thoracic aorta, porcine mesenteric artery and porcine coronary artery were examined. NAD evoked vasorelaxations through actions at smooth muscle A2A adenosine receptors in the rat thoracic aorta and porcine coronary artery. In the porcine mesenteric artery, however, NAD caused vasoconstriction, which was mediated through P2X-like receptors. NAD, therefore, evokes different effects (relaxation or contraction) in different blood vessels acting via different receptors.
In the rat thoracic aorta and porcine coronary artery, NAD caused endothelium-independent vasorelaxations. The effect of PaCoA, a P2Y1 receptor antagonist [23, 32, 33], on the NAD-evoked relaxations was investigated, since NAD can act as an agonist at P2Y1 receptors [10]. PaCoA had no significant effect on the NAD-evoked relaxations, which, together with the fact that these relaxations were endothelium independent (P2Y1 receptors are expressed on the endothelium in the rat aorta, see ‘Introduction’), indicates that P2Y1 receptors do not mediate these vasorelaxations. NAD has previously been shown to act as an agonist at adenosine receptors, without specification of which subtype was involved [16–18]. Hence, P1 receptor antagonists were initially used to characterise the NAD-mediated relaxations, followed by the use of more selective adenosine receptor antagonists. CGS15943, a non-selective adenosine receptor antagonist, significantly reduced the NAD-evoked relaxations, indicating the involvement of adenosine receptors. SCH58261, a selective A2A receptor antagonist, also significantly reduced the NAD-evoked relaxations, which indicates that NAD-evoked relaxations in the rat thoracic aorta and porcine coronary artery are mediated through A2A adenosine receptors. In contrast to the porcine coronary artery, relaxations to NAD in the rat thoracic aorta were not abolished in the presence of SCH58261, which may indicate the involvement of other adenosine receptors in addition to A2A receptors. An A2B adenosine receptor antagonist, MRS1754, had no significant effect on the NAD-evoked relaxations in the rat thoracic aorta, which appears to exclude an involvement of A2B adenosine receptors. In the rat thoracic aorta, NAD had an approximate pEC50 value of 4.24. This is similar to the pD2 value reported by Burnstock and Hoyle [16] for NAD-evoked relaxations in the guinea-pig taenia coli at P1 receptors (4.18) but is somewhat different from the pEC50 value identified by Mutafova-Yambolieva et al. [10] for NAD-evoked calcium responses in HEK cells at P2Y1 receptors (6.1). The potency differences presumably reflect simple differences in the receptors being activated. In rat thoracic and porcine coronary arteries, NAD appears to act at adenosine receptors, specifically as an A2A receptor-selective agonist.
In the rat thoracic aorta and porcine coronary artery, NAD-evoked relaxations were mediated mainly via actions at smooth muscle A2A receptors. This may be by NAD activating A2A receptors directly, by causing a release of adenosine or by being broken down into adenosine. NAD is suggested to be broken down to adenosine to exert its effects in the guinea-pig taenia coli [16]. However, NAD was reported to evoke the release of adenosine in rat vas deferens, guinea-pig taenia caecia and bladder [34]. We have previously shown in the porcine coronary artery that ADP may mediate relaxation through release of adenosine with subsequent actions on smooth muscle A2A receptors [31]. Hydrolysis of NAD could be investigated using selective inhibitors of CD38 (an NAD glycohydrolase), such as β-araF-NAD [35], but unfortunately these are not commercially available.
