
Abstract
Genetic variability of modulus of elasticity (MOE) was investigated in three genetic trials, including two progeny (16 years old) and one clonal (19 years old) trials of hybrid larch (Larix x eurolepis Henry). MOE was directly assessed on standing trees using the Rigidimeter, a bending device, and related to other traits including height, BH diameter and wood density. Mean MOE ranged from 5,183 to 9,228 MPa among families in the progeny trials and from 4,591 to 11,486 MPa in the clonal trial. Among traits studied, MOE was one of the most variable traits. It was strongly and positively related to wood density at both the individual and genotype mean levels. Interestingly too, wood stiffness did not seem, or only weakly, unfavourably linked to stem diameter at the phenotypical level, but it was negatively or not correlated to diameter at the genetic level. As well, MOE showed a high GxE stability over the two progeny trial sites. Narrow-sense heritabilities for MOE were moderate (around 0.36). In all three trials, they were lower than those for wood density or total height, and of the same level as for diameter. Improvement of wood stiffness of hybrid larch using the Rigidimeter seems possible and promising genetic gains are expected. Impacts of selection for growth traits on MOE are also discussed.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Hybrid larchFootnote 1 (Larix x eurolepis Henry) frequently exhibits a superior growth over a large range of environments, when compared to pure parental species (namely European larch (Larix decidua Mill.) and Japanese larch (Larix kaempferi (Lamb.) Carr.)) (Langner and Schneck 1998; Pâques 2000). This makes it particularly attractive to foresters. However, the impact of enhanced growth on wood properties has become a major concern among tree breeders as the proportion of juvenile wood characterised by lower mechanical properties is usually increased due to shorter rotations. Their aim is therefore to improve wood quality traits or at least not to alter some key wood properties through the selection process.
Wood properties of hybrid larch were shown to be either inferior (e.g. proportion of latewood: Nanson and Sacré 1978; Reck 1980; fibre thickness: Nanson and Sacré 1978), intermediate (e.g. shrinkage: Reck 1980; heartwood proportion: Pâques and Rozenberg 1995) or even superior (e.g. extractives content: Reck 1980) to pure species. For wood density as well, variable results have been observed with either inferior (Reck 1977), intermediate (Nanson and Sacré 1978; Bastien and Keller 1980) or superior (Langner and Reck 1966; Reck 1980; Pâques and Rozenberg 1995) performances of the hybrid compared to European and Japanese larches. Such apparent antagonisms are very often linked to specific hybrid combinations. In any case, a large enough genetic variability exists among hybrids, which should allow selection of outstanding hybrids, superior or at least equal to the best parent.
Breeders’ interest for mechanical properties such as stiffness is increasing, especially at the final phase of the selection process. Indeed, stiffness, for example, is one of the key properties for end-use products requesting structural quality. It is itself related to basic wood properties such as density and microfibril angle, the measurements of which are costly and highly variable within trees. However, few results about variability and genetic control level of stiffness and other mechanical properties have been published. This is explained by the heavy labour and high cost requested by standardised laboratory tests (such as NF B51-016 French standard) to evaluate modulus of elasticity (MOE). In addition, the destruction of trees imposed for specimen sampling is rarely compatible with breeders’ objectives.
In larch, Bastien and Keller (1980) observed a rather large superiority for static MOE of the hybrid compared to European (around 27%) and Japanese (nearly 33%) larch in a 15-year-old trial. In the same trial some 20 years later, Pâques and Rozenberg (1995) confirmed this superiority: around 23% over European larch and nearly 15% over Japanese larch. But besides these rare results at the mean species level, even fewer results are available on the genetic variability and heritability of this character for larch.
The development of non-destructive methods to evaluate MOE directly on standing trees now allows routine assessment and, thereby, genetic studies which are based on rather large datasets. Bending tests, as well as acoustic methods, are the most common technologies (e.g. Koizumi and Ueda 1986; Jacques et al. 2004; Matheson et al. 2008). For example, Koizumi et al. (1990b) used a bending test device in a half-sib progeny trial of Japanese larch to estimate heritability of stiffness. More recently, Jacques et al. (2004) and Fujimoto et al. (2006) have tested acoustic methods on standing trees to estimate MOE heritability in clonal and progeny trials of hybrid larch (L. x eurolepis and Larix gmelinii x L. kaempferi, respectively).
