
Abstract
During malolactic fermentation (MLF), lactic acid bacteria influence wine aroma and flavour by the production of volatile metabolites and the modification of aroma compounds derived from grapes and yeasts. The present study investigated the impact of different MLF inoculation strategies with two different Oenococcus oeni strains on cool climate Riesling wines and the volatile wine aroma profile. Four different timings were chosen for inoculation with bacteria to conduct MLF in a Riesling must/wine with a high acidity (pH 2.9–3.1). Treatments with simultaneous inoculation showed a reduced total fermentation time (alcoholic and malolactic) compared to the sequential inoculations. No negative impact of simultaneous alcoholic and malolactic fermentation on fermentation success and on the final wine volatile aroma composition was observed. Compared to sequential inoculation, wines with co-inoculation tended to have higher concentrations of ethyl and acetate esters, including acetic acid phenylethylester, acetic acid 3-methylbutylester, butyric acid ethylester, lactic acid ethylester and succinic acid diethylester. Results of this study provide some alternatives to diversify the number of wine styles by safely conducting MLF in low-pH, cool-climate white musts with potential high alcohol content.
Similar content being viewed by others


Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
Malolactic fermentation (MLF), the enzymatic decarboxylation rather of l-malic acid to l-lactic acid and carbon dioxide, is the important secondary fermentation conducted by lactic acid bacteria (LAB) (Versari et al. 1999). The success is influenced by several oenological parameters, such as pH, temperature, alcohol content and sulphur dioxide (SO2) concentration (Lerm et al. 2010). In addition to these parameters, the presence of some yeast inhibitory metabolites such as medium chain fatty acids (Alexandre et al. 2004) or peptic fractions (Nehme et al. 2010) can affect bacterial viability and MLF. Due to these possible antagonistic or undesirable interactions between yeast and bacteria, the correct choice and combination of yeast and bacterial strains is important for the success of MLF.
There are different MLF inoculation possibilities, such as simultaneous inoculation for alcoholic and malolactic fermentation (co-inoculation) with yeasts and LAB, inoculation during alcoholic fermentation (AF) and inoculation after the completion of AF (sequential inoculation) (Fugelsang and Edwards 2007; Henick-Kling 1993). Simultaneous inoculation can be an efficient alternative to overcome potential inhibition of LAB, due to high ethanol concentrations and reduced nutrient content (e.g., essential amino acids, vitamins or minerals) (Jussier et al. 2006; Zapparoli et al. 2009). Hence, a more successful induction of MLF due to a gradual adaption of bacteria to increasing alcohol concentrations and due to the benefit from higher nutrient availability present in the must, compared to the condition at the end of AF (Rosi et al. 2003). Likewise, simultaneous inoculation of musts or wines with high acidity but still low levels of ethanol and higher nutrient concentration may help to avoid potential MLF problems. It also would be beneficial regarding technical aspects: wines after successful co-inoculation would be immediately ready for downstream treatments, such as racking, fining, and SO2 addition, thus increasing microbiological stability and processing efficiency (Jussier et al. 2006). Various studies have been conducted to determine the best moment and condition for bacterial inoculation (Jussier et al. 2006; Massera et al. 2009; Rosi et al. 2003; Semon et al. 2001; Henick-Kling and Park 1994). Jussier et al. (2006) and Massera et al. (2009) observed no negative effects on fermentation success and kinetics, linked with simultaneous inoculation, compared to sequential inoculation and no difference in the final quality of Chardonnay and Malbec wines. The results pointed out the reduction of total fermentation time and a better control of MLF. Rosi et al. (2003) observed that pH and timing of bacterial inoculation were critical to how fast MLF starts. Low pH in a commercial white grape juice had a negative effect on bacterial viability; additionally inoculation halfway through AF caused a bacterial reduction (Rosi et al. 2003).
