
Abstract
The purpose of this study was to examine the relationship between species composition of tropical seagrasses and various physical environmental factors: depth, sediment thickness and silt–clay content in the sediments. We investigated species composition and abundance of seagrasses as well as the physical environmental factors for six transects around Ishigaki Island, southwest Japan. Eight species occurred in the quadrat census. The occurrence frequencies ranged from 66.8% (Thalassia hemprichii) to 4.5% (Enhalus acoroides). Both canonical correspondence analysis (CCA) and cluster analysis elucidated that depth was mainly responsible for the distributions of species and assemblage type. Monte Carlo permutation for partial CCA revealed that 37.5% of the variance was explained by depth, 10.3% by sediment thickness and 4.6% by silt–clay content in the sediment. Twenty-six sites were categorized into four assemblage types by a cluster analysis using the leaf area index (LAI; the ratio of total leaf area to bottom area) as a measure of species abundance. Type I was dominated by T. hemprichii and Cymodocea rotundata, Type II by C. serrulata, Type III by E. acoroides, and Type IV by Halodule pinifolia and Halophila ovalis. Type I occurred mostly in the intertidal zone (91.3±30.5 cm below MSL, mean sea level), Type II in the subtidal zone (179.1±75.0 cm below MSL) and Type IV in both shallow sites (between 47.8 and 75.6 cm below MSL) and in those with low silt–clay contents (between 2.0 and 3.8%).
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Seagrass meadows play essential roles in coastal ecosystems by offering substrata, trophic support, refuge from predation and/or nursery areas for many types of organisms, such as algal epiphytes, invertebrates, fishes, and endangered species such as green turtles and dugongs (Williams and Heck 2001). In the tropical and subtropical Indo-Pacific, seagrass meadows are characterized by high species diversity with mixed vegetation (Tsuda and Kamura 1990; Mukai 1993; Lewmanomont et al. 1996; Terrados et al. 1998; Hemminga and Duarte 2000). Morphology, biomass and growth rate vary among the seagrass species (Vermaat et al. 1995), and their associated faunal communities also change depending on the seagrass species (De Troch et al. 2001; Somerfield et al. 2002). Investigations of relationships between seagrass species composition and factors affecting their variation are essential when evaluating the functions of mixed seagrass meadows.
Several abiotic factors are thought to influence seagrass distribution. During low tide, emergence determines the upper limit of distribution in intertidal areas (Yabe et al. 1995, 1996; Leuschner et al. 1998; Seddon and Cheshire 2001; Tanaka and Nakaoka 2004). Conversely, the deeper limit is attributed to light attenuation by the water column (Dennison 1987; Duarte 1991; Lee Long et al. 1993; Yabe et al. 1996). Siltation is another influential abiotic factor causing both light attenuation and physical burial (Duarte et al. 1997; Bach et al. 1998; Terrados et al. 1998). Sediment depth also affects seagrass distribution, especially for larger species that extend roots and rhizomes deeper into sediments (Duarte et al. 1998).
Several studies have investigated the relationship between these abiotic factors and the distribution of tropical seagrass beds in the Indo-Pacific. In most studies, however, the performance of each species was compared individually with a single environmental factor axis, such as depth of distribution (Mukai et al. 1987; Björk et al. 1999; De Troch et al. 2001) or effect of siltation (Duarte et al. 1997; Terrados et al. 1998). Multiple environmental factor axes affect the distribution of seagrass species simultaneously (Vermaat et al. 1997; Bach et al. 1998), and the resultant distributions may not agree with those predicted by a single factor axis. Seagrass species probably interact with each other in mixed vegetation, which may lead to specific patterns of species composition. Depth and sediment condition are also pointed out as factors that may affect seagrass distribution in subtropical Japan (Kanamoto and Watanabe 1981; Kanamoto 2001). However, these studies did not quantify how much each environmental factor affects the seagrass species distribution. In such cases, multivariate analyses are useful for categorizing community structures and analyzing them in relation to environmental factors (Legendre and Legendre 1998). To our knowledge, only two studies have applied multivariate techniques to tropical seagrass meadows (Erftemeijer and Herman 1994; Bandeira 2002). The former considered only two species and the latter did not analyze environmental factors.