In a previous paper (Pâques and Rozenberg 2009), we showed that the Rigidimeter—a device inspired from Koizumi and Ueda’s (1986) bending test equipment and developed by Launay et al. (2000)— was a reliable tool to estimate MOE from standing trees and of practical interest for tree breeders as it allowed the proper ranking of genotypes for MOE when compared to laboratory results.
In this paper, we present genetic parameters for wood stiffness estimated in different hybrid larch trials and the levels of expected gains in MOE from selection using the Rigidimeter.
Material and methods
Plant material
Hybrid larch trees from two progeny trials and from one clonal trial were used in this study. The progeny trials are located at Beaumont-du-Lac in the Western Massif Central Mountain and at Baume-Les-Dames in the Jura Mountain; the clonal trial is established at Eclache in the central part of the Massif Central Mountain. Both progeny trials were planted in early spring 1985 using 2-year-old bare-root seedlings while the clonal trial was planted during spring 1984 with 2-year-old bare-root stecklings. A common spacing of 3 × 3 m was used in all three plantations. A short description of the three experiments and of the experimental designs is summarised in Table 1.
The progeny trials consisted of several full-sib families created through crosses of several European larches from the Sudetan Mountains near the border of the Czech Republic and Poland used as seed parents, and several Japanese larches from various locations used as pollen parents. The clonal trial included one hundred clones, the ortets of which were selected for total height at nursery stage (2 years old) from several hybrid larch full-sib families.
Measurements
MOE was indirectly evaluated on standing trees using the Rigidimeter (see Launay et al. 2000 and Pâques and Rozenberg 2009 for a description). MOE was evaluated in the same two perpendicular cardinal directions on each tree within a trial (excepted in Beaumont-du-Lac, only one direction). The bending deviation of trees due to the applied force was measured four times after complete unloading of the force created by the device. MOEs were computed and averaged over the last three replicates. At the same time, BH diameter over bark was carefully measured at the same height (1.30 m) as where bending deviation was measured. Total height of each tree was recorded as well.
These measurements took place during winter in Beaumont-du-Lac and in Baume-Les-Dames, when trees were, respectively, 16 and 19 years old from seed. In Eclache, trees were measured in early spring before bud flushing when trees were 19 years old from cutting.
A total of 268 and 222 trees were measured in Beaumont-du-Lac and in Baume-Les-Dames from, respectively, 25 and 23 different families (that is around 10.7 and 9.7 trees per family, respectively). Seventeen families were common to the two sites. In Eclache, 157 trees were assessed from 32 different clones (that is around 4.9 ramets per clone). Trees were randomly chosen within progenies in Beaumont-du-Lac, but in Baume-Les-Dames and in Eclache, trees were sampled around the progeny/clone means for BH diameter because upper-diameter classes of several progenies/clones exceeded the upper limit recommended for use of the Rigidimeter (28 cm). In addition, in Eclache, only one ramet per clone and per block was measured.
MOE data were related to growth measurements: height (HT), BH diameter (D) and individual tree volume (V) computed following Thill and Palm (1984). They were also related to the overall wood density (De) estimated through X-ray microdensitometry on increment cores collected on the same trees at 1 m above ground.
Statistical analysis
Analysis of variance was conducted on data with the following model:
where
- bi :
-
random effect of block i
- ge j :
-
random effect of genotype (family or clone) j
- \( \sigma_{ijk}^2 \) :
-
residual effect
Variance components were estimated using the S + -varcomp procedure using the REML option (Anonymous 1995). Narrow-sense heritabilities were then computed for each trait as:
where
- \( \sigma_{\text{A}}^2 \) :
-
2 \( \sigma_{\text{fa}}^2 \) with \( \sigma_{\text{fa}}^2 \) = family variance
- \( \sigma_{\text{p}}^2 \) :
-
phenotypic variance
where \( \sigma_{\text{G}}^2 = \sigma_{\text{cl}}^2 \), the clone variance
Broad-sense heritabilities were computed as:
where σ 2 p fa, σ 2 p cl = phenotypic variance at the family/clone mean level. Standard deviations of the genetic parameters were estimated using the Jacknife method developed in the Diogene software (Baradat et al. 1995).