Most of these studies have concentrated on the microbial interactions, bacterial viability, and only a few wine parameters, such as sugar, malic, citric or acetic acid levels. It has also been observed, that wines that have undergone simultaneous AF/MLF tend to be less buttery and are fruitier with slightly higher but sensorially insignificant levels of acetic acid (Henick-Kling 1993; Bartowsky et al. 2002; Jussier et al. 2006; Krieger 2006; Massera et al. 2009). However, little is known about the influence of the LAB inoculation timing on the volatile aroma composition of the wine, especially in cool climate Riesling wines. Early investigations on sequential MLF in Riesling wines showed intensified fruity and flowery aroma properties of this grape variety (Fischer 1998; Henick-Kling 1993). Moreover, Herjavec et al. (2001) observed that Riesling wines that underwent MLF, were of a better quality than the wines without MLF.
Research, mostly carried out in synthetic wine model solutions, Chardonnay or red wines (e.g. Merlot, Cabernet Sauvignon, Tannat), has shown that LAB have the potential to alter the aroma profile of wine by the production of volatile secondary metabolites or the modification of grape and yeast derived metabolites including ethyl esters, acetate esters, acids and alcohols (de Revel et al. 1999; Hernandez-Orte et al. 2009; Lonvaud-Funel 1999; Maicas et al. 1999; Bartowsky et al. 2010). Many of these alterations are strain dependent, however, the vinification technique can also affect the final wine aroma profile and these flavour impacts of individual bacterial strains are also of great interest for winemakers.
Due to the fact that induced MLF is often difficult to achieve in wines with high acidity, the development of strategies to favour a biological deacidification of low pH wines is important to prevent sluggish or stuck fermentation. Investigations of the present study focused on the impact of different inoculation scenarios with two different O. oeni strains on the volatile wine aroma profile, especially the fruity ester composition. Four different timings were chosen for inoculation with bacteria to conduct MLF. A cool-climate Riesling was chosen as a typical example of a white wine with high acidity.
Materials and methods
Bacterial strains, media and growth conditions
The O. oeni strains R1105 and R1124 (Lallemand, Toulouse, France) were obtained as freeze dried cultures. MRS-T agar (MRS agar containing 10% tomato juice, pH 5) was used for bacterial growth determination. The medium contained 50 mg L−1 Actistab (DSM Food Specalities Dairy Ingredients, Delft, The Netherlands) dissolved in ethanol which inhibited moulds and yeasts. The bacterial cell numbers were determined weekly during MLF. Fermentations were mixed well and samples taken to make a ten-fold dilution series. 100 μL of each dilution was spread plated onto MRS-T agar and the plates were incubated anaerobically at 30°C for 7 days. Colony counts were carried out and reported as colony-forming units per mL (CFU mL−1).
Micro-vinification
Riesling grapes from the Rheingau wine region (Germany) were harvested during the 2010 season. The grapes were destemmed and crushed and a standard addition of 30 mg L−1 sulphite (in form of potassium bisulphite) was added. The must was then settled over night and pasteurised at 82°C for 20 s. Standard juice and wine parameters are shown in Table 1.
After the pH had been adjusted to 3.1 with 5 M NaOH, the must was inoculated with the yeast strain Uvaferm GHM® (Lallemand, Germany) in combination with two different LAB starter cultures and four different inoculation strategies, namely (A) inoculation of LAB starter cultures 24 h after yeast addition; (B) inoculation at 40% of AF; (C) inoculation at 60% of AF and (D) inoculation after the completion of AF (residual sugar < 2 g L−1). The yeast was rehydrated beforehand using Go-Ferm® (Lallemand, Germany) according to the manufactures recommendations. Both O. oeni strains (R1105, R1124) were rehydrated and pre-acclimatised using the 1-Step® protocol (Lallemand) following the manufactures instructions. Both, yeast and bacterial strains were inoculated with approximately 106 CFU mL−1. All fermentations were carried out in triplicate at 20°C in green 2 L bottles with S-shaped airlocks filled with water. The wine with sequential MLF was racked at the end of AF, divided into the 2 L bottles and then inoculated with the bacterial strains.