The purpose of this study was to examine the species composition of seagrasses in Ishigaki Island, a subtropical island in southwest Japan. Ishigaki Island is located close to the northern end of the Indo-West Pacific seagrass community, and the number of distributed species is relatively small: nine species (Nozawa 1972; Kanamoto and Watanabe 1981; Toma 1999). Therefore, variations in species composition can be clearly detected. The small size of the island enables us to conduct a whole island survey by setting several research transects around the island.
In the present study, we first measured the occurrence and abundance of seagrass species and multiple physical environmental factors (depth, sediment thickness and silt–clay content in the sediments) on six transects located around Ishigaki Island, southwest Japan. We then analyzed the relationship between species composition and physical environmental factors using both canonical correspondence analysis (CCA) and cluster analysis.
Methods
Study sites
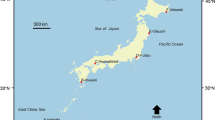
This study was conducted on Ishigaki Island, in the Ryukyu Islands, southwest Japan (24°20′ N, 124°9′ E, Fig. 1). The seagrass flora of the Ryukyu Islands are similar to those of the Philippines, Indonesia, and Papua New Guinea, which have the highest seagrass species richness in tropical seagrass beds worldwide. Among the Ryukyu Islands, Ishigaki Island has the highest seagrass species richness: a total of nine species (Nozawa 1972; Kanamoto and Watanabe 1981; Toma 1999). The total area of the seagrass beds around Ishigaki Island is estimated to be approximately 11 km2 (Okaji and Yamaguchi 1994).

Position of Ishigaki Island, southwest Japan, showing the seagrass beds and location names. E1 Shiraho, E2 Karadake, N1 Kuura, N2 Fukido, W Nagura, S Miyara
The tidal pattern at Ishigaki Island is a mixed dominant semidiurnal type. In 1998, the highest tide was 103 cm above mean sea level (MSL) during a day in summer, while the lowest (extreme low water of spring tide, ELWS) was 123 cm below MSL on a winter night. The mean low water (MLW) was 49 cm below MSL. The annual precipitation was 2,065 mm, the average air temperature was 24.0 °C (29.3 °C in July and 18.3 °C in January), and the average water temperature was 25.0 °C (29.1 °C in July and 20.5 °C in January). The water temperature at Ishigaki Island is relatively warm for its latitude due to the warm Kuroshio Current. The western coast of Ishigaki Island has a poorly developed coral reef crest, and is characterized by bays and inlets. By contrast, the eastern coast is characterized by a well-developed fringing reef with distinct topographical zonation: moat and reef crest (seaward rise of the coral reef flat). Although Ishigaki Island has a land area of 221 km2 and the rivers on the island are relatively small, some of the rivers yield considerable amounts of silt–clay, especially the Miyara and Todoroki Rivers (Fig. 1) (Okinawa Prefectural Central Council of Fisheries & Okinawa Prefectural Fisheries Promotion Fund 1989).
Seagrass beds were identified from color aerial photos of approximately 1/12,500 scale taken by Okinawa Prefecture in 1994 (Fig. 1). Based on the aerial photos and preliminary field surveys, we chose six sites with different water depth profiles and siltation loads, E1 (Shiraho), E2 (Karadake), N1 (Kuura), N2 (Fukido), W (Nagura), and S (Miyara) (Fig. 1). E1, E2 and S were situated inside the coral reef crest, while the others were not. E2, N2, W and S were located close to the river mouth, others did not have apparent river effects. At each site, we established one line transect perpendicular to the shoreline. Transects were set from the benchmarks located on the supralittoral zone to the edge of the seagrass distribution. The transects were 400, 600, 500, 500, 900, and 600 m long for E1, E2, N1, N2, W and S, respectively. We placed a “research point” every 10 m along the transects from the benchmarks. Three successive research points were grouped together and were named “sites” for this study. The centers of the three successive points were placed at 100-m intervals from the benchmarks.
Field survey
A field survey was conducted in September 1998. Water depth was measured at each research point. The measured depths were converted to a depth from the MSL using the tide table (Japan Meteorological Agency 1997). Sediment thickness was measured at every research point using a 90-cm-long peg, which was hammered into the bottom until it hit hard substratum. This was repeated three times at each research point and averaged. When the peg did not reach hard substratum, the value was considered to exceed 90 cm. At every 100-m research point along each transect, sediment samples were collected using a 25-cm-long plastic corer with an internal diameter of 5 cm. In cases where a large amount of organic matter, such as seagrass leaves, was contained in the samples, we removed them before drying. After drying at 105 °C for more than 24 h, the silt–clay was carefully washed away with running water through a 63-μm-mesh sieve. The samples were dried again under the same conditions and then weighed. The silt–clay content (%) in the sediment was calculated as follows: the difference in the weights before and after sieving (silt–clay) with respect to the whole sediment containing the silt–clay, fine sand and coarse sand.