Expected genetic gains from family (clone) selection were estimated from the above parameters with a selection intensity arbitrarily fixed at 5%, and the efficiency of indirect selection was calculated according to Falconer (1981).
In addition, type-B genetic correlations and Spearman rank correlations were computed across progeny trials, based on the 17 common progenies, in order to evaluate the importance of GxE interactions and progeny rank changes. Type-B correlations were computed following Burdon (1977):
where
- rfaxfay:
-
correlation between family means at sites x and y
- h2fa x , h2fa y :
-
heritabilities of family means at sites x and y
The main advantage of these methods is that they avoid the constraints from other methods relying on analysis of variance, in particular, homogeneity of residual variances across environments.
Results and discussion
Overall mean performances
While characterised by contrasting environmental conditions, the three sites clearly offered favourable growth conditions for hybrid larch. Survival was excellent (over 84%) except in Baume-Les-Dames, where mortality reached 23% due to plantation shock following a severe summer drought in 1985. As can be derived from Table 2, the mean annual height increment from the time of planting reached 76 cm in Eclache and over 97 cm in the two other sites. In all three sites, the mean annual ring width exceeded 6.3 mm on average.
Average MOE was very similar in the three experiments, with an average of 7,181 MPa in Beaumont-du-Lac, 7,368 MPa in Eclache and 7,353 MPa in Baume-Les-Dames. The mean wood density ranged from 412 kg/m3 in Beaumont-du-Lac up to 485 kg/m3 in Baume-Les-Dames.
Comparison with other published results is rather difficult. Indeed, in addition to a large variety of biological material (species, provenances, etc.) and ages, the studies used different types of wood specimens (from small normalised clear-wood specimen to logs or even boles) and methodologies (three- to four-point static bending, ultrasound, resonance frequency, etc). However, if one keeps in mind, the higher wood stiffness of standard specimen compared to that of non-normalised samples for a given testing methodology and the increase of stiffness from pith to bark and, thus, with age as shown, for example, in Pâques and Rozenberg (2009) and by several authors (e.g. Bendtsen and Senft 1986; Leban and Haines 1999; Brüchert et al. 2000; Fujimoto et al. 2006; Ivkovic et al. 2009), our results look mostly coherent with published results.
Using normalised specimen and the four-point static bending test, Dinelli (1992) found a mean MOE of 6,326 MPa (4,485–7,678) in a 10-year-old progeny test of hybrid larch; it reached 8,000 MPa (6,500–9,700) for 14-year-old hybrid clones (Jacques 2003; Jacques et al. 2004) and 9,330 MPa for 15-year-old trees from a hybrid larch variety (Deret and Keller 1979). Higher MOE are usually cited for older larch trees, exceeding 10,000 MPa (e.g. Ringard 1980; Leban and Haines 1999; CTBA 2000; Nakada 2002).
More closely comparable are results directly obtained from bending tests on standing trees using a special device developed by Koizumi and Ueda (1986). In a 18-year-old progeny trial of Japanese larch, Koizumi et al. (1990a) observed a mean MOE value of around 9,140 MPa (standard deviation (SD) = 1,343). For 25–30-year-old Japanese larch clones, Koizumi et al. (1990b) reported average MOE of 9,530 (SD = 1340) – 9,790 MPa (SD = 1630) according to testing sites. In 27–35-year-old Japanese larch plots, Koizumi and Ueda (1987) recorded MOE ranging from 7,280–8,580 MPa. Most values reported in these papers were higher than those found on average in this study. But as observed by Koizumi and Ueda (1987), MOE steeply increases between 10 and 20 years of age: for example, in a same site, mean MOE passed from less than 5,000 MPa at 9 years old up to 11,750 MPa at 27 years old.
Individual and mean genotype variability
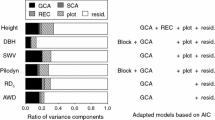
Consistently over sites, stiffness was among the most variable traits, both at the individual and at the family/clone levels. Coefficients of variation for MOE ranged from 22% to 36% at the individual level (CVi) and from 11% to 20% at the mean family/clone (CVfa/cl) level. They were comparable to those for stem volume (CVi = 23–27% and CVfa/cl = 11–15%) but two to three times larger than those of the other traits. Total height and wood density showed the lowest variability.