Alcoholic fermentation (AF) of the must for sequential MLF was monitored by sugar depletion, while the simultaneous fermentations were monitored by CO2 loss. Residual sugar was measured according to Dr. Rebelein (Iland et al. 2004). The wines were considered to be dry and AF concluded when the reducing sugar level was below 2 g L−1. MLF was monitored by malic acid degradation and lactic acid production. MLF was considered complete when malic acid concentration was less than 0.2 g L−1. To each wine sample, 80 mg L−1 of sulphite were added immediately upon completion of MLF. All samples were cold stabilised at 4°C and bottled without filtration and without prior addition of further fining agents.
Analysis of must and wine for organic acids
Must was analysed before inoculation and samples were collected during and after AF and MLF for organic acids (tartaric acid, malic acid, lactic acid, acetic acid and citric acid) using high performance liquid chromatography (HPLC). HPLC analysis was performed according to Schneider et al. (1987) with the following modifications: 5 μL of sample were injected into the Agilent Technologies 1,100 series liquid chromatograph equipped with a multiwavelength detector (MWD) and analysed using an Allure® Organic Acid column (250 × 4.6 mm inside diameter) (Restek GmbH, Bad Homburg, Germany) with a Security Guard™ Cartridge C18 4 × 3 mm (Phenomenex, Aschaffenburg, Germany). The eluent was distilled water with 0.0139% sulphuric acid and 0.5% (v/v) ethanol. The column was operated at 46°C with an eluent flow rate at 0.6 mL/min. Eluting compounds were detected by UV absorbance at 210 nm. Citric acid was also measured enzymatically (R-Biopharm AG, Darmstadt, Germany). Must components are reported as a single value without standard deviation. All other analyses are reported as the means of three determinations (one for each trial carried out in triplicate).
Gas chromatography—mass spectrometry (GC–MS) Analysis for volatile aroma compounds
Higher alcohols, esters, volatile fatty acids and terpenes were analysed using GC–MS. The analysis was performed using a GC Hewlett Packard (HP) 5,890 Series II (Agilent, Santa Clara, USA), coupled to a 5,972 HP Mass Selective Dectector (Agilent). The GC was fitted with a cooled injection system (CIS 3) (Gerstel GmbH, Mülheim, Germany). Compounds were separated on a Varian VF-5MS column (Palo Alto, USA) with dimensions of 60 m × 0.32 mm × 1 μm. Analysis was performed as described previously (Knoll et al. 2011).
Data analysis
Data was subjected to one-way analysis of variance (ANOVA) followed by Tukey’s studentised range (HSD) test to determine whether significant differences between the samples existed, using the SAS Enterprise Guide 4.1 (version 9.1.3, Procedure PROC GLM, SAS Institute, Germany). Differences between samples with a significance level of 5% (P ≤ 0.05) were considered as significant. Multivariate data analysis techniques were used to obtain a more comprehensible overview of the volatile aroma compounds and to investigate possible correlations amongst the analytes. Principal component analysis (PCA) was performed using The Unscrambler software (version 9.2.1, Camo ASA, Norway).
Results
Two O. oeni strains and four different MLF inoculation scenarios were evaluated in a cool climate Riesling must fermented with one yeast strain.
Impact on alcoholic fermentation
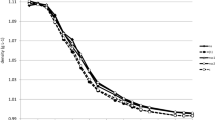
Alcoholic fermentation was completed in 13 or 14 days in all experiments independently of the timing of bacterial inoculation at 24 h, 40% (day 2) and 60% (day 3) of the AF (Fig. 1). The wine inoculated with LAB after completion of AF (EndAF), took as long for the AF as the other treatments inoculated simultaneously. The analytical parameters of the experimental wines at 40, 60% of AF and after completion of AF are shown in Table 1.