At every research point, the number of shoots of each seagrass species was counted using a 100-cm2 quadrat. Three quadrats were placed at each research point randomly and the values were averaged. We used a 625-cm2 quadrat for the large seagrass, Enhalus acoroides.
The abundance of each species was represented by the ratio of total leaf area to bottom area (LAI), calculated at each research point. To measure the leaf area per shoot, about 30 shoots of each species were collected randomly. The lengths and widths of all leaves were measured, the number of leaves on a shoot was counted, and the average leaf area (single-sided) was calculated for each shoot. The leaf area of a shoot multiplied by the shoot density per area equaled LAI. For Syringodium isoetifolium, which has cylindrical leaves, the leaf area was approximated using the following equation:
Statistical analysis
CCA was employed to summarize the distributions of seagrass species and their relationships to environmental variables. Twenty-five sites [at W-800 m we did not obtain silt and clay (%) data, so the site was omitted] and eight species of LAIs are displayed as points on the biplots, while environmental variables are indicated by arrows. Before ordination analysis, our data was transformed into log(x+1). To isolate the effects of each environmental factor, partial CCA with covariance was performed. This analysis was also applied in order to assess the effects of two of the factors on species composition, adjusted for the effect of other factors. In canonical analysis, eigenvalues can be tested for significance using the method of permutations (Legendre and Legendre 1998). The test statistics from Canoco 4.5 for Windows (Ter Braak and Smilauer 2002) give partial F ratios for the relationships between the environmental factors and the tropical seagrass species structure, and then the significance of each F ratio can be tested using Monte Carlo permutation for partial CCA, with 999 permutations employed at the 5% significance level (ter Braak and Smilauer 2002; Leps and Smilauer 2003).
Similarities in the assembly types of seagrasses among the 26 sites with vegetation were analyzed by cluster analysis using the Morisita–Horn similarity index C λ (Horn 1966), with a classification strategy that involved recalculating C λ at each step. The seagrass species composition and abundance at each site were represented by species richness (the number of species), total LAI, and the LAI of each species.
Results
Environmental factors
The depths of the research points ranged from 1 to 323 cm below MSL. The average depth of the transect was shallowest in transect S (54.4 cm) and deepest in E2 (164.7 cm) (Table 1, Fig. 2). The silt–clay content in the sediments across all sites ranged from 0.9 to 19.6%. The average silt–clay content in the sediments was relatively low in E1 (2.3%), N2 (3.6%) and W (4.1%), and high in N1 (10.0%) and S (10.3%). The sediment layer was thick in N1 (mean±SD, 67.6±20.2 cm) and W (78.8±23.9 cm), and thin in E1 (17.0±17.8 cm) and S (22.7±23.5 cm). The topographic profiles were uneven in transects E1 and E2, and smooth in N1, N2 and W (Fig. 2). In the uneven transects, the distribution of sand and seagrasses was intermittent. By contrast, the distribution of sand and seagrass was continuous in the smooth transect. In transect S, the topographic profile was smooth, although the distribution of sand and seagrass was intermittent.

Silt–clay content, species shoot density and topographical profiles of the six transects. The pie charts show the silt–clay (<63-μm-sized sediment) content in the sediments. The histograms indicate shoot density; each bar shows the average shoot numbers from three quadrats at every research point. Th Thalassia hemprichii, Cr Cymodocea rotundata, Cs C. serrulata, Si Syringodium isoetifolium, Hu Halodule uninervis, Ho Halophila ovalis, Hp Halodule pinifolia, Ea Enhalus acoroides. In the topographical profiles, the upper bold line indicates the depth from MSL, and the lower fine line shows the distance from MSL to hard substratum. The distance involves sediment thickness and water depth. A dotted line means that the sediments are over 90 cm thick. Abbreviations of location names are listed in Fig. 1
Species distribution
Nine species were observed around Ishigaki Island. The frequency of occurrence varied among species. Of 204 vegetated research points, Thalassia hemprichii was recorded at 136 research points (66.7%), followed by Cymodocea serrulata (45.1%), Cymodocea rotundata (37.3%), Halodule uninervis (25.0%), Halophila ovalis (15.7%), Halodule pinifolia (15.2%), S. isoetifolium (11.3%) and E. acoroides (4.4%). Small patches of Zostera japonica were detected around the shallower edge of the seagrass beds in N2, and only three shoots of E. acoroides was found around S-600, although they did not occur in any of the quadrats. Some seagrass beds were so mixed that six species coexisted at one research point (triplicate of 100 cm2). Five to seven species (mean±SD, 6.0±0.9) were recorded along each transect (Table 2).