Significant differences among genotypes (families or clones) were found in the two progeny tests for all traits. For stiffness, the best progeny had a mean MOE of 8,809 MPa in Beaumont-du-Lac against 5,183 MPa for the weakest one; in Baume-Les-Dames, MOE for the best one reached on average nearly 9,300 and 5,700 MPa for the weakest one. In Eclache, the clonal range is even wider with the highest MOE value reaching 11,486 MPa and the lowest, 4,591 MPa, but the level of significance for clonal differences was lower (p = 0.067 for MOE) due to the low average number of ramets/clone.
Wood stiffness of larch is apparently characterised by a larger variability compared to wood density or even growth traits. For example, as in our study, Koizumi et al. (1990b) found coefficients of variation at the individual tree level of 14.1–16.7% for MOE, compared to 8.5–10.1% for BH diameter and 7.3–7.9% for wood density. As well, Jacques (2003) observed the same trends at the clone mean level with CVclMOE = 13.5% > CVclBH girth = 7.3% > CVclwood density = 5.3%. In the same study in three other clonal trials, he found as well that MOE had the largest variability (CVcl = 11.1–17.8%), together with BH diameter (CVcl = 12.8–19%), well above that observed for total height (CVcl = 7.5–11.2%) and density (CVcl = 5.9–8.7%).
Phenotypic correlations of MOE with other traits
Over the three sites, trunk stiffness was positively correlated with wood density at the individual phenotypic level but even more strongly at the genotype mean level (Table 3). In contrast, correlation coefficients of MOE with both BH diameter and volume were either positive or negative but never significantly different from 0 in both progeny trials, except in Baume with diameter. In the clonal trial, a positive correlation, significantly different from 0, was observed at the individual level, but a negative correlation (only significantly different from 0 with V) was observed at the mean clonal level. A positive correlation (highly significantly different from 0 at the individual level) was observed between MOE and total height.
Interestingly, larch wood stiffness does not seem, or is only weakly, unfavourably correlated with stem growth. Many results are becoming available and confirm this major trend in larch wood. Several authors have already noticed the little effect of growth rate on MOE, like Koizumi and Ueda (1987) for Japanese larch, CTBA (2000) for mature European larches (r = −0.14 ns) and Dinelli (1992) and Jacques (2003) for young hybrid larches (r = −0.40 ns). At the clone/family mean level as well, Nakada (2002) and Koizumi et al. (1990b) found no correlation significantly different from 0 between MOE and DBH. Yet, some opposite results at the individual tree level were also given by Miyajima and Hasegawa (1978, 37 years old) and Koizumi et al. (1990a) (r = −0.28**) for Japanese larch, by Jacques (2003) on young hybrid larch (r = −0.31* − 0.35**) and by Leban and Haines (1999) for a single seed orchard progeny of hybrid larch (33 years old). But in several of these studies, genetic correlations were then not significant (Koizumi et al. 1990b; Jacques 2003).
The absence or weakness of links between clearwood MOE and diameter growth (or weak negative links between indirect MOE estimation on standing trees and DBH) is not unique for larch: it was also reported for some other species like 13-year-old Douglas-firs (Pseudotsuga menziesii) (Mamdy et al. 1999; Launay et al. 2000) and 13–14-year-old radiata pines (Pinus radiata) (Kumar 2004). Yet, the opposite trend is usually found for these species and others at the individual phenotypic level. Nepveu and Blachon (1989) observed a clear decrease of stiffness with the increase of ring width for Douglas-fir (Pseudotsuga menziesii) Sitka (Picea sitchensis) and Norway (Picea abies) spruces, maritime (Pinus pinaster) and Scots (Pinus sylvestris) pines and fir (Abies sp.) For example, MOE dropped from more than 16,000 MPa down to less than 11,000 MPa when ring width increased from 3 to 10 mm in a population of 36–80-year-old Douglas-fir trees. Johnson et al. (2005) reported a correlation as high as 0.77 between MOE and the number of rings per centimetre in Douglas-fir (20–28 years old).
Genetic correlations
Additive genetic and genotypic correlation coefficients between MOE and diameter and volume appeared mostly strongly negative at Baume-Les-Dames and Eclache but surprisingly not in Beaumont, where they were found not significant (Table 3). Figure 1 showed the variable link between MOE and BH diameter at the progeny mean level in Beaumont and Baume-Les-Dames. MOE was positively (significantly or not) linked to total height. As for phenotypic correlations, strong positive links were consistently found between MOE and wood density.