In the porcine mesenteric arteries, NAD caused an endothelium-independent vasoconstriction up to a concentration of 300 μM. The P2 receptor antagonist suramin had no significant effect on these contractile responses to NAD. αβ-meATP (a P2X receptor desensitising agonist) caused a significant attenuation of the NAD-evoked contractions, suggesting activation of suramin-insensitive P2X receptors. αβ-meATP has been shown to produce a transient contractile response in porcine mesenteric arteries, indicating that rapidly desensitising P2X1 receptors are expressed in these arteries [36]. The responses to NAD in the porcine mesenteric arteries, however, were not completely abolished in the presence of αβ-meATP. It was reported previously that rat large mesenteric arteries have a 25–100-fold lower sensitivity to αβ-meATP as an agonist than smaller arteries and are insensitive to suramin, which is not a characteristic of P2X1 receptors [37]. Moreover, NF449, a P2X1 receptor antagonist, had no significant effect on the contractile response to NAD. The fact that cumulative contractile response curves could be generated in the porcine mesenteric arteries indicated a relative lack of desensitisation, which is also inconsistent with NAD actions at rapidly desensitising P2X1 receptors. Since relatively large mesenteric arteries were used in this study, this could suggest that, as in the rat large mesenteric arteries, non-P2X1 receptors or heteromeric P2X receptors are involved, but the response could also involve P2Y receptors. In our porcine mesenteric artery segments, the pEC50 for NAD was 4.54. NAD has been identified as an agonist at P2X1, P2X4 and P2X7 receptors in human monocytes [22], and NADPH-mediated contractions of rat aorta were mediated via P2X1 receptors [38], but there are no reported potency estimates for NAD acting through P2X receptors in the literature.
In a number of cell types, extracellular NAD signals through conversion into cADPR, which in turn mobilises intracellular Ca2+ [12, 14, 39–41]. In human monocytes, exposure to NAD resulted in activation of P2X receptors and a rapid increase in [Ca2+]i caused by an influx of extracellular Ca2+ independent of intracellular Ca2+; the possibility of NAD acting through a degradation product was ruled out using selective inhibitors of CD38 and a stable NAD analogue [22, 42]. Pfister et al. analysed the NAD degradation products in human monocytes and found that NAD was mainly degraded to ADP ribose, nicotinamide and minor amounts of AMP, ADP and cADPR, but adenosine was not detected [43]. These metabolites can all have extracellular actions mediated through different purine receptors. Neither ADP nor AMP are agonists at P2X receptors, but ADP ribose (but not nicotinamide) mimicked the effect of NAD at P2X receptors in human monocytes [42]. Thus, ADP ribose, formed from metabolism of NAD, is potentially involved in the vasocontractile responses that we observed in the porcine mesenteric arteries.
Since multiple contractile and relaxant purine receptors are expressed on blood vessels, we hypothesised that the contractile actions of NAD in the porcine mesenteric artery could be augmented following antagonism of opposing NAD-mediated vasorelaxation. However, neither CGS15943 nor SCH58261 had a significant effect on the contractile response to NAD in the porcine mesenteric artery, indicating a lack of involvement of adenosine receptors. The reduction in NAD-evoked contraction which occurred at the highest concentration of NAD (1 mM) was blocked by suramin, suggesting that it may be mediated through P2 receptors. PaCoA did not mimic the effect of suramin, suggesting that NAD fails to act through P2Y1 receptors in this tissue, either directly, or for example, through metabolism to ADP ribose, which has been shown to be a ligand for the P2Y1 receptor [44].
The action of NAD in the porcine mesenteric artery is clearly different to its effects in in the rat aorta and porcine coronary artery, and also in mesenteric arteries in other species. We showed that adenosine produces concentration-dependent vasorelaxation of the porcine mesenteric artery, which is important because this shows that the pronounced differences in vasomotor responses of NAD are not simply due to a differential expression of adenosine receptors between the different blood vessels. In the rat mesenteric arterial bed, NAD did not elicit vasocontraction but caused vasorelaxation and inhibited sympathetic and sensory neurotransmission, effects which were antagonised by 8-para-sulfophenyltheophylline and potentiated in the presence of an adenosine deaminase inhibitor, indicating an involvement of adenosine and adenosine receptors [18]. These effects of NAD mimic the actions of adenosine in the rat mesenteric arterial bed indicating a likely involvement of postjunctional vasorelaxant A2B receptors and prejunctional A1 receptors, respectively [45, 46]. NAD also inhibited neuronal noradrenaline release in the dog mesenteric artery [8], an effect likely mediated via A1 adenosine receptors given their widespread occurrence prejunctionally on sympathetic perivascular nerves [3]. Thus, NAD actions in the porcine mesenteric artery appear to deviate from those in other arteries for reasons which are currently unclear. It would be interesting to carry out a comparative investigation of ectoenzyme expression of the porcine mesenteric and other arteries.