Shoot size and density varied among species (Table 3, Fig. 2). E. acoroides had the largest shoot leaf area, T. hemprichii, C. rotundata, and C. serrulata were middle-sized, and the other four species were small. The density of H. pinifolia exceeded 100 shoots/100 cm2 at some research points. Halophila ovalis and Halodule uninervis were relatively dense, while E. acoroides was sparse, less than one shoot/100 cm2. The densities of the other four species, T. hemprichii, C. rotundata, C. serrulata and S. isoetifolium, were around five to ten shoots /100 cm2. Based on the LAI, T. hemprichii, C. rotundata and C. serrulata were abundant (Table 2). Except for N2, where the largest species (E. acoroides) grew, the relative abundances of these three species exceeded 90% of the total leaf area in each transect.
Relationship between each environmental factor and the species distribution
The distributions of the seagrass species overlapped broadly when plotted against each environmental factor (Fig. 3). Along the depth gradient, H. pinifolia occurred in the shallowest parts of the beds, followed by H. ovalis, C. rotundata, T. hemprichii, S. isoetifolium, H. uninervis, C. serrulata, and E. acoroides, in that order. The entire range of H. pinifolia was above ELWS, whereas almost all of the range of E. acoroides was below ELWS. Along the gradient of silt–clay content, H. ovalis was distributed at sites with the lowest silt–clay content, followed by H. pinifolia, E. acoroides, C. serrulata, S. isoetifolium, H. uninervis, T. hemprichii, and C. rotundata. For sediment thickness, E. acoroides occurred at points with the thinnest sediment layer, followed by H. ovalis, T. hemprichii, H. uninervis, H. pinifolia, C. serrulata, S. isoetifolium, and C. rotundata. H. ovalis and T. hemprichii even occurred at sites without a sediment layer.

Depth, silt–clay content, and sediment thickness for each species. Dots indicate the mean. Boxes show the standard deviations. Bars indicate ranges. Abbreviations of species names are listed in Fig. 2. The numbers in parentheses are the sample sizes. ELWS is indicated on the depth graph
Canonical correspondence analysis
Depth was the most explainable environmental factor in this study. Monte Carlo permutation revealed that 37.5% of the variance was explained by depth (Table 4). Generally in CCA diagrams, objects on the edge of the diagram provide the largest contribution to overall variance, and those near the origin are of minor importance (ter Braak and Smilauer 2002). N2-500 and E2-600 (Fig. 4a) and LAI of C. serrulata and E. acoroides (Fig. 4b) had positive correlation with depth. On the other hand, LAI of H. pinifolia had negative correlation.

CCA biplot of analysis with three environmental variables: depth, sediment thickness and silt–clay content of the sediment. Scaling type II was employed. a Environmental variables versus sites. b Environmental variables versus species. Site names (e.g., E1-100) indicate location names and distances from the benchmarks (m). Abbreviations of location names are listed in Fig. 1. LAI leaf area index. Abbreviations of species names are listed in Fig. 2
10.3% of the variance was explained by sediment thickness. Sites on transect W (e.g., W-800) gave roughly the same directions with the sediment arrow (Fig. 4a). LAI of H. pinifolia was the closest to the edge of the arrow (Fig. 4b). 4.6% of the variance was explained by the silt–clay content in the sediments.
Assemblage type categorization by cluster analysis
The cluster analysis categorized the 26 sites into four distinct assemblage types (Fig. 5). Thirteen sites were classified as Type I, nine sites as Type II, one site as Type III, and three sites as Type IV. Actually, Type IV contained two categories which branched off at around Cλ=0.5. However, it only contained three sites and was branched at the first junction from the others. Therefore, we united them into one type, IV. In Type I, we identified two subgroups (I-a and I-b). T. hemprichii was dominant at the sites in I-a (six sites) and they occupied more than 60% of the total LAI. T. hemprichii and C. rotundata, combined, were dominant at the sites in I-b (seven sites) and they occupied more than 79% of the total LAI. Similarly in Type II, we identified two subgroups (II-a and II-b). In both subgroups, C. serrulata was dominant. In II-a (four sites), the share of C. serrulata ranged from 29 to 46%, whereas that in II-b (five sites) ranged from 66 to 100%. Type III consisted only of one site (N2-400 m), where E. acoroides occurred. In Type IV, only H. pinifolia and H. ovalis were observed.

