
Abstract
Curculigo orchioides Gaertn. (family Hypoxidaceae) is an endangered anticarcinogenic and aphrodisiac herb, native of India. This study reports the effect of three arbuscular mycorrhizal (AM) fungal inocula on post-transplanting performance of ‘in vitro’ raised C. orchioides plantlets. The three AM fungal inocula consisted of two monospecific cultures of Glomus geosporum and G. microcarpum and one crude consortium of AM fungal spores isolated from rhizosphere soil of C. orchioides growing in natural habitat. Complete plantlets of C. orchioides were raised by direct organogenesis of leaf explants on half strength Murashige and Skoog’s medium devoid of any growth hormone. C. orchioides plantlets responded significantly different to all three mycorrhizal treatments. Mycorrhization enhanced the survival rate of C. orchioides plantlets to 100%. The inoculated plantlets fared significantly better than the uninoculated ones in terms of biomass production and number of leaves and roots per plant. Mycorrhizal plantlets exhibited higher concentrations of photosynthetic pigments as well as minerals P, Mg, Cu, Zn, Mn and Fe in both shoots and roots. Among the three inocula tested, plantlets inoculated with the mixed consortium of AM fungi consistently performed better in terms of the parameters evaluated. The study suggests use of mixed consortium of AM fungi over monospecific cultures for the sustainable cultivation and conservation of endangered medicinal plant: Curculigo orchioides.
Similar content being viewed by others


Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
Curculigo orchioides Gaertn. is an endangered medicinal herb belonging to family Hypoxidaceae. It is an important medicinal herb by virtue of the anticancerous properties of its rhizomes (Prajapati et al. 2003). The species is native to India and has wide distribution in the sub-tropical Himalayas from Kumaon eastwards and in the Western Ghats from Konkan southwards (Bafna and Mishra 2005).
Curculigo orchioides can be propagated in nature by seeds or rhizomes. However, the plant shows poor seed set and germination, and high incidence of bacterial and viral diseases of rhizome under natural conditions (Wala and Jasrai 2003). In addition to these, lack of organized culture and over-exploitation of tuberous roots are amongst the other factors responsible for the declining population of this plant in the wild. C. orchioides grows during rainy season and can be harvested for drug preparation only during a very short period (Vaidya et al. 2005). Therefore, to ensure the availability of C. orchioides throughout the year, micropropagation technology becomes the most viable option for the conservation of this valuable herb. Although micropropagation protocols for this plant have been standardized, limited information is available on the performance of C. orchioides plantlets after transplantation (Prajapati et al. 2003; Vaidya et al. 2005).
The potential of arbuscular mycorrhizal (AM) fungi as bioprotectors and biofertilizers has been well documented in earlier studies (Kapoor et al. 2002). The major beneficial effects associated with the use of AM fungi include: better nutrient acquisition (Kapoor et al. 2004), improved plant water relations, tolerance to abiotic stresses (Kapoor and Bhatnagar 2007), such as drought (Giri et al. 2005) and salinity (Giri et al. 2007), resistance to plant pathogens and tolerance to transplantation shock experienced by micropropagated plant species at the time of their transplantation to the field. AM inoculation and colonization of tissue-cultured plantlets of other plant species have been reported to reduce transplantation shock during acclimatization, thus increasing plant survival and establishment rates (Estrada-Luna et al. 2000). The benefits associated with the use of AM inoculation for ‘in vitro’ raised plantlets have been reported in several horticultural and forest tree species (Rai 2001). However, little is known about the influence of AM fungi on survival and growth of micropropagated medicinal plant species. The objective of this research was to determine whether AM inoculation could enhance the survival and post-transplantation performance of ‘in vitro’ raised plantlets of C. orchioides and thus help in the conservation and sustainable utilization of this endangered plant for medicinal purposes.
Materials and methods
Plant material and micropropagation procedures
Curculigo orchioides plants along with the rhizomes and roots were collected from Western Ghats (India) and transplanted in pots in the Botanical Garden, University of Delhi. The young leaves (4–6 cm length) of these plants served as source of explants for micropropagation.
Complete plantlets of C. orchioides consisting of well developed roots and leaves were obtained by direct organogenesis of leaf explants (8–10 mm length) on half strength MS (Murashige and Skoog 1962) medium containing 3% sucrose and 0.2% activated charcoal following modified protocol by Prajapati et al. (2003). All cultures were maintained in a culture room at 24°C under 16 h light and 8 h dark cycle with a light intensity of 50 μmol m−2 s−2 provided by cool white fluorescent lamps (Philips) with 55–60% relative humidity.