The most robust evidence for a physiological role for extracellular NAD is as a neurotransmitter in nerves of the gastrointestinal tract and bladder [11]. Whether this is relevant for the sparsely innervated rat thoracic aorta and porcine coronary artery remains to be determined, but it could be relevant for sympathetic neurotransmission in the porcine mesenteric artery; it would be interesting to determine whether there is a role for NAD as a neurotransmitter in porcine mesenteric arteries, as shown in canine mesenteric arteries [8]. Extracellular signalling of NAD could also be important in pathophysiological conditions leading to cell lysis during tissue injury and inflammation, conditions which are known to release other purine nucleotides [3].
In conclusion, there are clear differences in the actions of NAD in the porcine mesenteric artery compared with the rat thoracic aorta and porcine coronary artery; in both rat thoracic aorta and porcine coronary artery, NAD mediates relaxation through P1 receptors, while in porcine mesenteric artery, NAD causes contraction which appears to involve P2 receptors whose identity remains to be defined. The suggestion that NAD may be broken down to adenosine to act through P1 receptors may indicate higher levels of ectonucleotidases in rat thoracic aorta and porcine coronary artery than in the porcine mesenteric artery.
References
Ralevic V, Burnstock G (1998) Receptors for purines and pyrimidines. Pharmacol Rev 50:413–492
Ralevic V (2009) Purines as neurotransmitters and neuromodulators in blood vessels. Curr Vasc Pharmacol 7(1):3–14
Burnstock G, Ralevic V (2014) Purinergic signaling and blood vessels in health and disease. Pharmacol Rev 66(1):102–192
Ralevic V (2012) P2X receptors in the cardiovascular system. WIREs Membr Transp Signal 1(5):663–674
Ziegler M (2000) New functions of a long-known molecule emerging roles of NAD in cellular signaling. Eur J Biochem 267(6):1550–1564
Ziegler M (2005) A vital link between energy and signal transduction. FEBS J 272(18):4561–4564
Lee HC (2001) Physiological functions of cyclic ADP-ribose and NAADP as calcium messengers. Annu Rev Pharmacol Toxicol 41:317–345
Smyth LM, Bobalova J, Mendoza MG, Lew C, Mutafova-Yambolieva VN (2004) Release of β-nicotinamide adenine dinucleotide upon stimulation of postganglionic nerve terminals in blood vessels and urinary bladder. J Biol Chem 279(47):48893–48903
Breen LT, Smyth LM, Yambolieva IA, Mutafova-Yambolieva VN, (2006) β-NAD is a novel nucleotide released on stimulation of nerve terminals in human urinary bladder detrusor muscle. Am J Physiol Renal Physiol 290(2):F486–95
Mutafova-Yambolieva VN, Hwang SJ, Hao X, Chen H, Zhu MX, Wood JD, Ward SM, Sanders KM (2007) β-nicotinamide adenine dinucleotide is an inhibitory neurotransmitter in visceral smooth muscle. Proc Natl Acad Sci U S A 104(41):16359–16364
Mutafova-Yambolieva VN (2012) Neuronal and extraneuronal release of ATP and NAD+ in smooth muscle. IUBMB Life 64(10):817–824
De Flora A, Zocchi E, Guida L, Franco L, Bruzzone S (2004) Autocrine and paracrine calcium signaling by the CD38/NAD+/cyclic ADP-ribose system. Ann N Y Acad Sci 1028:176–191
Bruzzone S, Guida L, Zocchi E, Franco L, De Flora A (2001) Connexin 43 hemichannels mediate Ca2+-regulated transmembrane NAD fluxes in intact cells. FASEB J 15(1):10–12
Romanello M, Bicego M, Pirulli D, Crovella S, Moro L, D’Andrea P (2002) Extracellular NAD+: a novel autocrine/paracrine signal in osteoblast physiology. Biochem Biophys Res Commun 299(3):424–431