Categorization of the sites based on species richness, total leaf area index (LAI), and the LAI of each species from the cluster analysis. Site names (e.g., E1-100) are explained in Fig. 4
The four assemblage types categorized by the cluster analysis clearly separated when the sites were plotted against depth and silt–clay content (Fig. 6a). Types I (between 35.6 and 153.1 cm below MSL) and IV (between 47.8 and 75.6 cm) occurred in shallower water, whereas Type II was distributed in deeper water (between 91.3 and 305.8 cm). Upon comparing the silt–clay content in the sediments, Type IV was confined to sites with lower silt–clay contents (between 2.0 and 3.8%), while Types I (between 1.2 and 16.9%) and II (between 3.5 and 16.0%) occurred in a broad range of silt–clay contents. The distribution of assemblage types was not clear when the sites were plotted against sediment thickness, except to say that only Type I-a occurred at sites with a sediment thickness less than 20 cm (Fig. 6b).

Relationship between assemblage types and environmental factors: a depth and silt–clay content, b depth and sediment thickness. Abbreviations of location names are listed in Fig. 1
Discussion
Nine species of seagrasses were found around Ishigaki Island in this study, agreeing with the previous reports on the seagrass distribution for the whole Ishigaki Island (Nozawa 1972; Kanamoto and Watanabe 1981; Tsuda and Kamura 1990; Toma 1999). The dominance of T. hemprichii in the Yaeyama Islands (including Ishigaki Island) is similar to that observed in previous studies (Takada and Abe 2002). The number of species in Ishigaki Island was small compared to the numbers of species seen in studies of whole seagrass flora within countries in the Indo-Pacific tropical area: 14 species in the Philippines and India, 13 species in Papua New Guinea, and 12 species in Thailand (Brouns 1987; Mukai 1993; Jagtap 1996; Lewmanomont et al. 1996). The dominant species in those areas are the same as those on Ishigaki Island when they are compared at a similar spatial scale as to the present study—between 150 and 500 m of transects or points within several km2 (Nienhuis et al. 1989; Tomasko et al. 1993; Vermaat et al. 1995; Bach et al. 1998)—except for E. acoroides, which only occurred at one site in this study. Ishigaki Island is the northern limit of E. acoroides (Nozawa 1972; Kanamoto and Watanabe 1981; Toma 1999), and this may be related to its limited occurrence around the island.
Each assemblage type was not confined to a specific transect. Type I appeared in all six transects, Type II in four, and Type IV in two transects. These occurrences of assemblage types suggest that an environmental gradient affecting assemblage types occurs within each transect.
In the present study, CCA quantified the contribution of each physical environmental factor as a new result in comparison with the conclusions of previous studies (Kanamoto and Watanabe 1981; Kanamoto 2001). Among the various possible physical environmental factors, depth made the largest contribution to species distribution (37.5%). On the other hand, CCA did not detect the simultaneous effects of multiple physical environmental factors on seagrass species composition (Table 4).
Cluster analysis also elucidated that depth was mainly responsible for assemblage type distributions. Simultaneous effects from multiple physical environmental factors were only detected for Type IV, which occurred in both shallow and low-silt–clay sites. The ELWS of Ishigaki Island is about 120 cm below MSL. Therefore, Types IV (mean depth 64.3 cm below MSL) and I (91.3 cm) were dominant in the intertidal zone, whereas Type II (179.1 cm) was subtidal. Tanaka and Nakaoka (2004) revealed that C. serrulata, which was dominant in Type II, was sparse under intertidal conditions because its long vertical rhizome exposes it to severe desiccation. T. hemprichii and C. rotundata, which were dominant in Type I, were larger than H. pinifolia and H. ovalis, which were dominant in Type IV (Table 3). Small flexible species might survive emergence by lying flat on moist sediment, as Björk et al. (1999) pointed out. On the other hand, small species are likely to lose out in the competition for light. Since the vertical distribution range was narrow (about 30 cm), Type IV was noticeable in sites where the slope was gentle and the sediment was continuous, such as close to a river mouth.