The rooted micropropagated plantlets were removed from the agar and transplanted in thermocol cups containing moist cocopeat supplemented with organic manure (1:10) for acclimatization. The cups were covered with transparent polythene bags (to maintain humidity) with holes (to provide aeration) and grown for 15 days under the same light and temperature conditions provided during micropropagation.
After 15 days of acclimatization, C. orchioides plantlets with approximately same growth (equal number and same size of leaves and roots) were selected for transplantation studies.
Arbuscular mycorrhizal inoculum
The arbuscular mycorrhizal (AM) spores present in rhizosphere soil of Curculigo orchioides plants collected from six distantly located sites in the Western Ghats (India) were isolated using wet sieving and decanting technique (Gerdemann and Nicolson 1963) and identified with the help of standard manual by Schenck and Perez (1990). The AM spores belonging to Glomus geosporum and G. microcarpum were most abundant in all the rhizosphere soils analyzed. Thus, monospecific cultures of G. geosporum (Nicolson & Gerdemann) Walker and G. microcarpum Tulasne & Tulasne were raised from a single viable spore respectively on Sorghum halepense roots (Brundett and Juniper 1995). A crude mixed consortium of ten AM fungal spores was multiplied from rhizosphere soil collected from a site in Western Ghats using S. halepense as the trap plant. The crude mixed consortium consisted of spores of Acaulospora laevis Gerdemann & Trappe, Gigaspora albida Schenck & Smith, Gigaspora sp. (Probably ‘in. det.’), Glomus etunicatum Becker & Gerdemann, G. fasciculatum (Thaxter) Gerdemann & Trappe emend. Walker & Koske, G. fuegianum (Spegazzini) Trappe & Gerdemann, G. geosporum (Nicolson & Gerdemann) Walker, G. microcarpum Tulasne & Tulasne, G. mosseae (Nicolson & Gerdemann) Gerdmann & Trappe and G. reticulatum Bhattacharjee & Mukerji.
Experimental design
The experiment consisted of four treatments arranged in a completely randomized design. The four mycorrhizal treatments were viz., (i) control (autoclaved sterile soil) (ii) inoculation with Glomus geosporum (GG), (iii) inoculation with Glomus microcarpum (GM), and (iv) inoculation with crude consortium of AM fungi (MC). Each treatment consisted of 20 pots, one plantlet per pot. The acclimatized plantlets were transplanted into individual plastic pots (9.5 × 6.5 × 9.5 cm) containing sterilized autoclaved soil. Mycorrhizal treatments were at the level of 150 infectious propagules per plant. The inoculum was mixed uniformly with soil in each pot. The plantlets were allowed to grow and no pesticide or fertilizer was added during the course of the experiment.
Plant growth measurements and nutrient concentration
Three randomly chosen plantlets (four-months-old) from each treatment were harvested along with complete roots. Plants were separated just above the crown into roots and shoots, and their number and fresh weights were determined. Dry weight of shoots and roots were determined after drying the tissue in an oven at 80°C for 48–72 h. The 0.2 g oven-dried ground material was digested in a triple acid mixture (HN03, H2SO4 and 60% HClO4 in a ratio of 10:1:4) and used for nutrient analysis. The phosphorus in the digested sample was estimated by the molybdenum blue method (Allen 1989). The concentrations of Mg, Cu, Zn, Mn and Fe were determined using atomic absorption spectrophotometry following Allen 1989. Working standards were procured from Sigma Aldrich, USA.
Photosynthetic leaf pigments concentration (mg/g fresh weight)
Leaf chlorophyll was determined by Hiscox and Israelstam’s (1979) method. The concentrations of chl a, chl b and carotenoids were calculated by Arnon’s formulae (Arnon 1949).
Statistical analyses
Unifactorial analysis of variance (ANOVA) was performed on each variable. Tukey’s Post Hoc multiple mean comparison test was used to test for significant differences between treatments (at 5% level). All statistical analyses were performed with Statistical Package for Social Sciences (SPSS, version 10).
Results and discussion
All the mycorrhizal plantlets of Curculigo orchioides survived transplantation shock as compared to 70% survival rate in non-mycorrhizal control plantlets. The observations of the present study are consistent with the previous reports on other plant species (Gaur and Adholeya, 1999).
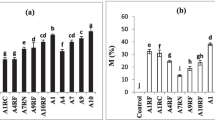
At the time of harvest (four months after transplantation), mycorrhizal plantlets had better growth and leaf production than non-mycorrhizal plantlets, however the magnitude of response varied with mycorrhizal treatment (Table 1). Mycorrhizal symbiosis enhanced number of roots per plant, root fresh weight and dry weight significantly. The increase in shoot dry weight on mycorrhization was statistically insignificant (P ≤ 0.05).
