
Abstract
In forest ecosystems, fine roots have a considerable role in carbon cycling. To investigate the seasonal pattern of fine root demography, we observed the fine root production and decomposition processes using a minirhizotron system in a Betula-dominated forest with understory evergreen dwarf bamboo. The length density of fine roots decreased with increasing soil depth. The seasonal patterns of each fine root demographic parameter (length density of visible roots, rates of stand-total fine root production and decomposition) were almost the same at different soil depths. The peak seasons of the fine root demographic parameters were observed in the order: stand-total fine root production rate (late summer) > length density of the visible roots (early autumn) > stand-total fine root decomposition rate (autumn, and a second small peak in spring). The fine root production rate was high in the latter part of the plant growing season. Fine root production peaked in late summer and remained high until the end of the tree defoliation season. The higher stand-total fine root production rate in autumn suggests the effect of understory evergreen bamboo on the stand-total fine root demography. The stand-total fine root decomposition rate was high in late autumn. In the snow-cover period, the rates of both fine root production and decomposition were low. The fine root demographic parameters appeared to show seasonal patterns. The fine root production rate had a clearer seasonality than the fine root decomposition rate. The seasonal pattern of stand-total fine root production rate could be explained by both overstory and understory above-ground productivities.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Below-ground parts of plants, especially roots in the finest diameter class (fine roots), play a considerable role in carbon cycling in forest ecosystems because fine roots have a fast turnover in proportion to their small biomass (Grier et al. 1981; Keyes and Grier 1981; Comeau and Kimmins 1989). It is known that the fine root biomass accounts for only 1–12% of forest tree total biomass, although 8–69% of net primary production (NPP) of trees in the forest is consumed by the production–death–decomposition cycle (PDD cycle) of fine roots (Grier et al. 1981; Vogt et al. 1982; Comeau and Kimmins 1989). Fine roots consume great amounts of NPP in spite of their small biomass simply because of the fast turnover of their PDD cycle. The value of the turnover effect is large because consumption of NPP by fine roots is usually estimated by the multiplication of the fine root biomass and turnover. Therefore, more accurate estimation of the fine root turnover is needed to better understand forest carbon dynamics. To estimate fine root turnover, information about fine root demography, such as production, growth, death and decomposition, is necessary.
The minirhizotron (MR) method is a non-destructive method in which long transparent tubes are placed in the study field, and fine roots on the soil-tube boundary layer are observed with a CCD (charge-coupled device) camera (Smit et al. 2000; Johnson et al. 2001; Satomura et al. 2001). This method generally provides more accurate estimates of fine root demography than do other methods. Using the MR method, fine root demography has been studied for decades (e.g., Eissenstat and Caldwell 1988; Hendrick and Pregitzer 1993a). These studies have shown the seasonal patterns of fine root production and decomposition (Steele et al. 1997; Cheng and Bledsoe 2002; Baddeley and Watson 2004), life span and turnover of fine roots (Tingey et al. 2000; Arnone III et al. 2000) by monitoring the fine root production, death and decomposition processes. The peaks of the fine root production rate have been observed in the growing season, that is, spring (Hendrick and Pregitzer 1993b; Joslin et al. 2001) and summer (Burton et al. 2000). Possible factors controlling the peak seasons of stand-total fine root production rate are fine root damage in winter (Tierney et al. 2001), strong prolonged summer drought (Joslin et al. 2000; Xu and Baldocchi 2003), and the fine root biomass and its dynamics of understory plants (Cheng and Bleadsoe 2002). However, much more data on the seasonal patterns of fine root production are needed to clarify the factors and mechanisms determining the peak season of fine root production rate.
Above-ground photosynthetic activity also possibly affects the fine root production rate (Fitter et al. 1998; Edwards et al. 2004). The seasonal pattern of the stand-total above-ground photosynthetic activity is closely related to the forest structure and plant composition (Lei et al. 1994). The seasonal pattern of the stand-total fine root production rate, therefore, could be associated with the pattern of the above-ground photosynthetic activity. However, forests for which data are available on both stand-total fine root production rate and above-ground photosynthetic activity are valuable. Therefore, in forest ecosystems, it is normally difficult to examine the relationship between these data. We have both data sets in a long-term ecological research site (the Takayama Experimental Forest), which enables us to examine this relationship. The Takayama Experimental Forest is a cool-temperate forest with clear two-layered vegetation structure (overstory trees and understory dwarf bamboo) dominated by Betula species (Betula ermanii and Betula platyphylla).
In cool-temperate regions in the northern hemisphere, there are seasonal deciduous forests, in which active below-ground fine root production would be expected in the growing season (the season showing high above-ground photosynthetic activities). The seasonal patterns of the above-ground productivity and leaf dynamics are different between overstory and understory plants in a seasonal deciduous forest. The amount of leaves and leaf photosynthetic capacity of the overstory deciduous trees, are normally lower in the spring and autumn seasons (the tree leaf emergence and defoliation periods, respectively) and they are higher in the summer season (Uemura 1994; Maeno and Hiura 2000). On the other hand, the amount of leaves and the ability of leaves to photosynthesize in the understory plants are highest in the spring and autumn, that is, the seasons in which the overstory leaf cover is less, resulting in better light conditions for the understory plants (Koizumi and Oshima 1985; Lei et al. 1994). Moreover, the seasonal patterns of the above-ground productivity and leaf dynamics of understory plants depend on their life form (evergreenness or deciduousness). Evergreen plants have a longer active photosynthesis season than do deciduous plants.
Betula-dominated forests are found in cool-temperate regions and have either deciduous understory vegetation composed of many different tree species or evergreen understory vegetation that is dominated by evergreen dwarf bamboo (Tansley 1965; Walter 1979; Miyawaki and Okuda 1990). In the former type of forest, the fine root production rate peaks in late summer and immediately decreases with the beginning of tree leaf defoliation in the autumn (Ovington and Murray 1968). In the latter type of forest such as our study site, the Takayama Experimental Forest, dwarf bamboo produces below-ground parts mainly in the autumn while it produces above-ground parts mainly in the spring (Nishimura et al. 2004). If the stand-total fine root production rate is closely related to phenological traits of the dominant tree species and there is no rubric limit to environmental factors during the growing season, the stand-total fine root production rate in the latter type of forest would also peak in late summer. And if the effect of understory evergreen bamboo on the seasonal pattern of the stand-total fine root production rate is considerable, the stand-total fine root production could be continuously active from summer until autumn in the latter type of forest.