Previous studies describing the depth distribution of each seagrass can be explained using our assemblage type classification. Brouns and Heijs (1991) found that T. hemprichii and C. rotundata were the dominant seagrass species at their study sites in Indonesia and Papua New Guinea. This combination occurred in the intertidal zone (between ELWS+0.5 and +0.2 m) on a variety of substrata. This relationship corresponds to our Type I assemblage. C. serrulata was abundant at most of the stations in the survey at Negros Oriental, in the Philippines (Tomasko et al. 1993). All of the stations in the study were situated deeper than 67 cm below MLW, equivalent to our Type II assemblage. At Cape Bolinao, in the Philippines, T. hemprichii was most abundant around MLT, C. rotundata at 0.5 m below MLT, and C. serrulata at 1.0 m below MLT (Bach et al. 1998). These distributions correspond to our Types I and II. Therefore, Types I and II of our community classification are not specific to Ishigaki Island, but occur widely in the Indo-Pacific tropical area. We could not find appropriate examples to show the relationship between depth and assemblage type for Types III and IV. This will be elucidated in future studies.
Sediment thickness appeared to be less related to the distribution of species and assemblage type distribution. The roots and rhizomes of each species, except for E. acoroides, were distributed at sites with a sediment layer thinner than 40 cm (Duarte et al. 1998). In this study, sediment depth exceeded 40 cm at 74.4% of the research points (calculated from raw data). Sediment depth was not a major factor in determining the seagrass distribution at Ishigaki Island. Type I-a communities were mostly found at sites with thin sediment layers (less than 20 cm). These sites were situated in transects with intermittent sediment distributions (E1, E2, and S), where the hard substrata was frequently exposed. In Type I-a, only T. hemprichii was dominant. The internode length on horizontal rhizomes of T. hemprichii was significantly shorter than in other species (Vermaat et al. 1995), allowing this species to grow flexibly to fit the interstices in the substrata or dead coral. This morphological feature of T. hemprichii was probably related to the occurrence of Type I-a at sites with the lowest sediment thickness.
Silt–clay content in the sediment did not show strong relationships to distribution of species and assemblage type in this study, although Type IV assemblage appeared at sites with low silt–clay content. Small H. pinifolia and H. ovalis may be easily buried when the silt–clay content in the sediment is high. The maximum silt–clay content in this study was 17%, and most of sites were below 12%. Terrados et al. (1998) reported sites with more than 40% silt–clay content, along which changes in species distribution were discovered. The weak effect of silt–clay content in this study may be due to the limited range on this island.
In this study, we quantified the contributions of three physical environmental factors and concluded that depth is the factor most responsible for seagrass species composition. Around half (49.6%) of the variance was explained by three factors: depth, sediment thickness and silt–clay content in the sediments. The rest of the variance may be caused by several possible factors, such as nutrient content in the sediment, salinity, turbidity, grazing pressure, epiphyte and interspecific competition of seagrasses.