Average alcoholic fermentation process of the treatments with bacterial inoculation at 24 h, 40% (day 2) and 60% (day 3) of AF
Effect of bacterial inoculation timing on MLF
Treatments with simultaneous inoculation showed a reduced total fermentation time (AF + MLF) compared to the sequential inoculations. Only, in the 24 h treatment inoculated with R1124 the length of MLF itself in was longer than its respective sequential treatment.
The 24 h treatment was inoculated for MLF at day one of AF, the 40% treatment at day two and the 60% treatment at day three of AF. The 24 h treatment inoculated with O. oeni R1105 took 49 days to complete MLF. O. oeni R1124 took 77 days. The 40% treatment inoculated with O. oeni R1105 took 62 days to conduct MLF and O. oeni R1124 68 days. In the 60% treatment MLF was completed in 49 days (Figs. 2, 3 and 4). The sequential inoculations concluded MLF in 70–84 days (Fig. 5).

Cell numbers (CFU mL−1) of O. oeni R1105 and R1124 (open symbols) and mean values and SD of l-malic acid concentration (filled symbols) during simultaneous fermenation in the treatment with bacterial inoculation after 24 h of AF. Arrow indicates end of AF

Cell numbers (CFU) of O. oeni R1105 and R1124 (open symbols) and mean values and SD of l-malic acid concentration (filled symbols) during simultaneous fermenation in the treatment with bacterial inoculation after 40% of AF. Arrow indicates end of AF

Cell numbers (CFU) of O. oeni R1105 and R1124 (open symbols) and mean values and SD of l-malic acid concentration (filled symbols) during simultaneous fermenation in the treatment with bacterial inoculation after 60% of AF. Arrow indicates end of AF

Cell numbers (CFU) of O. oeni R1105 and R1124 (open symbols) and mean values and SD of l-malic acid concentration (filled symbols) during malolactic fermenation in the treatment with sequential MLF inoculation. Day 0 corresponds to the day of bacterial inoculation
When bacteria were inoculated after 24 h and at 60% of AF, the malic acid decrease began between day 8 and 15 of AF. The population of O. oeni R1105 dropped to 105 CFU mL−1 at both inoculation times. The population of R1124 decreased as low as 103 CFU mL−1 in the 24 h treatment and to 104 CFU mL−1 in the 60% treatment. When bacteria were inoculated at 40% of AF, the population of both strains dropped to 104 CFU mL−1 and it took 14–18 days till malic acid decrease started. From the analytical data recorded in Table 1 it is evident that at 40% of AF the pH was lower compared to the initial value and at 60% of AF. In the wines with sequential inoculation the bacterial population dropped to 103 CFU mL−1 and malic acid degradation commenced after approximately 18 days.
The volatile acidity increased to 0.6–0.7 g L−1 after MLF. However, the concentration in the fermenting musts before inoculation with O. oeni was already 0.5–0.6 g L−1 and in the wine at the end of AF 0.8 g L−1 (Tables 1, 2).
The citric acid, initially present in the grape juice at concentration of 0.25 g L−1, was completely utilized by O. oeni R1124 in all treatments. O. oeni R1105 only partially degraded citric acid in all treatments (Table 2).
Modification of free volatile aroma compounds by MLF
Various volatile components, including alcohols, esters and acids were identified and quantified. A one-way analysis of variance followed by a Tukey’s test was used to detect significant changes in the wine composition. PCA were performed in order to observe underlying trends in the data and to obtain more information about variations in wine composition as a result of different bacterial strains and to compare the influence of the different inoculation timings on MLF. Table 3 lists concentrations of the volatile compounds determined in the wines after alcoholic fermentation and mean values of these compounds after MLF. Results show that MLF and inoculation timing as well as bacterial strains caused significant changes in the volatile aroma composition of the wines.
The concentration of total higher alcohols increased in most treatments after MLF except in the treatment inoculated after 24 h with R1105 and the treatment inoculated at 60% of AF with R1124. The content of hexanol, 3-methylbutanol and 2-phenylethanol increased while the concentration of 2-methylbutanol decreased after MLF.