To clarify the relationship between the seasonal patterns of fine root demography and above-ground photosynthetic activity, we studied the fine root demography in relation to soil depth for over a year in a Betula-dominated forest with understory evergreen bamboo (the Takayama Experimental Forest) using the MR method. In this forest, the seasonal patterns of leaf dynamics and photosynthetic activity in both overstory and understory plants are similar to those in other general cool-temperate deciduous forests described previously (Kawamura et al. 2001; Sakai et al. 2002; Nishimura et al. 2004; Muraoka and Koizumi 2005; Sakai and Akiyama 2005). Bamboo root dynamics has been roughly characterized in our forest (Nishimura et al. 2004). However, very little information is available on tree fine root dynamics. Both rates of stand-total fine root production and fine root decomposition were quantified in relation to soil depth in each observation period. Based on the results, we describe how fine root demography changes with season and depth and discuss how it is related to above-ground productivity. We also discuss the effects of a layered vegetation structure and its plant species composition on the seasonal pattern of the fine root production rate. Active fine root production in our study forest is expected in both summer and autumn seasons.
Materials and methods
Study site
The study site is a long-term ecological research site, the Takayama Experimental Forest (Q50) of Gifu University. This site Q50 (1-ha plot) is located at 1,430 m a.s.l. on the north-west slope of Mt. Norikura in Gifu Prefecture (36°8′N, 137°26′E), the central region of the main island of Japan. The 1980–2000 annual means of air temperature and precipitation are 6.1°C and 2,175 mm (data from Takayama Experimental Station, River Basin Research Center, Gifu University). Average air and soil temperatures during the study period (July 2000 to June 2001) were 7.0 and 8.2°C, respectively. Even in mid-winter, daily soil temperature did not fall below freezing (Fig. 1a) since the study site is covered with thick snow (maximum snow depth during normal years is 1–2 m) due to its Sea of Japan coastal climate. During the survey period, water potential was low for a short period in late summer and higher in other seasons (Fig. 1b). The study site is on a sedimentary rock formed from an argillaceous sandy soil of the Quaternary period in the Cenozoic (Igi 1991). The soil type is brown forest soil with an A0 layer (thickness about 3–7 cm) and an A layer (thickness about 20–50 cm) (for details see Jia et al. 2002).

Air temperature (°C), soil temperature (°C) and soil water potential (kPa) during the survey period in a cool-temperate deciduous forest in Japan (36°08′25″N, 137°25′35″E, altitude about 1,400 m). Air temperature data are taken at a height of 1.5 m. Soil data are taken at a depth of 5 cm. Air temperature and soil temperature were recorded by a thermo-data logger (StowAway TidBit; Onset Computer Co., Mass, USA) and soil water potential was checked using a tension meter (DM-8; Takemura, Tokyo, Japan). Soil temperature was never lower than 0°C even in winter due to the thick snow layer. Water potential was low for a short period in the summer season. Bars at each point represent standard errors
The study area is an approximately 50-year-old secondary cool-temperate deciduous forest that has two distinct structural components, that is, overstory and understory. The overstory components are comprised of diverse species of trees (37 species) and shrubs (7 species). The dominant species are Quercus crispula Blume, B. ermanii Cham and B. platyphylla Sukatchev var. japonica Hara. The relative basal areas of these trees are 27.2%, 25.4% and 15.0%, respectively (Kawamura et al. 2001). Other relatively abundant species include Magnolia obovata Thunb, Acer rufinerve Sieb. Et Zucc. and Tilia japonica Simonkai. Total basal area of the trees and tree density in the study area are 32.34 m2 ha−1 and 1,907 ha−1, respectively (Ohtsuka et al. 2005). The understory vegetation is mainly Sasa senanensis Rehd (bamboo coverage more than 80%).
The trees gradually produce leaves in spring, become fully foliated in summer, and defoliate in autumn (Kawamura et al. 2001). The NPP of the canopy tree leaves is highest in late summer (Muraoka and Koizumi 2005). During the non-snow-cover period, the NPP of the bamboo tends to be high when the tree canopy is not closed, that is, in the spring and autumn (Sakai and Akiyama 2005). During the snow-cover period, the above-ground parts of the bamboo are buried in snow. The bamboo vigorously produces new organs (above- and below-ground parts) in the growing season and the bamboo net production is highest in May and October (Nishimura et al. 2004).
The tree and bamboo fine root biomass have been estimated to be 389.8 g m−2 and 88.1 g m−2, respectively, corresponding to 81.6% and 18.4% of total fine-root biomass (tree + bamboo fine roots), respectively (Satomura 2003). The above-ground parts of bamboo are produced mainly in May, while the below-ground parts are produced mainly in October (Nishimura et al. 2004). Yearly total production of the below-ground parts (rhizome + fine roots) of bamboo is roughly 71.1–112.7 g m−2 year−1 (Nishimura et al. 2004).
Tube installation for fine root observation
Eight clear CAB (cellulose acetate butyrate) MR observation tubes with an inside diameter of 4.4 cm, an outside diameter of 5.7 cm and a length of about 90 cm were installed in the main experimental plot in October 1999. The tubes were set along two lines, four tubes in each line at 5-m intervals. The tubes were inserted into the ground at a 45° angle a distance of 60 cm along the axis (42 cm vertical depth). The lower ends of the tubes were sealed to prevent the inflow of water from the soil and the upper ends were covered with opaque PVC (polyvinyl chloride) tubes, the upper end of which were sealed with black sticky tape to prevent the entry of light. All MR tubes had four camera-fixing holes for observations in four directions along the major axis (long axis) of the tube–soil transparent interface.