Nikiforov A, Dölle C, Niere M, Ziegler M (2011) Pathways and subcellular compartmentation of NAD biosynthesis in human cells: From entry of extracellular precursors to mitochondrial NAD generation. J Biol Chem 286(24):21767–21778
Burnstock G, Hoyle CH (1985) Actions of adenine dinucleotides in the guinea-pig taenia coli: NAD acts indirectly on P1-purinoceptors; NADP acts like a P2-purinoceptor agonist. Br J Pharmacol 84(4):825–831
Hoyle CH (1990) Pharmacological activity of adenine dinucleotides in the periphery: possible receptor classes and transmitter function. Gen Pharmacol 21(6):827–831
Ralevic V, Hoyle CH, Burnstock G (1995) Pivotal role of phosphate chain length in vasoconstrictor versus vasodilator actions of adenine dinucleotides in rat mesenteric arteries. J Physiol 483(Pt 3):703–713
Cosker F, Cheviron N, Yamasaki M, Menteyne A, Lund FE, Moutin MJ, Galione A, Cancela JM (2010) The ecto-enzyme CD38 is a nicotinic acid adenine dinucleotide phosphate (NAADP) synthase that couples receptor activation to Ca2+ mobilization from lysosomes in pancreatic acinar cells. J Biol Chem 285(49):38251–38259
Berthelier V, Tixier JM, Muller-Steffner H, Schuber F, Deterre P (1998) Human CD38 is an authentic NAP(P)+ glycohydrolase. Biochem J 330(Pt 3):1383–1390
Moreschi I, Bruzzone S, Nicholas RA, Fruscione F, Sturla L, Benvenuto F, Usai C, Meis S, Kassack MU, Zocchi E, De Flora A (2006) Extracellular NAD+ is an agonist of the human P2Y1 purinergic receptor in human granulocytes. J Biol Chem 281(42):31419–31429
Grahnert A, Klein C, Hauschildt S (2009) Involvement of P2X receptors in the NAD+-induced rise in [Ca2+]i in human monocytes. Purinergic Signal 5(3):309–319
Alefishat E, Alexander SP, Ralevic V (2013) Antagonism of P2Y1-induced vasorelaxation by acyl CoA: a critical role for palmitate and 3′-phosphate. Br J Pharmacol 168(8):1911–1922
Hansmann G, Bültmann R, Tuluc F, Starke K (1997) Characterization by antagonists of P2-receptors mediating endothelium-dependent relaxation in the rat aorta. Naunyn Schmiedebergs Arch Pharmacol 356(5):641–652
Rose’Meyer RB, Hope W (1990) Evidence that A2 purinoceptors are involved in endothelium-dependent relaxation of the rat thoracic aorta. Br J Pharmacol 100(3):576–580
Balwierczak JL, Sharif R, Krulan CM, Field FP, Weiss GB, Miller MJ (1991) Comparative effects of a selective adenosine A2 receptor agonist, CGS 21680, and nitroprusside in vascular smooth muscle. Eur J Pharmacol 196(2):117–123
Merkel LA, Lappe RW, Rivera LM, Cox BF, Perrone MH (1992) Demonstration of vasorelaxant activity with an A1-selective adenosine agonist in porcine coronary artery: involvement of potassium channels. J Pharmacol Exp Ther 260(2):437–443
Monopoli A, Conti A, Zocchi C, Casati C, Volpini R, Cristalli G, Ongini E (1994) Pharmacology of the new selective A2a adenosine receptor agonist 2-hexynyl-5′-N-ethylcarboxamidoadenosine. Arzneimittelforschung 44(12):1296–1304
Abebe W, Makujina SR, Mustafa SJ (1994) Adenosine receptor-mediated relaxation of porcine coronary artery in presence and absence of endothelium. Am J Physiol 266(5 Pt 2):H2018–H2025
Olivecrona GK, Gotberg M, Harnek J, Wang L, Jacobson KA, Erlinge D (2004) Coronary artery reperfusion: the ADP receptor P2Y1 mediates early reactive hyperemia in vivo in pigs. Purinergic Signal 1(1):59–65
Rayment SJ, Latif ML, Ralevic V, Alexander SPH (2007) Evidence for the expression of multiple uracil nucleotide-stimulated P2 receptors coupled to smooth muscle contraction in porcine isolated arteries. Br J Pharmacol 150(5):604–612