Plot of progeny means for BH diameter (D) and MOE in Beaumont (left) and Baume-Les-Dames (right)
Jacques (2003) found, in a hybrid larch clonal trial (age, 14 years old), negative genotypic correlation coefficients between MOE and girth, ring width and volume; −0.57, −0.53 and −0.33, respectively. In three other clonal trials of similar ages, this author also observed negative genotypic correlations (−0.26, −0.41 and −0.38) between girth and speed of ultrasounds measured by Sylvatest for indirect MOE assessment. But in all these trials, none of the correlation coefficients was significant. Additionally, Fujimoto et al. (2006) found only a close-to-zero negative genetic correlation (−0.08) in L. gmelinii x L. kaempferi hybrids.
Negative, but also non-significantly different from 0, genetic correlations coefficients between wood stiffness and diameter have been found also in radiata pine (Matheson et al. 2008), while a strong negative correlation (−0.51) was observed by Johnson and Gartner (2006) in Douglas-fir.
As in our study, high genotypic correlations were found between MOE and wood density: 0.88 and 0.79, respectively, by Jacques (2003) for L. x eurolepis hybrids and by Fujimoto et al. (2006) for L. gmelinii x L. kaempferi hybrids. Moderate to high genetic correlations were also usually observed for several other species, like Douglas-fir and radiata pine (Kumar 2004; Johnson and Gartner 2006).
Inter-site stability among the two progeny trials
Type-B genetic correlations were computed in Beaumont and Baume-Les-Dames, based on the 17 common progenies (Table 4). The high level of type-B correlations for height, stiffness and density contrasted with the moderate to low level for other growth traits (diameter and volume), indicating a low level of GxE interaction, in particular for MOE.
Moreover, a correct ranking of the genotypes over the two sites was revealed for MOE (Fig. 2) and total height as revealed also by quite high Spearman rank correlations; in contrast, ranking changes looked important for D and V. Besides a strong site effect, this strong interaction effect for D and V may result from the rather high mortality already mentioned in Baume-Les-Dames. This has created contrasting inter-tree competition between sites, to which BH diameter is well-known to be more sensitive than total height.

MOE progeny means in Beaumont and Baume-Les-Dames progeny trials
Very few published results exist on genotype stability over sites for wood stiffness. Jacques (2003) observed a highly significant GxE interaction for MOE estimated by Sylvatest for clones over three sites and weak Pearson correlation coefficients (0.10–0.48) between clone means over sites. Nevertheless, it seemed that only three out of the 17 clones were responsible for this interaction.
In other species, high genetic correlations (or type-B genetic correlations) for stiffness have been found between sites as in our study (e.g. Kumar 2004; Matheson et al. 2008 for radiata pine; Johnson and Gartner 2006 for Douglas-fir).
Genetic control of traits—heritability
Moderate and equal narrow-sense heritabilities (0.36) were observed in the two progeny trials for MOE (Table 5). They were weaker than those for total height but mostly comparable or higher than heritabilities for BH diameter and wood density. In Eclache clonal trial, narrow-sense heritability for MOE was greater than that for diameter, close to that for volume but smaller than for total height and wood density. Narrow-sense heritabilities estimated in the two progeny trials were probably over-estimated as we neglected non-additive variance components. Indeed, in contrast to Fujimoto et al. (2006), who found that MOE was strictly under additive effects, we showed in a previous study that the non-additive variance component was of the same size as the additive one (Pâques 2004). But in both studies, the mating designs used were not optimum to estimate non-additive effects. This aspect should be further investigated in the future.
However, the level of heritability for MOE observed in the two progeny trials was consistent with that found by Koizumi et al. (1990a) on standing trees in a 18-year-old half-sib progeny trial of Japanese larch (\( h_{\text{ns}}^2 = 0.41 \)). It was close to static MOE heritability found by Fujimoto et al. (2006) on clearwood specimen (0.44) but smaller than that for dynamic MOE obtained on logs (0.61). For the other traits, heritabilities were mostly within the range of heritability confidence limits reported by Stener et al. (2002) for Larix x eurolepis for diameter (0.11–0.49 and 0.19–0.65, respectively, between 18 and 21 years old) and for total height (0.16–0.52 and 0.11–0.47, respectively).