Fine root observation and image data taking
A MR camera (Minirhizotron BTC-100X; Bartz Technology, Santa Barbara, Calif.) was inserted into each observation tube and images were recorded by a videocassette recorder (Videowalkman GV-D900; Sony, Tokyo, Japan) attached to a monitor along the main axis of the tube using one of the four camera-fixing holes (one soil profile image at each soil depth per tube). In an adjacent cool-temperate deciduous forest, more than 90% of the fine roots were distributed within the upper 20 cm of the soil (Hashimoto and Hyakumachi 1998). Therefore, the soil profile images were taken at depths between 0 and 28 cm at 1-cm intervals along the tube long axis on each observation date. Calibrating the actual soil depth, images were taken between 0 and 20 cm. The ground surface, that is the top of the A0 layer, was assigned a depth of 0. Images were recorded six times during the year of the study: on 19 July (early summer), 25 August (late summer), 4 October (mid-autumn) and 7 November (late autumn) in 2000, and on 26 May (spring) and 21 July (early summer) in 2001. A total of 1,344 images (28 depths per tube × 8 tubes × 6 observation times) were used for the image analysis (see next).
Image analysis
Video images were converted to still pictures and stored in a computer (VAIO PCV-RX65; Sony, Tokyo, Japan) using video-recording software (Giga Pocket ver. 4.5; Sony) through an IEEE 1394 cable. To make fixed-depth images at 5-cm intervals (actual depth), seven sequential still images taken along the tube main axis were combined to form a single image (calibrating the actual soil depth, between 0 and 20 cm at 5-cm intervals) using image-editing software (Photoshop ver. 5.0; Adobe Systems, San Jose, Calif.).
All rootlets in the MR images were categorized according to their demographic features as follows: first, the whole fine roots at the initial observation date were categorized as the initial visible roots. The (initial) visible roots contained both live and dead roots. Over a period of time, some new roots will appear and some will decompose and disappear. The newly appeared roots are called new roots. Roots that had appeared before but were not present at subsequent observation times were called disappeared roots. Roots that were observed again after the first recognition were referred to as remaining roots. Starting with the second observation, the visible fine roots were categorized into two types, namely new roots and remaining roots. Visible roots were then defined as the sum of new roots and remaining roots, which include not only live roots but also dead roots that were not decomposing. Disappeared roots were the roots broken down by the decomposition process (accordingly, disappeared roots are defined as ‘decomposed roots’). The carbon in roots that disappeared over a given period was considered to be put into the soil as organic matter or released to the air as carbon dioxide during the same period.
Two transparent sheets were placed over each combined image generated on a computer with image-processing software (Photoshop ver. 6.0; Adobe Systems). Initial, new, remaining and disappeared fine roots (less than 1 mm in actual diameter) were discriminated by referencing the previous image, and then the new and disappeared fine roots were separately painted manually in red and blue, respectively, on each transparent sheet. The image data were two-dimensional, thus the data obtained directly by the MR method represent the number and length of roots per unit observation area. Each root length was calculated by root image analysis using free software (UTHSCSA Image Tool ver. 2.0; The University of Texas Health Science Center at San Antonio, TX, USA) (http://ddsdx.uthscsa.edu/dig/itdesc.html).
Calculations
Root density (R) was determined as the root length observed in a unit image area (millimeters per centimeter squared). Initial visible root length density, that is the length density of visible roots (Rv) at the initial observation date, was determined by summing the total root length found in a single image (5 cm in depth) at the beginning of the observation period (19 July 2000) and dividing by the total surface area of the observed images, following previous MR studies (Hendrick and Pregitzer 1997; Gill et al. 2002). At the beginning of the study period (July 2000), average values of Rv, which were based on the total length densities of new roots (Rn) and remaining roots (Rr) that could be recognized, were 9.0 mm cm−2 (range 4.7–13.6 mm cm−2) in the depth range 0–5 cm, 2.8 mm cm−2 (0.6–10.5 mm cm−2) in the depth range 5–10 cm, 0.5 mm cm−2 (0.0–1.8 mm cm−2) in the depth range 10–15 cm, and 0.2 mm cm−2 (0.0–0.5 mm cm−2) in the depth range 15–20 cm (n=8 for all depths). Length densities of new and disappeared roots (Rn and Rd) were also determined in the same manner. In each image, the length density of visible roots at time t2 (Rvt2) was calculated using the following equation:
where Rvt1 is the length density of visible roots at time t1, Rnt2 is Rn at time t2 and Rdt2 is Rd at time t2. Rr at time t2 (Rrt2) was also calculated using the following equation:
The rate of root production (millimeters per centimeter squared per month) between times t1 and t2 was defined as Rnt2/(t2−t1). Similarly, the rate of root decomposition (millimeters per centimeter squared per month) was defined as Rdt2/(t2−t1). We calculated the temporary cumulative root production by summing the new root length at this time and that at the previous time (e.g., for time t2 the sum was Rnt1+Rnt2) for each observation time. The temporary cumulative root decomposition was calculated in the same way. The sum of Rn over all observation times was equivalent to the total amount of roots produced over the survey year (yearly total root production, millimeters per centimeter squared per year). Similarly, the sum of Rd over all observation times is equivalent to the total amount of roots decomposed over the survey year (yearly total root decomposition, millimeters per centimeter squared per year).
Root turnover values were calculated from two equations (a root production turnover equation and a root decomposition turnover equation) developed by Gill et al. (2002). In the present study, the term ‘root decomposition’ is used instead of root mortality. Thus, the values of root turnover were classified into two types, that is turnover of root production and turnover of root decomposition. These values were calculated as:
where Rvmax is the maximal Rv during the survey period. More complete definitions of the turnover of root production and the turnover of root decomposition are available in the review by Tingey et al. (2000). We used the concept shown in Fig. 7b in their review.
These calculations were executed based on each 5-cm depth interval per camera-fixing hole of each MR tube. The single-depth data (data from 0–5, 5–10, 10–15, 15–20 cm) obtained by each MR tube were used for the calculation of the site average value (n=8). Values of the yearly total root production and yearly total root decomposition over the survey year and Rvmax in each depth range in each observation tube were used for the calculation (n=8).