The content of all acetate esters, except for acetic acid ethylester, decreased after MLF in all treatments. Compared to the co-inoculation, the treatments with sequential inoculation had the lowest concentration of acetic acid phenylethylester (floral, fruity aroma) and acetic acid 3-methylbutylester (banana odour). Moreover, in the wines fermented with O. oeni R1105, higher concentrations of acetic acid 2-methylbutylester and acetic acid 3-methylbutylester were detected.
Quantitatively, levels of short–chain esters such as lactic acid ethylester (milky notes, strawberry), succinic acid diethylester (fruity) and acetic acid ethylester (fruity) were the esters with the main concentration increases during MLF. The treatments with sequential inoculation had the lowest concentration of lactic acid ethylester and succinic acid diethylester, but the highest content of acetic acid ethylester. These compounds were also affected by the bacterial strain. Strain R1105 showed largest increase in lactic acid ethylester and acetic acid ethylester, while the wines inoculated with R1124 at 24 h and 40% of AF the highest content of succinic acid diethylester was detected.
The content of the fruity ethylesters propionic acid ethylester, i-butyric acid ethylester and butyric acid ethylester increased significantly in all wines after MLF. In the wines with sequential MLF the highest concentration of propionic acid ethylester and i-butyric acid ethylester was noted while the butyric acid ethylester showed the lowest increase compared to the wines with simultaneous inoculation. In addition, bacterial strain differences were observed. O. oeni R1105 tended to produce higher concentrations of ethyl esters.
A decrease of the longer chained esters, such as hexanoic-, octanoic- and decanoic acid ethylester, was observed in most wines after MLF. An higher content of hexanoic acid ethylester (green apple) was noted in the wines fermented with R1105.
Figure 6 illustrates the concentration of total ethyl esters (without lactic acid ethylester) and the concentration of total acetate esters (without acetic acid ethylester) in the wines after MLF.

Average concentrations of total ethyl esters (without lactic acid ethylester) and the concentration of total acetate esters (without acetic acid ethylester) at the end of MLF. a Wines inoculated with O. oeni R1105; b Wines inoculated with O. oeni R1124
Volatile fatty acids such as hexanoic and decanoic acid, generally increased in all the wines once MLF had finished while the content of octanoic acid decreased. The lowest concentration of hexanoic acid was found in the wines with sequential inoculation.
A raise of the concentration of the terpenols trans-linalool oxide, cis-linalool oxide, linalool and α-terpineol was observed in all wines after MLF. The wines with sequential MLF had the highest content of trans-linalool oxide, cis-linalool oxide and α-terpineol and total terpenols.
Multivariate data analysis
As an overview of the results, a principal component analysis (PCA) of the volatile aroma compounds of the wines was performed. 81% of the variance was explained by the first two principal components. As shown in Fig. 7a, these PCA’s separated the samples according to inoculation time. Moreover, the treatments inoculated for MLF at 24 h and 40% of AF could be further separated according to the bacterial strain used. Also the wine without MLF is clearly distinguishable from the wines with MLF. On the score plot separation along PC1 was associated with discrimination of treatments inoculated at 40% of AF and at 24 h with R1124 from the treatments inoculated at 60% of AF, at the end of AF and at 24 h with R1105. Loadings for succinic acid diethylester were correlated with treatments inoculated at 40% of AF and at 24 h with R1124, while loadings for acetic acid 3-methylbutylester were correlated with the inoculation at 24 h with R1105 (Fig. 7b).