For clonal material, heritabilities instead looked comparable or in the lower range of published results, whatever the traits. For example, Jacques (2003) reported for young hybrid larch around 16 years old, heritabilities over four sites in the range of 0.49–0.74 for MOE (measured on specimen by static bending or on standing trees by Sylvatest ultrasound speed), 0.38–0.60 for total height, 0.24–0.49 for BH girth, 0.53 for volume (one site only) and 0.43–0.71 for density. But, due to the low number of clones and ramets/clone, these heritabilities were accompanied by large SDs and our values fitted well within their confidence limits.
Expected genetic gains
Promising genetic gains are expected from family or clone selection for MOE: indeed, if, for example, the 5% best genotypes were selected (that is i = 2.063), relative gains would rise up to 12%–19% in the progeny trials and to 30% in the clonal trial (Table 6). Direct selection for MOE also proved to be mostly more efficient than indirect selection for a correlated trait, namely for wood density, except in Beaumont. Compared to indirect selection, direct selection for MOE would bring an extra gain of around 370 MPa in Baume and 670 MPa in Eclache.
Other technical or financial considerations might favour indirect selection, for example, if the assessment of the correlated trait is easier, faster or cheaper than that of the target trait. This is not really the case for wood density but it could be for tools like pilodyn, which proved reliable to indirectly estimate density. Still, in advanced selection steps, direct selection for MOE on standing trees proved efficient, and this property—well correlated with standard MOE (Pâques and Rozenberg 2009)—is closer to properties of interest for structural products.
The goal of tree breeding is often to increase growth while simultaneously improving or at least maintaining original wood properties. It is therefore of interest to evaluate the impact of selection for growth traits on wood stiffness. Results varied from trait to trait (Table 7). Selection for total height had no or a slight positive effect on MOE (relative gains from 1% to 8%), whereas selection for BH diameter strongly affected MOE (relative loss from −15% in Baume up to −26% in Eclache), except in Beaumont (+2%). Overall, selection for stem volume would reduce MOE by 11% to 13% in Baume and Eclache, respectively, but increase it by 5% in Beaumont.
Such a negative impact of selection for diameter on MOE has been found by several authors (e.g. Fujimoto et al. 2006 on L. gmelinii x L. kaempferi hybrids, Johnson and Gartner 2006 on Douglas-fir), while some positive responses have also been found (e.g. Cherry et al. 2008 on Douglas-fir). As in our study, Johnson and Gartner (2006) showed a stronger negative impact on MOE when selecting for diameter than for height.
While expected gains for stiffness reported in this study are promising, only scarce information is available for hybrid larch on juvenile–mature correlations for wood stiffness. Fujimoto et al. (2006) showed that static MOE of inner and outer wood specimen were highly correlated (rg = 0.63), a result confirmed by Pâques and Rozenberg (2009), who found a genetic correlation of 0.90. However, the question of the efficiency of such an early testing and selection for wood stiffness remains mostly open.
Conclusions
In a previous paper, we showed that the Rigidimeter was a reliable tool to non-destructively evaluate wood stiffness right on standing trees. In addition, it proved to be easy enough to handle so that it can be used routinely in genetic trials for MOE assessment.
In this paper, we showed that a large genetic variability was available in hybrid larch both at the family and at the individual tree levels for MOE together with high levels of heritabilities. Moreover, ranking of genotypes was mostly conserved over sites. Hence, improvement of wood stiffness through selection with the Rigidimeter looked possible with high expected genetic gains, and it seemed more efficient than with indirect selection for wood density. The impact of selection for growth traits on MOE is still uncertain since the links between these traits had variable trends according to sites in this study. More information on genetic correlations between growth and stiffness is still needed. Additionally, the level of correlation between juvenile and mature wood MOE must be further studied.