References
Bach SS, Borum J, Fortes MD, Duarte CM (1998) Species composition and plant performance of mixed seagrass beds along a siltation gradient at Cape Bolinao, The Philippines. Mar Ecol Prog Ser 174:247–256
Bandeira SO (2002) Diversity and distribution of seagrasses around Inhaca Island, southern Mozambique. S Afr J Bot 68:191–198
Björk M, Uku J, Weil A, Beer S (1999) Photosynthetic tolerances to desiccation of tropical intertidal seagrasses. Mar Ecol Prog Ser 191:121–126
Brouns JJWM (1987) Quantitative and dynamic aspects of a mixed seagrass meadow in Papua New Guinea. Aquat Bot 29:33–47
Brouns JJWM, Heijs FML (1991) Seagrass ecosystem in the tropical west Pacific. In: Mathieson AC, Nienhuis PH (Eds) Ecosystems of the world 24, intertidal and littoral ecosystems. Elsevier, Amsterdam, pp 371–390
Dennison WC (1987) Effects of light on seagrass photosynthesis, growth and depth distribution. Aquat Bot 27:15–26
De Troch M, Gurdebeke S, Fiers F, Vincx M (2001) Zonation and structuring factors of meiofauna communities in a tropical seagrass bed (Gazi Bay, Kenya). J Sea Res 45:45–61
Duarte CM (1991) Seagrass depth limits. Aquat Bot 40:363–377
Duarte CM, Terrados J, Agawin NSR, Fortes MD, Bach S, Kenworthy WJ (1997) Response of a mixed Philippine seagrass meadow to experimental burial. Mar Ecol Prog Ser 147:285–294
Duarte CM, Merino M, Agawin NSR, Uri J, Fortes MD, Gallegos ME, Marba N, Hemminga MA (1998) Root production and belowground seagrass biomass. Mar Ecol Prog Ser 171:97–108
Erftemeijer PLA, Herman PMJ (1994) Seasonal changes in environmental variables, biomass, production and nutrient contents in two contrasting tropical intertidal seagrass beds in South Sulawesi, Indonesia. Oecologia 99:45–59
Hemminga MA, Duarte CM (2000) Diversity of seagrass meadow. In: Hemminga MA, Duarte CM (eds) Seagrass ecology. Cambridge University Press, Cambridge, pp 9–14
Horn HS (1966) Measurement of “overlap” in comparative ecological studies. Am Nat 100:419–424
Jagtap TG (1996) Some quantitative aspects of structural components of seagrass meadows from the southeast coast of India. Bot Mar 39:39–45
Japan Meteorological Agency (1997) Tide tables for the year 1998 (in Japanese). Japan Meteorological Agency, Tokyo
Kanamoto Z (2001) Spatial distributions of seagrass and their seasonal change in Nagura Bay, Ishigaki Island, Okinawa. Otsuchi Mar Sci 26:28–39
Kanamoto Z, Watanabe T (1981) Ecological study of the seagrass meadows in Nagura Bay Ishigaki Island Okinawa 1. Ecological distribution of seagrass. Benthos Res 21/22:1–14
Lee Long WJ, Mellors JE, Coles RG (1993) Seagrasses between Cape York and Hervey Bay, Queensland, Australia. Aust J Mar Freshw Res 44:19–31
Legendre P, Legendre L (1998) Numerical ecology, 2nd Engl edn. Elsevier, Amsterdam
Leps J, Smilauer P (2003) Multivariate analysis of ecological data using CANOCO. Cambridge University Press, Cambridge
Leuschner C, Landwehr S, Mehling U (1998) Limitation of carbon assimilation of intertidal Zostera noltii and Z. marina by desiccation at low tide. Aquat Bot 62:171–176
Lewmanomont K, Deetae S, Srimanobhas V (1996) Seagrasses of Thailand. In: Kuo J, Phillips RC, Walker DI, Kirkman H (eds) Seagrass biology: Proceedings of an international workshop, Rottnest Island, Western Australia, 25–29 January 1996. Faculty of Science, The University of Western Australia, Nedlands, Australia, pp 21–26
Mukai H (1993) Biogeography of the tropical seagrasses in the Western Pacific. Aust J Mar Freshw Res 44:1–17
Mukai H, Nojima S, Nishihira M (1987) Seagrass coverage and distribution in Loloata seagrass bed. In: Hattori A (eds) Studies on dynamics of the biological community in tropical seagrass ecosystems in Papua New Guinea: the second report. Ocean Research Institute, University of Tokyo, Tokyo, pp 18–27
Nienhuis PH, Coosen J, Kiswara W (1989) Community structure and biomass distribution of seagrasses and macrofauna in the Flores Sea, Indonesia. Neth J Sea Res 23:197–214
Nozawa Y (1972) On the sea-grass from Ishigaki Island. Mem Kagoshima Junshin Jr Coll 2:56–66
Okaji K, Yamaguchi M (1994) Okinawa prefecture. In: The report of the Marine Biotic Environment Survey in the 4th National Survey on the Natural Environment, Vol. 2: Algal and sea-grass beds, March 1994. Nature Conservation Bureau, Environment Agency and Marine Parks Center of Japan, Tokyo, pp. 349–359 (in Japanese)