Installation of equipment in the soil causes a short-term disturbance. During the first observation year after tube installation, the amount of fine roots and their demography could have been affected around the tubes (Joslin and Wolfe 1999). However, no major differences have been found in the seasonal pattern of the root production rate between the first year and subsequent years in temperate deciduous forests (Baddeley and Watson 2004; Tierney et al. 2001; Burton et al. 2000; Joslin et al. 2001). Thus, we assumed that the seasonal patterns of Rv and root production rate and root decomposition rate could be observed even in the first year after tube installation in the Takayama Experimental Forest. For each tube, single soil cores were taken in July, August and November of 2000 and in July of 2001 at a distance of 2–3 m from the tube (taking care to avoid previous core holes). The first three cores were split into two samples representing depths of 0–5 cm and 5–10 cm. The last core only sampled the depth range 0–5 cm. Thus, a total of 56 soil samples [8 tubes×(3 times×2 depths + 1 time×1 depth)] were collected. A significant correlation was found between the core sampling data (C; grams per meter squared ground area) (Satomura 2003) and the MR data (MR; millimeters per centimeter squared image area; P<0.01, R=0.479, n=56; StatView J-5.0 for Windows; SAS Institute, Cary, N.C., USA). The relationship is expressed as: MR=1.894+0.052C. This suggests that the MR tube installation had little effect on the MR data, at least with respect to the vertical distribution pattern during the observation period. Consequently, we mainly examined the seasonal pattern of root demography as a function of soil depth. The length densities of the fine root production and decomposition and their turnovers are also shown. The estimated turnover values would represent the maximum rates.
Statistical analysis
The effects of observation date (19 July, 25 August, 4 October and 7 November in 2000, 26 May and 21 July in 2001) and soil depth (0–5, 5–10, 10–15, 15–20 cm) on Rv, temporary cumulative root production or temporary cumulative root decomposition were analyzed according to the split-plot design analysis of variance (ANOVA). The effects of observation period (from 19 July to 25 August, from 25 August to 4 October, from 4 October to 7 November, from 7 November 2000 to 26 May 2001, and from 26 May to 21 July) and soil depth (0–5, 5–10, 10–15, 15–20 cm) on the rate of root length production (rate of root production) or the rate of root length decomposition (rate of root decomposition) were also analyzed according to the split-plot design ANOVA. The effect of soil depth (0–5, 5–10, 10–15, 15–20 cm) on the turnover of root production or the turnover of root decomposition was analyzed according to Scheffe’s F-test based on one-way ANOVA. For all such analyses, the 0.01 was added to each value of the visible root length density, the rates of root production and decomposition, root production turnover, and root decomposition turnover to avoid a value of zero and then the data were normalized using the Box-Cox transformation. For each split-plot design ANOVA analysis, the tube was set as a BLOCK, while the observation date (or observation period) and soil depth were defined as the two primary factors.
The difference between turnover values of root production and root decomposition in each soil depth was tested by the t-test. For the t-test, the 0.01 was added to each value of root production turnover and root decomposition turnover to avoid a value of zero and then data were transformed to logarithmic values.
The data were transformed and split-plot design ANOVA was conducted using JMP IN ver. 5.1.1 (SAS). Using StatView J-5.0 for Windows (SAS), correlation between parameters was determined and one-way ANOVA, t-test and linear regression analysis were conducted.
Results
Fine root demography trend along soil depths
The values of fine root demographic parameters decreased with soil depth (except for root production turnover, see below). The depth profiles of these parameters were clearly similar.
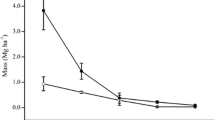
The effect of the soil depth on Rv was significant (split-plot design ANOVA, P<0.0001; Table 1). The yearly average Rv was in the order depth 0–5 cm (11.2 mm cm−2) > 5–10 cm (4.7 mm cm−2) > 10–15 cm (1.4 mm cm−2) > 15–20 cm (0.7 mm cm−2). The Rv within the same soil depth range slightly fluctuated among the observation dates. At depth 0–5 cm, Rv was higher during late summer to late autumn (August 2000, October 2000 and November 2000) and was lower during spring to early summer (July 2000, May 2001 and July 2001; Fig. 2). At the soil surface, the value of Rv in July 2001 was similar to that in July 2000 (Fig. 2). On the other hand, Rv at lower depths (5–10, 10–15, 15–20 cm) tended to gradually increase with time (Fig. 2). Consequently, the average Rv for all depths was highest in late autumn (5.6 mm cm−2 in November 2000) and lowest in early summer (3.1 mm cm−2 in July 2000).

Fluctuations of the density of visible roots (Rv) per unit observation area at soil depths in the range 0–20 cm at 5-cm intervals over a year (July 2000 to July 2001) in a cool-temperate deciduous forest in Japan. Circles 0–5 cm, squares 5–10 cm, diamonds 10–15 cm, triangles 15–20 cm. Bars at each point represent standard errors
The effects of soil depth on the temporary cumulative root production and root decomposition values were also significant (split-plot design ANOVA, P<0.0001 for root production and P<0.0001 for root decomposition; Table 1). The temporary cumulative root production and root decomposition values decreased with increasing soil depth (Fig. 3). They also clearly increased with time (Fig. 3). The average values (±SE) of the yearly total root production at each depth were 16.4±2.7 mm cm−2 year−1 at 0–5 cm, 8.4±1.5 mm cm−2 year−1 at 5–10 cm, 3.0±0.5 mm cm−2 year−1 at 10–15 cm and 1.4±0.6 mm cm−2 year−1 at 15–20 cm. The average (±SE) of the yearly total root decomposition at each depth were 16.5±2.5 mm cm−2 year−1 at 0–5 cm, 5.6±1.9 mm cm−2 year−1 at 5–10 cm, 1.2±0.4 mm cm−2 year−1 at 10–15 cm and 0.3±0.0 mm cm−2 year−1 at 15–20 cm. The yearly total root production tended to be higher than the yearly total root decomposition at the lower soil depths.