Notes
Hybrid larch will explicitly refer to Larix x eurolepis unless specified
References
Anonymous (1995) S-Plus, guide to statistical and mathematical analysis. StatSci Division, MathSoft, Seattle
Baradat P, Labbé T, Bouvet JM (1995) Conception d’index pour la sélection réciproque récurrente. Aspects génétiques, statistiques et informatiques. In: Traitements statistiques des essais de sélection, Ed CIRAD, pp 101–150
Bastien J-C, Keller R (1980) Intérêts comparés du mélèze hybride (Larix x eurolepis Henry) avec les deux espèces parentes. Rev For Fr XXXII:521–530
Bendtsen BA, Senft J (1986) Mechanical and anatomical properties in individual growth rings of plantation-grown eastern cottonwood and loblolly pine. Wood Fiber Sci 18(1):23–38
Brüchert F, Becker G, Speck T (2000) The mechanics of Norway spruce (Picea abies (L.) Karst): mechanical properties of standing trees from different thinning regimes. For Ecol Manage 135:45–62
Burdon RD (1977) Genetic correlation as a concept for studying genotype–environment interaction in forest tree breeding. Silvae Genet 26(5–6):168–175
Cherry ML, Vikram V, Briggs D, Cress DW, Howe GT (2008) Genetic variation in direct and indirect measures of wood stiffness in coastal Douglas-fir. Can J For Res 38:2476–2486
CTBA (2000) Classification du Mélèze d’Europe. Norme et structure en bois. Rapport final, CTBA Bordeaux
Deret E, Keller R (1979) Etude comparative du bois de mélèze d’Europe (Larix decidua Mill.), de mélèze du Japon (Larix leptolepis Gord.) et de leur hybride (Larix x eurolepis Henry). INRA Nancy, doc n° 1979/3
Dinelli B (1992) Etude de quelques paramètres de la qualité du bois de mélèze hybride. Rapport de stage, ESEM Orléans
Falconer DS (1981) Introduction to quantitative genetics. Longman, London
Fujimoto T, Akutsu H, Nei M, Kita K, Kuromaru M, Oda K (2006) Genetic variation in wood stiffness and strength properties of hybrid larch (Larix gmelinii var. japonica x L.kaempferi). J For Res 11:343–349
Ivkovic M, Gapare WJ, Abarquez A, Ilic J, Powell MB, Wu HX (2009) Prediction of wood stiffness, strength, and shrinkage in juvenile wood of radiata pine. Wood Sci Technol 43:237–257
Jacques D (2003) Amélioration du module d’élasticité du bois de Mélèze hybride (Larix x eurolepis Henry) par sélection clonale. Ph-D thesis, Fac Univ Sc Agr, Gembloux
Jacques D, Marchal M, Curnel Y (2004) Relative efficiency of alternative methods to evaluate wood stiffness in the frame of hybrid larch (Larix x eurolepis Henry) clonal selection. Ann For Sci 61:35–43
Johnson GR, Gartner B (2006) Genetic variation in basic density and modulus of elasticity of coastal Douglas-fir. Tree Genet Genom 3:25–33
Johnson GR, Grotta AT, Gartner BL, Downes G (2005) Impact of the foliar pathogen Swiss needle cast on wood quality of Douglas-fir. Can J For Res 35:331–339
Koizumi A, Ueda K (1986) Estimation of the mechanical properties of standing trees by bending tests. I. Test method to measure the stiffness of a tree trunk. Mokuzai Gakkaishi 32(9):669–676
Koizumi A, Ueda K (1987) Estimation of mechanical properties of standing trees by bending tests III. Modulus of elasticity of tree trunks of plantation-grown conifers. Mokuzai Gakkaishi 33(6):450–456
Koizumi A, Takada K, Ueda K (1990a) Radial growth and wood quality of plus-trees of Japanese larch II. Diameters at breast heights and trunk moduli of elasticity of 18-year-old offspring families. Mokuzai Gakkaishi 36(9):704–708
Koizumi A, Takada K, Ueda K, Katayose T (1990b) Radial growth and wood quality of plus-trees of Japanese larch I. Radial growth, density, and trunk modulus of elasticity of grafted clones. Mokuzai Gakkaishi 36(2):98–102
Kumar S (2004) Genetic parameter estimates for wood stiffness, strength, internal checking, and resin bleeding for radiata pine. Can J For Res 34:2601–2610
Langner W, Reck S (1966) Vergleichende Untersuchungen über das Holz von Larix decidua Mill. Larix leptolepis Gord. Und deren Hybriden. Holzforsch. Berlin 20:192–199