PCA score plot derived from volatile aroma compounds of all Riesling wines following MLF and the control wine at the end of AF without MLF (no MLF). a Scores plot and b The corresponding loadings plot
Discussion
Successful MLF in cool climate Riesling wines is often difficult to achieve. This study has verified the feasibility of simultaneous inoculation in low pH wines with two suitable yeast-bacterium combinations. The results shown here point out that it is possible to inoculate the bacterial culture at different timings of AF without, on the one hand, inhibiting AF or, on the other hand, causing failure of MLF. However, pH values and timing of bacterial inoculation were shown to be important for how rapidly MLF commences. These results are in agreement with a previous study (Rosi et al. 2003), carried out in a commercial white grape juice, reporting the possible inoculation with LAB at the beginning, middle, and end of AF without slowing down or inhibiting AF or causing failure of MLF. Yet, at pH 3.2 a lowering of bacterial viability was observed (Rosi et al. 2003). In the present study, the co-inoculation at 40% of AF seemed to be the most inhibitory time for malolactic bacteria to start MLF. At this time the pH of the medium was 2.9 which can be associated with production of acids by yeast metabolism. These findings were also noted by Rosi et al. (2003), when bacteria were inoculated halfway through AF. On the other hand, when MLF was carried out at the end of AF, the ethanol content displayed an additional inhibiting factor which delayed the beginning of MLF. Generally, a reduction in total fermentation time was observed when using simultaneous inoculation techniques compared to traditional sequential MLF. The time gained was ranging between 25 and 50 days, depending on the bacterial strain and inoculation time used. This represents an important advantage for the wineries regarding the process efficiency. However, when O. oeni R1124 was used, the length of MLF itself in the treatments with simultaneous inoculation was similar or longer than their respective sequential treatment. O. oeni R1105, on the other hand, carried out MLF faster in the simultaneous treatments than in the consecutive ones and was generally less inhibited by the low pH than R1124. O. oeni R1124 seems to be better suited for a sequential MLF, while strain R1105 can be used for both, co-inoculation and sequential MLF.
The benefits and risks of sequential and simultaneous AF/MLF remain controversial. In this study no negative effect on final wine quality could be substantiated. It was suggested that simultaneous inoculation of yeast and bacteria could result in increased concentration of acetic acid produced by LAB in the presence of available sugars in the must (Davis et al. 1985). In the co-inoculation treatments, levels of acetic acid never exceeded 0.7 g L−1, considering that the wines without bacterial inoculation contained levels between 0.5 and 0.8 g L−1. It can be assumed that in none of the simultaneous fermentations the bacteria produced worrisome levels of acetic acid from sugar and that the yeast metabolism contributed to the elevated acetic acid levels. Thus confirming results of other studies (Massera et al. 2009; Semon et al. 2001; Jussier et al. 2006), demonstrating the possibility of simultaneous fermentation without excessive increase of volatile acidity.
Little is known about the impact of simultaneous inoculation on the production of volatile aroma compounds in Riesling wines. All acetate esters, except acetic acid ethylester, decreased following MLF, while the ethyl esters increased. This is in accordance with previous studies on sequential MLF in red wines (Bartowsky et al. 2008; Ugliano and Moio 2005). Based on sensorial data, it was suggested that Chardonnay, Malbec and Shiraz wines fermented with co-inoculation tend to be fruitier than the wines with sequential inoculation (Bartowsky et al. 2002; Jussier et al. 2006; Massera et al. 2009). In this study the Riesling wines with sequential MLF had the lowest concentration of acetate esters and ethyl esters, most notably due to lower concentrations of acetic acid phenylethylester, acetic acid 3-methylbutylester, butyric acid ethylester, lactic acid ethylester and succinic acid diethylester. This might potentially result in decreased fruitiness in wines with consecutive MLF. The wines with the 24 h inoculation, on the other hand, had the highest concentration of fruity ethyl esters. In addition, changes in the ester concentrations were also affected by the bacterial strain used. O. oeni R1105 seemed to produce higher concentrations of various fruity esters, such as propionic acid ethylester, butyric acid ethylester or lactic acid ethylester, associated with fruitiness, milky notes and mouthfeel, respectively. Increased concentration of these or other esters and strain dependency was also observed by others (Boido et al. 2009; Maicas et al. 1999; Pozo-Bayón et al. 2005; Ugliano and Moio 2005).