Okinawa Prefectural Central Council of Fisheries & Okinawa Prefectural Fisheries Promotion Fund (1989) The results of a survey on the water pollution by reddish soil. In: The water pollution by reddish soil in coastal area of Okinawa, current condition of reddish soil and preventive measures. Okinawa Prefectural Central Council of Fisheries & Okinawa Prefectural Fisheries Promotion Fund, Okinawa, Japan, pp 39–43 (in Japanese)
Seddon S, Cheshire AC (2001) Photosynthetic response of Amphibolis antarctica and Posidonia australis to temperature and desiccation using chlorophyll fluorescence. Mar Ecol Prog Ser 220:119–130
Somerfield PJ, Yodnarasri S, Aryuthaka C (2002) Relationship between seagrass biodiversity and infaunal communities: implications for studies of biodiversity effects. Mar Ecol Prog Ser 237:97–109
Takada Y, Abe O (2002) Local variations in shoot density, shoot morphology, and bite mark frequency of subtropical seagrasses in Japan. Bull Fish Res Agency Jpn 4:1–10
Tanaka Y, Nakaoka M (2004) Emergence stress and morphological constraints affect the species distribution and growth of subtropical intertidal seagrasses. Mar Ecol Prog Ser 284:117–131
Ter Braak CJF, Smilauer P (2002) Canoco reference manual and CanoDraw for Windows user’s guide: Software for Canonical Community Ordination (version 4.5). Microcomputer Power, Ithaca, NY
Terrados J, Duarte CM, Fortes MD, Borum J, Agawin NSR, Bach S, Thampanya U, Kamp-Nielsen L, Kenworthy WJ, Geertz-Hansen O, Vermaat J (1998) Changes in community structure and biomass of seagrass communities along gradients of siltation in SE Asia. Est Coast Shelf Sci 46:757–768
Toma T (1999) Seagrasses from the Ryukyu Islands—I, species and distribution. Biol Mag Okinawa 37:75–92 (in Japanese with English abstract)
Tomasko DA, Dawes CJ, Fortes MD, Largo DB, Alava MNR (1993) Observations on a multi-species seagrass meadow offshore of Negros Oriental, Republic of the Philippines. Bot Mar 36:303–311
Tsuda RT, Kamura S (1990) Comparative review on the floristics, phytogeography, seasonal aspects and assemblage patterns of the seagrass flora in Micronesia and the Rhykyu Islands. Galaxea 9:77–93
Vermaat JE, Agawin NSR, Duarte CM, Fortes MD, Marba N, Uri JS (1995) Meadow maintenance, growth and productivity of a mixed Philippine seagrass bed. Mar Ecol Prog Ser 124:215–225
Vermaat JE, Agawin NSR, Fortes MD, Uri JS, Duarte CM, Marba N, Enriquez S, Viersen WV (1997) The capacity of seagrass to survive increased turbidity and siltation: the significance of growth form and light use. Ambio 26:499–504
Williams SL, Heck KL Jr (2001) Seagrass community ecology. In: Bartness MD, Gaines SD, Hey ME (eds) Marine community ecology. Sinauer Associates Inc., Sunderland, MA, pp 317–337
Yabe T, Ikushima I, Tsuchiya T (1995) Production and population ecology of Phyllospadix iwatensis Makino. I. Leaf growth and biomass in an intertidal zone. Ecol Res 10:291–299
Yabe T, Ikushima I, Tsuchiya T (1996) Production and population ecology of Phyllospadix iwatensis Makino. II. Comparative studies on leaf characteristics, foliage structure and biomass change in an intertidal and subtidal zone. Ecol Res 11:291–297
Acknowledgments
We would like to thank Dr. M. Nakaoka for his invaluable suggestions. Dr. K. Suetsugu and Dr. M. Shimanaga helped us with statistical analyses. We are also grateful to Dr. K. Aioi, who helped us to identify the seagrass species. Dr. J. Kuo, Dr. H. Yamano, Dr. M. Yamamuro and Dr. I. Koike offered supportive comments, and thanks are due to them. In addition, we would like to thank Dr. Y. Umezawa, Dr. A. Watanabe, and N. Shinohara for their assistance in the field survey. We thank Fuyo Ocean Development Co. Ltd., Marine Ecological Institute Co. Ltd., H. Shimabukuro and M. Taira for help with our field research. We also thank Dr. H. Matsuda, Dr. A. Tamaki and other anonymous reviewers for the constructive comments. This research was supported by CREST, the Japan Science and Technology Corporation.
Author information
Authors and Affiliations
Corresponding author
About this article
Cite this article
Tanaka, Y., Kayanne, H. Relationship of species composition of tropical seagrass meadows to multiple physical environmental factors. Ecol Res 22, 87–96 (2007). https://doi.org/10.1007/s11284-006-0189-3
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11284-006-0189-3