Langner W, Schneck V (1998) Ein Beitrag zur Züchtung von Hybridlärchen (Larix x eurolepis Henry). Sauerländer’s Verlag, Frankfurt am Main
Launay J, Rozenberg P, Pâques LE, Dewitte J-M (2000) A new experimental device for rapid measurement of the trunk equivalent modulus of elasticity on standing trees. Ann For Sci 57:351–359
Leban J-M, Haines DW (1999) The modulus of elasticity of hybrid larch predicted by density, rings per centimeter, and age. Wood Fiber Sci 31:394–402
Mamdy C, Rozenberg P, Franc A, Launay J, Schermann N, Bastien JC (1999) Genetic control of stiffness of standing Douglas fir; from the standing stem to the standardised wood sample, relationships between modulus of elasticity and wood density parameters. Part I. Ann For Sc 56:133–143
Matheson AC, Gapare WJ, Ilic J, Wu HX (2008) Inheritance and genetic gain in wood stiffness in Radiata pine assessed acoustically in young standing trees. Silvae Genetica 57:56–64
Miyajima H, Hasegawa K (1978) Wood quality of plantation-grown hybrid larch trees. Research Bulletins, College Experiment Forests, Hokkaido 35:139–156
Nakada R (2002) Clonal variations of wood properties in Larix kaempferi. In: Proc Int IUFRO Symposium LARIX2002, Gap September 16–21, 2002. Ed Pâques LE
Nanson A, Sacré E (1978) A propos de l’hétérosis de Larix x eurolepis, en particulier pour les propriétés du bois. Bull Rech Agron Gembloux 13:323–336
Nepveu G, Blachon JL (1989) Largeur de cerne et aptitude à l’usage en structure de quelques conifères: douglas, pin sylvestre, pin maritime, épicéa de Sitka, épicéa commun, sapin pectiné. Rev For Fr XLI-6:497–506
Pâques LE (2000) Interspecific hybridisation in larch: the long way to get outstanding varieties. In: QFRI/CRC SPF Symposium ‘Hybrid Breeding and Genetics of Forest trees’, April 9–14, 2000. Noosa, Queensland, Australia
Pâques LE (2004) Role of European and Japanese larch in the genetic control of growth, architecture and wood quality traits in interspecific hybrids (Larix x eurolepis Henry). Ann For Sci 60:1–9
Pâques LE, Rozenberg P (1995) Intraspecific variability of European larch for wood properties: preliminary results. Swedish University of Agricultural Sciences, Dpt of Silviculture, Reports n°39
Pâques LE, Rozenberg P (2009) Ranking larch genotypes with the Rigidimeter: relationships between modulus of elasticity of standing trees and of sawn timber. Ann For Sci 66:414 (1–7)
Reck S (1977) Ergebnisse einer Versuchsanlage mit europäischen Lärchen und Hybridlärchen. Silvae Genet 26:95–101
Reck S (1980) Untersuchung über das Holz der Hybridlärche. Allg Forst-u J Ztg 151:117–120
Ringard E (1980) Mélèze. Queyras, Embrunnais. Mémoire ENITEF, ONF Embrun, INRA Nancy, septembre 1980
Stener L-G, Stenlid J, Karlsson B (2002) Genetic evaluation of growth, external stem quality, wood density and sensitivity to root and butt rot in a Larix x eurolepis trial in South Sweden. In: Proc Int IUFRO Symposium LARIX2002, Gap September 16–21, 2002. Ed Pâques LE
Thill A, Palm R (1984) Etude dendrométrique des mélèzes. Notes techniques du centre d’écologie forestière et rurale, Faculté des Sciences agronomiques de Gembloux
Acknowledgments
This project was partly supported by the EU FAIR project grant CT98-3354 ‘Towards a European Larch Wood Chain’. We are grateful to the technical teams from Orléans and Nancy experimental units for their help with the Rigidimeter measurements. We also thank the anonymous reviewers for their constructive comments.
Author information
Authors and Affiliations
Corresponding author
Additional information
Communicated by S. Aitken
Rights and permissions
About this article
Cite this article
Pâques, L.E., Millier, F. & Rozenberg, P. Selection perspectives for genetic improvement of wood stiffness in hybrid larch (Larix x eurolepis Henry). Tree Genetics & Genomes 6, 83–92 (2010). https://doi.org/10.1007/s11295-009-0230-0
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11295-009-0230-0