Temporary cumulative root production per unit observation area and temporary cumulative root decomposition per unit observation area (mm cm−2) at soil depths in the range 0–20 cm at 5-cm intervals over time in a cool-temperate deciduous forest in Japan. The slope of each line indicates the rate of root production or root decomposition in each period. Bars at each point represent standard errors. Circles 0–5 cm, squares 5–10 cm, diamonds 10–15 cm, triangles 15–20 cm
Both rates of root production and root decomposition clearly decreased with increasing soil depth (Fig. 4). Depth had a significant effect on the rate of root production (split-plot design ANOVA, P<0.0001) and the rate of root decomposition (split-plot design ANOVA, P<0.0001; Table 2). Period had a significant effect on the rates of root production (split-plot design ANOVA, P<0.0001) and root decomposition (split-plot design ANOVA, P=0.0008; Table 2). Fluctuations in these rates tended to be higher at the soil surface than below the surface. These rates were low during late autumn to early spring (November 2000 to May 2001; Fig. 4), although this period was long (more than half a year). Root production in this period accounted for 12.7% of yearly total root production and root decomposition in this period occupied 42.8% of yearly total root decomposition. The root production rate was high in the period from early summer to mid-autumn (July 2000 to October 2000; Fig. 4), which corresponds to the active vegetation season. On the contrary, the root decomposition rate was high in the period from mid-autumn to late autumn (October 2000 to November 2000; Fig. 4), which corresponds to the overstory deciduous tree defoliation season, suggesting that both leaves and fine roots died concurrently during the autumn.

Seasonal pattern of (a) rate of root production and (b) rate of root decomposition per unit observation area at soil depths in the range 0–20 cm at 5-cm intervals during a year (July 2000 to July 2001) in a cool-temperate deciduous forest in Japan. Rates of both root production and root decomposition in each period are plotted for the median date during the period. Bars at each point represent standard errors. Circles 0–5 cm, squares 5–10 cm, diamonds 10–15 cm, triangles 15–20 cm
Fine root turnover
The average turnover values of root production were nearly the same at each depth (0.94–1.22 year−1; Fig. 5). The effect of depth on the turnover of root production was not significant (one-way ANOVA, P=0.12). The average turnover value of root production for all depths was 1.11 mm mm−1 year−1. On the contrary, the average turnover value of root decomposition at each depth clearly decreased with increasing soil depth (Fig. 5). The effect of depth on the turnover of root decomposition was significant (one-way ANOVA, P<0.01). Turnover of root decomposition at different depths was in the order 0–5 cm > 5–10 cm > 10–15 cm > 15–20 cm. The corresponding values were 1.14, 0.73, 0.48 and 0.35 year−1, respectively. The relationship between soil depth, D (cm), and turnover of root decomposition (mm mm−1 year−1) can be expressed by the equation: turnover of root decomposition=1.528−0.409 × ln(D) (P=0.002, R 2=0.995; Fig. 5).

Average fine-root turnover (mm mm−1 year−1) through the soil depth during July 2000 to July 2001 in a cool-temperate deciduous forest at the Takayama survey site in Japan (n=8; horizontal bars standard error). Based on each observation area, production turnover (i.e., yearly total root production/Rvmax) and decomposition turnover (i.e., yearly total root decomposition/Rvmax) were calculated, where Rvmax is the maximum value of visible root length density during the survey year. Values with the same letter are not significantly different among soil depths according to Scheffe’s F-test based on one-way ANOVA (level of significance 0.05). Bars at each point represent standard errors
Significant differences were detected between turnover values of root production and root decomposition at lower depths (10–20 cm) (t-test: P=0.94 for 0–5 cm, P=0.09 for 5–10 cm, P<0.01 for 10–15 cm, P=0.03 for 15–20 cm).
Discussion
Seasonal characteristics in fine root demography
Our results show that the parameters of fine root demography changed with season. Rv peaked after the peak of root production and before the peak of root decomposition (Figs. 2 and 4). It seems reasonable that the root production rate would peak in the growing season and that the root decomposition rate would peak in the latter part of the tree leaf defoliation period in a deciduous forest because the absorption of water and nutrients by fine roots is necessary for vigorous photosynthetic activity of leaves, while considerable amounts of photosynthate are required for growth and maintenance of fine roots. In this case, Rv peaked between the peaks of root production and root decomposition (Figs. 2 and 4). However, in other studies of forest ecosystems, the peak of Rv did not always occur between these two peaks (Table 3). Rv and root production peaked at the same observation time when the observation frequency was low (Baddeley and Watson 2004). In another study, Burton et al. (2000) found that root decomposition peaked before root production at one of four sites and that root production peaked before root decomposition at the other three sites. Each root demographic parameter in forest ecosystems does not always have a single peak (e.g., Burke and Raynal 1994; Joslin et al. 2001). For example, in our study forest, root decomposition rate peaked a second time at the end of spring. Peak patterns found by Baddeley and Watson (2004) and Burton et al. (2000) and our findings of the second peak in the rate of root decomposition imply that the root demographic pattern varies among the forest types. The following points may characterize our study forest: (1) root production rate peaks earlier than root decomposition rate, (2) Rv peaks between the peaks of root production rate and root decomposition rate, (3) root production rate is lower and root decomposition rate is higher in the spring, which results in a lower Rv in spring, (4) root production rate is higher in the latter half of the plant growing season, (5) root decomposition rate is higher at the beginning of the tree dormant season, and (6) Rv is higher in the latter periods of the tree and bamboo growing season.