Comparison of fermentation-derived compounds from treatments with simultaneous and sequentially inoculated MLF, has illustrated that the profiles of the wines produced, were very different as a result of the MLF inoculation regime and O. oeni strain. The profiles of fermentation-derived compounds of the wines that conducted MLF are clearly distinguishable from those that did not. In addition, wines with complete MLF could be clearly separated according to inoculation timing and distinguished from the wine without MLF. Also, the treatments inoculated for MLF at 24 h and 40% of AF could be further separated according to bacterial strain used.
In conclusion, to our knowledge, for the first time the impact of different bacterial inoculation timings on the MLF performance and on the production of volatile aroma compounds in low pH Riesling must was accomplished. No negative impact of simultaneous AF/MLF on the fermentation success and on the final wine quality was observed. It was demonstrated that inoculation timing and the bacterial strain used can affect the outcome of the final volatile aroma composition of the wine. Applying a co-inoculation protocol may offer microbiological, technological and sensorial advantages, especially in low-pH, cool-climate white musts with potential high alcohol content. However, the success of simultaneous vinification will also depend on the selection of suitable yeast-bacterium combinations (Alexandre et al. 2004).
References
Alexandre H, Costello PJ, Remize F, Guzzo J, Guilloux-Benatier M (2004) Saccharomyces cerevisiae-Oenococcus oeni interactions in wine: current knowledge and perspectives. Int J Food Microbiol 93(2):141–154
Bartowsky EJ, Costello PJ, Henschke PA (2002) Management of malolactic fermentation—wine flavour manipulation. Aust NZ Grapegrow Winemak 461:7–8 and 10–12
Bartowsky EJ, Costello PJ, McCarthy J (2008) MLF-adding an ‘extra dimension’ to wine flavour and quality. Aust NZ Grapegrow Winemak 533a:60–65
Bartowsky EJ, Costello PJ, Krieger S, Markides A, Leigh F, Travis B (2010) Influence of malolactic fermentation on the fruity characters of red wine—bringing wine chemistry and sensory together. In: Deutscher Weinbauverband e.V. (ed) Proceedings of International Intervitis Interfructa Congress. Stuttgart, Germany. http://shop.dwv-online.de/product_info.php?products_id=47
Boido E, Medina K, Faria L, Carrau F, Versini G, Dellacassa E (2009) The effect of bacterial strain and aging on the secondary volatile metabolites produced during malolactic fermentation of Tannat red wine. J Agric Food Chem 57(14):6271–6278
Davis CR, Wibowo D, Eschenbruch R, Lee TH, Fleet GH (1985) Practical implications of malolactic fermentation: a review. Am J Enol Vitic 36(4):290–301
de Revel G, Martin N, Pripis-Nicolau L, Lonvaud-Funel A, Bertrand A (1999) Contribution to the knowledge of malolactic fermentation influence on wine aroma. J Agric Food Chem 47(10):4003–4008
Fischer U (1998) Sensorische Aspekte des biologischen Säureabbaus Der Deutsche Weinbau 24:12–15
Fugelsang KC, Edwards CG (eds) (2007) Wine Microbiology. In: Lactic acid bacteria, 2nd edn. Springer Science + Business Media, New York
Henick-Kling T (1993) Malolactic fermentation. In: Fleet GH (ed) Wine Microbiology and Biotechnology. Harwood Academic Publishers, Chur Switzerland, pp 286–326
Henick-Kling T, Park YH (1994) Considerations for the use of yeast and bacterial starter cultures: SO2 and timing of inoculation. Am J Enol Vitic 45(4):464–469
Herjavec S, Tupajić P, Majdak A (2001) Influence of malolactic fermentation on the quality of Riesling wine. ACS 66(1):59–64