Coddou C, Loyola G, Boyer JL, Bronfman M, Huidobro-Toro JP (2003) The hypolipidemic drug metabolites nafenopin-CoA and ciprofibroyl-CoA are competitive P2Y1 receptor antagonists. FEBS Lett 536(1–3):145–150
Manolopoulos P, Glenn JR, Fox SC, May JA, Dovlatova NL, Tang SW, Thomas NR, Ralevic V, Heptinstall S (2008) Acyl derivatives of coenzyme A inhibit platelet function via antagonism at P2Y1 and P2Y12 receptors: a new finding that may influence the design of anti-thrombotic agents. Platelets 19(2):134–145
Stone TW (1981) Actions of adenine dinucleotides on the vas deferens, guinea-pig taenia caeci and bladder. Eur J Pharmacol 75(2–3):93–102
Muller-Steffner HM, Malver O, Hosie L, Oppenheimer NJ, Schuber F (1992) Slow-binding inhibition of NAD+ glycohydrolase by arabino analogues of β-NAD+. J Biol Chem 267(14):9606–9611
Shatarat, A., Dunn, W.R., Ralevic, V., Raised tone reveals ATP as a sympathetic neurotransmitter in the porcine mesenteric arterial bed. Purinergic Signalling, in press
Gitterman DP, Evans RJ (2000) Properties of P2X and P2Y receptors are dependent on artery diameter in the rat mesenteric bed. Br J Pharmacol 131(8):1561–1568
Judkins CP, Sobey CG, Dang TT, Miller AA, Dusting GJ, Drummond GR (2006) NADPH-induced contractions of mouse aorta do not involve NADPH oxidase: a role for P2X receptors. J Pharmacol Exp Ther 317(2):644–650
Sun L, Adebanjo OA, Moonga BS, Corisdeo S, Anandatheerthavarada HK, Biswas G, Arakawa T, Hakeda Y, Koval A, Sodam B, Bevis PJ, Moser AJ, Lai FA, Epstein S, Troen BR, Kumegawa M, Zaidi M (1999) CD38/ADP-ribosyl cyclase: a new role in the regulation of osteoclastic bone resorption. J Cell Biol 146(5):1161–1172
Verderio C, Bruzzone S, Zocchi E, Fedele E, Schenk U, De Flora A, Matteoli M (2001) Evidence of a role for cyclic ADP-ribose in calcium signalling and neurotransmitter release in cultured astrocytes. J Neurochem 78(3):646–657
Esguerra M, Miller RF (2002) CD38 expression and NAD+-induced intracellular Ca2+ mobilization in isolated retinal Muller cells. Glia 39(3):314–319
Gerth A, Nieber K, Oppenheimer NJ, Hauschildt S (2004) Extracellular NAD+ regulates intracellular free calcium concentration in human monocytes. Biochem J 382(Pt 3):849–856
Pfister M, Ogilvie A, da Silva CP, Grahnert A, Guse AH, Hauschildt S (2001) NAD degradation and regulation of CD38 expression by human monocytes/macrophages. Eur J Biochem 268(21):5601–5608
Gustafsson AJ, Muraro L, Dahlberg C, Migaud M, Chevallier O, Khanh HN, Krishnan K, Li N, Islam MS (2011) ADP ribose is an endogenous ligand for the purinergic P2Y1 receptor. Mol Cell Endocrinol 333(1):8–19
Rubino A, Ralevic V, Burnstock G (1993) The P1-purinoceptors that mediate the prejunctional inhibitory effect of adenosine on capsaicin-sensitive nonadrenergic noncholinergic neurotransmission in the rat mesenteric arterial bed are of the A1 subtype. J Pharmacol Exp Ther 267(3):1100–1104
Rubino A, Ralevic V, Burnstock G (1995) Contribution of P1-(A2b subtype) and P2-purinoceptors to the control of vascular tone in the rat isolated mesenteric arterial bed. Br J Pharmacol 115(4):648–652
Acknowledgement
The support of the University of Jordan is gratefully acknowledged
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article

Cite this article
Alefishat, E., Alexander, S.P.H. & Ralevic, V. Effects of NAD at purine receptors in isolated blood vessels. Purinergic Signalling 11, 47–57 (2015). https://doi.org/10.1007/s11302-014-9428-1
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11302-014-9428-1