The fine root production rate peaked in late summer in our study site (Fig. 4). Some other studies have also shown a summer peak in northern hardwood forests (e.g., Burke and Raynal 1994; Fahey and Hughes 1994; Tierney et al. 2001) and an oak-hickory forest (Reich et al. 1980) (Table 3). On the contrary, some studies have shown a spring peak or autumn peak in fine root production rate (Table 3). For example, the root production rate was highest in spring for a northern hardwood deciduous forest dominated by A. saccharum (Hendrick and Pregitzer 1993b) and a mixed woodland dominated by Prunus avium (Baddeley and Watson 2004). In a Quercus-dominated forest, the stand-total fine root production rate peaked in autumn (Cheng and Bledsoe 2002). These differences in the peak seasons of the fine root production rate could be due to factors such as (1) severe root death in winter as a result of soil freezing, (2) severe arid soil conditions in summer, and (3) understory root biomass and dynamics. In the case of soil freezing, root mortality in winter has been shown to be high when the snow pack is removed (Tierney et al. 2001). Subsequently, the root production rate peaks was earlier and more intense in the snow-removal soil than in control soil that had a normal thick snow cover in winter. The earlier intense root production that Tierney et al. (2001) observed may supply enough water and nutrients for rapid growth of leaf biomass. In support of such a balance between root and leaf biomass, Reich et al. (1980) observed alternative repeated leaf expansion and root elongation periods in Q. alba seedlings, whose leaves expanded several times during the growing period. In the case of prolonged summer drought, the peak of leaf photosynthetic activity would be in the spring (Xu and Baldocchi 2003), which could lead to a lower fine root production rate in summer (Edward et al. 2004). However, summer drought was not severe at our site (Fig. 1b). Therefore, the synchronous summer peak of the root production rate and leaf photosynthetic activity rate in our study site is not inconsistent with the findings of previous studies. In the third case, understory plants, which generally show greater leaf photosynthetic activity when the overstory plants have fewer leaves (e.g., Koizumi and Oshima 1985), could affect the stand-total fine root production peak. In a Quercus-dominated forest, Chen and Bledsoe (2002) found an autumn peak in the stand-total (trees + grasses) fine root production rate due to the active fine root production of understory grasses. In their forest, the understory grasses showed a higher fine root production rate in autumn, while overstory trees showed a higher fine root production rate in spring (Table 3). The fine root biomass was considerably greater in the understory grasses than in the overstory trees, which resulted in the autumn peak in the rate of stand-total fine root production (Table 3). As a result, the peak season in our study forest and the temperate deciduous forests in the literature were observed in the growing season (Table 3), while slight differences in the peak seasons (spring, summer or autumn) in these forests would be influenced by environmental factors, species composition and their physiological characteristics.
Seasonal fluctuation pattern in the above-ground and below-ground parameters
In our study forest, the productivity of the overstory deciduous trees is highest at the end of summer (Kawamura et al. 2001; Muraoka and Koizumi 2005), while that of the understory evergreen bamboo is higher in spring and autumn (Nishimura et al. 2004; Sakai and Akiyama 2005) (Fig. 6). The rate of the stand-total fine root increase (synonymous with the root production rate) peaked in late summer and was continuously high during the autumn when the overstory trees lose their leaves (Figs. 4 and 6). In other words, the rate of the stand-total fine root increase was higher when the overstory or understory plants were vigorously leafing and photosynthesizing (Figs. 4 and 6).

Schematic diagrams of the assumed seasonal changes in the above-ground characteristics of plants, the stand-total fine root dynamics and carbon movement through fine root turnover in a layered cool-temperate deciduous forest with a snow cover period (i.e., Betula-dominated forest with understory dwarf bamboo in Takayama, central Japan). a Leaf density or leaf area index of overstory (black), and relative light intensity beneath the overstory (grey) (Kawamura et al. 2001). b Photosynthetic productivity of overstory (black) and understory (grey) plants (Nishimura et al. 2004; Muraoka and Koizumi 2005; Sakai and Akiyama 2005). c Fine root density including both overstory and understory plant roots (Fig. 2). d Rates of fine root increase (black) and fine root decrease (grey); they are evaluated from the stand-total fine root density including both overstory and understory plant roots (Fig. 4)
To verify the relationship between stand productivity and stand-total fine root production rate, stand-level daily NPP (gC m−2 day−1) was estimated by:
where NEP (gC m−2 day−1) is the net ecosystem production taken by a tower-based flux measurement and HR (gC m−2 day−1) is heterotrophic respiration, SR (gC m−2 day−1) is soil respiration (carbon dioxide released from the soil to the air) determined by a chamber method, and RR (gC m−2 day−1) is root respiration estimated by a root trench experiment. During the survey year in the present study, daily estimated NPP values were obtained from the measured values on a total of 43 days using the data from Saigusa, Lee and Mo (personal communications) (relevant data and detail methods were obtained from Saigusa et al. 2002; Lee et al. 2003, 2005; Mo et al. 2005). A significant correlation was found between stand-level daily NPP and stand-total fine root production rate (P<0.0001, R=0.797, n=43). At the stand level, the correlations mentioned here were not in conflict with the hypothesis that the seasonal pattern of the stand-total fine root production rate is associated with the pattern of the stand-level above-ground photosynthetic activity. The rate of increase of the stand-total fine roots was clearly influenced by the photosynthetic productivity of the above-ground parts in both overstory and understory plants.
Takayama Experimental Forest is dominated by the genus Betula species (B. ermanii and B. platyphylla) (Kawamura et al. 2001). In the B. varrucosa- and B. pubescens-dominated forest in Holme Fen, England, the fine root production rate peaks in the late summer and sharply decreases in the deciduous tree defoliation season (autumn) (Ovington and Murray 1968). The peaks of fine root production rate in the Betula-dominated forests in Holme Fen and Takayama occur in the same season. However, fine root production rate is still high in the middle of the tree defoliation season in Takayama, while it immediately decreases at the beginning of the defoliation season in Holme Fen (Ovington and Murray 1968). Because Holme Fen and other forests in England have no bamboo and a lower frequency of evergreen understory plants (Tansley 1965), the higher fine root production in the tree defoliation season in Takayama is considered to be due to the understory evergreen bamboo. Consequently, our study suggests that the plants dominating the understory and their life-history strategy and foliation characteristics affect the productivity of the below-ground parts in forest ecosystems.
Implications of the fine root demography for the forest dynamics
Our results show that not only visible roots (Rv) but also the other fine root parameters (production and decomposition) decrease with increasing soil depths. The seasonal patterns of the fine root demography parameters were almost the same at different soil depths. The fine root production rate in our study site was high not only in summer but also in autumn, presumably due to the presence of understory evergreen bamboo. The seasonality of fine root production rate was supposedly affected by the understory bamboo and the overstory trees. Our findings could suggest that the foliation strategy of the understory plant species has a significant effect on the stand-total fine root demography.
The MR method is a powerful tool for studying seasonal patterns of the fine root demography and for long-term root studies. As shown by our results, fine root decomposition turnover values could differ among soil depths, which suggest that discrimination of soil depths is necessary for studies of fine root dynamics. Long-term root observations would provide a better understanding of soil carbon dynamics through fine roots. The inter-annual fluctuations of the fine root demographic parameters would also be important parameters in the carbon dynamics in forest ecosystems, in the same way that fluctuations in inter-annual leaf demographic parameters are used now.