Hernandez-Orte P, Cersosimo M, Loscos N, Cacho J, Garcia-Moruno E, Ferreira V (2009) Aroma development from non-floral grape precursors by wine lactic acid bacteria. Food Res Int 42:773–781
Iland P, Bruer N, Edwards G, Weeks S, Wilkes E (eds) (2004) Chemical analysis of grapes and wine: techniques and concepts. In: Patrick Iland Wine Promotions Pty/Lltd, Campbell Town, South Australia
Jussier D, Dube Morneau A, Mira de Orduna R (2006) Effect of simultaneous inoculation with yeast and bacteria on fermentation kinetics and key wine parameters of cool-climate Chardonnay. Appl Environ Microbiol 72(1):221–227
Knoll C, Fritsch S, Schnell S, Grossmann M, Rauhut D, du Toit M (2011) Influence of pH and ethanol on malolactic fermentation and volatile aroma compound composition in white wines. LWT Food Sci Technol 44(2011):2077–2086
Krieger S (2006) Determining when to add malolactic bacteria. In: Morenzoni R (ed) Malolactic fermentation in wine—understanding the science and the practice. Lallemand, Montréal, Canada, pp 12.01–12.10
Lerm E, Engelbrecht L, du Toit M (2010) Malolactic fermentation: the ABC’s of MLF. S Afr J Enol Vitic 31(2):186–212
Lonvaud-Funel A (1999) Lactic acid bacteria in the quality improvement and depreciation of wine. Antonie van Leeuwenhoek 76:317–331
Maicas S, Gil J-V, Pardo I, Ferrer S (1999) Improvement of volatile composition of wines by controlled addition of malolactic bacteria. Food Res Int 32(7):491–496
Massera A, Soria A, Catania C, Krieger S, Combina M (2009) Simultaneous inoculation of Malbec (Vitis vinifera) musts with yeast and bacteria: effects on fermentation performance, sensory and sanitary attributes of wines. Wine Food Technol Biotechnol 47(2):192–201
Nehme N, Mathieu F, Taillandier P (2010) Impact of the co-culture of Saccharomyces cerevisiae-Oenococcus oeni on malolactic fermentation and partial characterization of a yeast-derived inhibitory peptidic fraction. Food Microbiol 27(1):150–157
Pozo-Bayón MA, G-Alegria E, Polo MC, Tenorio C, Martin-Alvarez PJ, CalvodelaBanda MT, Ruiz-Larrea F, Moreno-Arribas MV (2005) Wine volatile and amino acid composition after malolactic fermentation: effect of Oenococcus oeni and Lactobacillus plantarum starter cultures. J Agric Food Chem 53(22):8729–8735
Rosi I, Fia G, Canuti V (2003) Influence of different pH values and inoculation time on the growth and malolatic activity of a strain of Oenococcus oeni. Aust J Grape Wine Res 9:194–199
Schneider A, Gerbi V, Redoglia M (1987) A rapid HPLC method for separation and determination of major organic acids in grape musts and wines. Am J Enol Vitic 38(2):151–155
Semon MJ, Edwards CG, Forsyth D, Dinn C (2001) Inducing malolactic fermentation in Chardonnay musts and wines using different strains of Oenococcus oeni. Aust J Grape Wine Res 7(1):52–59
Ugliano M, Moio L (2005) Changes in the concentration of yeast-derived volatile compounds of red wine during malolactic fermentation with four commercial starter cultures of Oenococcus oeni. J Agric Food Chem 53(26):10134–10139
Versari A, Parpinello GP, Cattaneo M (1999) Leuconostoc oenos and malolactic fermentation in wine: a review. J Ind Microbiol 23(6):447–455
Zapparoli G, Tosi E, Azzolini M, Vagnoli P, Krieger S (2009) Bacterial inoculation strategies for the achievement of malolactic fermentation in high-alcohol wines. S Afr J Enol Vitic 30(1):49–55
Acknowledgment
We wish to thank Anna Dalljo for her technical assistance.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Knoll, C., Fritsch, S., Schnell, S. et al. Impact of different malolactic fermentation inoculation scenarios on Riesling wine aroma. World J Microbiol Biotechnol 28, 1143–1153 (2012). https://doi.org/10.1007/s11274-011-0917-x
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11274-011-0917-x