Abbreviations
- HR:
-
Heterotrophic respiration
- MR:
-
Minirhizotron
- NEP:
-
Net ecosystem production
- NPP:
-
Net primary production
- PDD cycle :
-
Production–death–decomposition cycle
- Rv:
-
Length density of visible roots (mm cm−2 image area)
- Rd:
-
Length density of disappeared roots (i.e., decomposed roots) (mm cm−2 image area)
- Rn:
-
Length density of new roots (i.e., produced roots) (mm cm−2 image area)
- Rr:
-
Length density of remaining roots that have appeared at the previous observation date (mm cm−2 image area)
- RR:
-
Root respiration
- SR:
-
Soil respiration
References
Arnone AJ III, Zaller JG, Spehn EM, Niklaus PA, Wells CE, Körner C (2000) Dynamics of root systems in native grasslands: effects of elevated atmospheric CO2. New Phytol 147:73–85
Baddeley JA, Watson CA (2004) Seasonal patterns of fine-root production and mortality in Prunus avium in Scotland. Can J For Res 34:1534–1537
Burke MK, Raynal DJ (1994) Fine root growth phenology, production, and turnover in a northern hardwood forest ecosystem. Plant Soil 162:135–146
Burton AJ, Pregitzer KS, Hendrick RL (2000) Relationships between fine root dynamics and nitrogen availability in Michigan northern hardwood forests. Oecologia 125:389–399
Cheng X, Bledsoe CS (2002) Contrasting seasonal patterns of fine root production for blue oaks (Quercus douglasii) and annual grasses in California oak woodland. Plant Soil 240:263–274
Comeau PG, Kimmins JP (1989) Above- and below-ground biomass and production of lodgepole pine on sites with different soil moisture regimes. Can J For Res 19:447–454
Edwards EJ, Benham DG, Marland LA, Fitter AH (2004) Root production is determined by radiation flux in a temperate grassland community. Global Change Biol 10:209–227
Eissenstat DM, Caldwell MM (1988) Seasonal timing of root growth in favorable microsites. Ecology 69:870–873
Fahey TJ, Hughes JW (1994) Fine root dynamics in a northern hardwood forest ecosystem, Hubbard Brook Experimental Forest, NH. J Ecol 82:533–548
Fitter AH, Graves JD, Self GK, Brown TK, Bogie DS, Taylor K (1998) Root production, turnover and respiration under two grassland types along an altitudinal gradient: influence of temperature and solar radiation. Oecologia 114:20–30
Gill RA, Burke IC, Lauenroth WK, Milchunas DG (2002) Longevity and turnover of roots in the shortgrass steppe: influence of diameter and depth. Plant Ecol 159:241–251
Grier CC, Vogt KA, Keyes MR, Edmonds RL (1981) Biomass distribution and above- and below-ground production in young and mature Abies amabilis zone ecosystems of the Washington Cascades. Can J For Res 11:155–167
Hashimoto Y, Hyakumachi M (1998) Distribution of ectomycorrhizas and ectomycorrhizal fungal inoculum with soil depth in a birch forest. J For Res 3:243–245
Hendrick RL, Pregitzer KS (1993a) Patterns of fine root mortality in two sugar maple forests. Nature 361:59–61
Hendrick RL, Pregitzer KS (1993b) The dynamics of fine root length, biomass, and nitrogen content in two northern hardwood ecosystems. Can J For Res 23:2507–2520
Hendrick RL, Pregitzer KS (1996) Temporal and depth-related patterns of fine root dynamics in northern hardwood forests. J Ecol 84:167–176
Hendrick RL, Pregitzer KS (1997) The relationship between fine root demography and the soil environment in northern hardwood forests. Ecoscience 4:99–105
Igi S (1991) Geologic atlas of Japan: Chubu region. Asakura, Tokyo, p 56–57
Jia S, Akiyama T, Sakai T, Koizumi H (2002) Study on the carbon dynamics of rhizosphere in a cool-temperate deciduous forest. 1. Relations between topography, vegetation and soil type distribution (in Japanese with English summary). J JASS 18:26–35
Johnson MG, Tingey DT, Phillips DL, Storm MJ (2001) Advancing fine root research with minirhizotrons. Environ Exp Bot 45:263–289
Joslin JD, Wolfe MH (1999) Disturbances during minirhizotron installation can affect root observation data. Soil Sci Soc Am J 63:218–221
Joslin JD, Wolfe MH, Hanson PJ (2000) Effects of altered water regimes on forest root systems. New Phytol 147:117–129
Joslin JD, Wolfe MH, Hanson PJ (2001) Factors controlling the timing of root elongation intensity in a mature upland oak stand. Plant Soil 228:201–212
Kawamura K, Hashimoto Y, Sakai T, Akiyama T (2001) Effects of phenological changes of canopy leaf on the spatial and seasonal variations of understory light environment in a cool-temperate deciduous broadleaved forest (in Japanese with English summary). J Jpn For Soc 83:231–237
Keyes MR, Grier CC (1981) Above- and below-ground net production in 40-year-old Douglas-fir stands on low and high productivity sites. Can J For Res 11:599–605
Koizumi H, Oshima Y (1985) Seasonal changes in photosynthesis of four understory herbs in deciduous forests. Bot Mag Tokyo 98:1–13
Lee M-S, Nakane K, Nakatsubo T, Koizumi H (2003) Seasonal changes in the contribution of root respiration to total soil respiration in a cool-temperate deciduous forest. Plant Soil 255:311–318
Lee M-S, Nakane K, Nakatsubo T, Koizumi H (2005) The importance of root respiration in annual soil carbon fluxes in a cool-temperate deciduous forest. Agric For Meteorol 134:95–101
Lei TT, Mori S, Takahashi K, Koike T (1994) Seasonal photosynthetic patterns of Sasa senanensis in natural open and forest shade sites in Hokkaido, Japan. Bamboo J 12:49–55
Maeno H, Hiura T (2000) The effect of leaf phenology of overstory trees on the reproductive success of an understory shrub, Staphylea bumalda DC. Can J Bot 78:781–785
Miyawaki A, Okuda S (1990) Vegetation of Japan, illustrated. Shibundo, Tokyo
Mo W, Lee M-S, Uchida M, Inatomi M, Saigusa N, Mariko S, Koizumi H (2005) Seasonal and annual variations in soil respiration in a cool-temperate deciduous broad-leaved forest in Japan. Agric For Meteorol 134:81–94
Muraoka H, Koizumi H. (2005) Photosynthetic and structural characteristics of canopy and shrub trees in a cool-temperate deciduous broadleaved forest: implication to the ecosystem carbon gain. Agric For Meteorol 134:39–59
Nishimura N, Matsui Y, Ueyama T, Mo W, Saijo Y, Tsuda S, Yamamoto S, Koizumi H (2004) Evaluation of carbon budgets of a forest floor Sasa senanensis community in a cool-temperate forest ecosystem, central Japan (in Japanese with English summary). Jpn J Ecol 54:143–158
Ohtsuka T, Akiyama T, Hashimoto Y, Inatomi M, Sakai T, Jia S, Mo W, Tsuda S, Koizumi H (2005) Biometric based estimates of net primary production (NPP) in a cool-temperate deciduous forest stand beneath a flux tower. Agric For Meteorol 134:27–38
Ovington JD, Murray G (1968) Seasonal periodicity of root growth of birch trees. In: Ghilarov MS, Kovda VA, Novichkova-Ivanova LN, Rodin LE, Sveshnikova VM (eds) Methods of productivity studies in root systems and rhizosphere organisms. Nauka, Leningrad, pp 146–154
Reich PB, Teskey RO, Johnson PS, Hinckley TM (1980) Periodic root and shoot growth in oak. For Sci 26:590–598
Saigusa N, Yamamoto S, Murayama S, Kondo H, Nishimura N (2002) Gross primary production and net ecosystem exchange of a cool-temperate deciduous forest estimated by the eddy covariance method. Agric For Meteorol 112:203–215
Sakai T, Akiyama T (2005) Quantifying the spatio-temporal variability of net primary production of the understory species, Sasa senanensis, using multipoint measuring techniques. Agric For Meteorol 134:60–69
Sakai T, Jia S, Kawamura K, Akiyama T (2002) Estimation of above-ground biomass and LAI of understory plant (Sasa senanensis) using a hand-held spectro-radiometer (in Japanese with English summary). J Jpn S Photogr Remote Sensing 41:27–35
Satomura T (2003) Biomass of fine roots and mycorrhizal fungi in forest ecosystems. Doctorial Thesis. Graduate School of Biosphere Science, Hiroshima University, Higashi-Hiroshima, Japan (main text in Japanese; summary, figures and tables in English)
Satomura T, Nakane K, Horikoshi T (2001) Analysis of fine-root net primary productivity of trees using minirhizotron method (in Japanese). Root Res 10:3–12
Smit AL, George E, Groenwold J (2000) Root observations and measurements at (transparent) interfaces with soil. In: Smit AL, Bengough AG, Engels C, Van Noordwijk M, Pellerin S, Van de Geijn SC (eds) Root methods. A handbook. Springer, Berlin Heidelberg New York, pp 235–271
Steele SJ, Gower ST, Vogel JG, Norman JM (1997) Root mass, net primary production and turnover in aspen, jack pine and black spruce forests in Saskachewan and Manitoba, Canada. Tree Physiol 17:577–587
Tansley AG (1965) The British islands and their vegetation. Cambridge University Press, Cambridge
Teskey RO, Hinckley TR (1981) Influence of temperature and water potential on root growth of white oak. Physiol Plant 52:363–369
Tierney GL, Fahey TJ, Groffman PM, Hardy JP, Fitzhugh RD, Driscoll CT (2001) Soil freezing alters fine root dynamics in a northern hardwood forest. Biogeochemistry 56:175–190
Tingey DT, Phillips DL, Johnson MG (2000) Elevated CO2 and conifer roots: effects on growth, life span and turnover. New Phytol 147:87–103
Uemura S (1994) Patterns of phenology in forest understory. Can J Bot 72:409–414
Vogt KA, Grier CC, Meier CA, Edmonds RL (1982) Mycorrhizal role in net primary production and nutrient cycling in Abies amabilis ecosystems in western Washington. Ecology 63:370–380
Walter H (1979) Vegetation of the earth and ecological systems of the geo-biosphere. 2nd edn. Springer, Berlin Heidelberg New York
Xu L, Baldocchi DD (2003) Seasonal trends in photosynthetic parameters and stomatal conductance of blue oak (Quercus douglasii) under prolonged summer drought and high temperature. Tree Physiol 23:865–877
Acknowledgements
We are deeply grateful to Mr. K. Kurumado, Mr. N. Miyamoto and other members of Gifu University for their assistance in the field survey. We also thank Dr. M. Inatomi of Frontier Research Center for Global Change and Dr. M. Uchida of National Institute of Polar Research, and for advice and cooperation in the field measurements. Dr. W. Mo and Dr. M.-S. Lee of the River Basin Research Center, Gifu University, and Dr. N. Saigusa of the National Institute of Advanced Industrial Science and Technology, are gratefully acknowledged for providing their data to check the relationship between NPP and fine root production rate. Three anonymous reviewers and Dr. T. Kobayashi of Kagawa University supplied helpful comments and suggestions on earlier versions of the manuscript. This research was partly supported by Grants for the JSPS 21st Century COE Program “Satellite Ecology” of Gifu University and for the JSPS 21st Century COE Program “Biodiversity Research” (A2) of Kyoto University and Sasagawa Scientific Research Grant from the Japan Science Society (no. 13-249).
Author information
Authors and Affiliations
Corresponding author
About this article
Cite this article
Satomura, T., Hashimoto, Y., Koizumi, H. et al. Seasonal patterns of fine root demography in a cool-temperate deciduous forest in central Japan . Ecol Res 21, 741–753 (2006). https://doi.org/10.1007/s11284-006-0182-x
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11284-006-0182-x