High mortality in micropropagated plantlets is primarily due to a poorly developed or non-functional root system (Wang et al. 1993). Increase in survival of micropropagated mycorrhizal C. orchioides plantlets may be attributed to better root development. This is further supported by higher root/shoot ratio in mycorrhizal plantlets (Table 1). Higher root/shoot ratios indicate highly developed and extensive root system with efficient nutrient absorption (Clapperton and Reid 1992).
Mycorrhizal plantlets had higher P concentrations in the roots as well as shoots in comparison to non-mycorrhizal plantlets (Table 2). Assessment of other nutrients indicated that mycorrhizal colonization altered the acquisition of these nutrient elements. Mycorrhizal plantlets contained significantly higher concentrations of Cu, Zn, Mg, Mn and Fe in comparison to non-mycorrhizal plantlets in both root as well shoot tissues.
In most cases, except for Cu in MC and GG plantlets, and Zn in GM inoculated plantlets, the root/shoot ratios of micronutrient (Cu, Mn, Zn and Fe) concentrations were higher in mycorrhizal plantlets (Table 3). This suggests greater accumulation of these elements in roots than in shoots of mycorrhizal plantlets as compared to non-mycorrhizal control plantlets, reflecting a change in nutrient allocation resulting from mycorrhizal fungal inoculation. AM fungal inoculation improves the uptake of Cu, Zn, Mn and Fe but at the same time protect shoots from their excess (Sylvia and Williams 1992). However, there was no change in the distribution of Mg in mycorrhizal and non-mycorrhizal plantlets. This uptake selectivity reflects a well-regulated nutrient homeostasis in mycorrhizal plants (Ning and Cumming 2001), that helps micropropagated plantlets withstand nutrient stress on transplantation.
Leaf photosynthetic pigment concentrations differed among the mycorrhizal treatments (Table 4). Overall, a considerable increase was recorded in leaf chlorophyll concentration of mycorrhizal plantlets. As much as 1.52-fold concentration of chl a, 1.36-fold concentration of chl b and 1.43-fold concentration of carotenoids were obtained in leaves of MC inoculated plantlets over their respective concentrations in leaves of control plantlets. These results are in accordance with the results obtained in earlier studies (Estrada-Luna et al. 2000; Estrada-Luna and Davies Jr. 2003). Mg and Cu are essential components of chlorophyll molecule and chlorophyll protein plastocyanin (Taiz and Zeiger 1998). Thus, the enhancement in concentration of photosynthetic pigments observed in the present study may be attributed to increased concentrations of Mg and Cu in leaves of AM-inoculated plantlets of C. orchioides in comparison to the non-inoculated control plantlets. Micropropagated plantlets typically have leaves with low chlorophyll content and low photosynthetic rates that impede growth (Grout and Millan 1985). Improved growth and survivability of mycorrhizal C. orchioides plantlets in this study may be due to an increase in chlorophyll concentration in their leaves.
Different plant species within natural communities have different AM fungal assemblages on their roots and the occurrence and abundance of a vascular plant species in a particular community may depend on the presence of one to several specific arbuscular mycorrhizal fungi (van der Heijden et al. 1998). Certain combinations of host and fungus are more or less compatible than others (Parke and Kaeppler 2000; Ravnskov and Jakobsen 1995). The species and strains of AM fungi vary in their capacity to take up P from soil and transfer it to the host plant (Smith et al. 2000). Furthur, it is acknowledged that there is high functional diversity among AM fungal species (Munkrold et al. 2004). These fungi are multifunctional and there may exist trade-off among fungi in their abilities to perform these functions. The fact that AM fungi differ with respect to function, and because plants differ in their response to AM fungi, connote a benefit to systems with a high AM fungal species richness; more AM fungal species means more functions fulfilled and more opportunities for beneficial relationships to develop. This could be the reason that in the present study, among various mycorrhizal treatments tested, inoculation with a mixed consortium of ten species of AM fungal spores (MC) proved to be more beneficial than the two monospecific cultures.
In conclusion, arbuscular mycorrhizal inoculation ensures maximal survival and growth of micropropagated Curculigo orchioides plantlets after out planting. The association of AM fungi with the roots of C. orchioides plants not only influence P acquisition but other nutrients such as Mg, Cu, Zn, Fe and Mn that have poor mobility rates. Improved root development, enhanced levels of photosynthetic pigments are some of the indirect effects of mycorrhization that help micropropagated C. orchioides plantlets overcome transplantation shock and increase plant growth.
The study highlights the use of crude mixed consortium of AM fungal spores over monospecific cultures for improving the post-transplantation performance of ‘in vitro’ raised plantlets of C. orchioides. Though improved survival and establishment of micropropagated plantlets have been reported by using monospecific cultures of AM fungal spores in previous studies (Estrada-Luna & Davies Jr. 2003; Jaizme-Vega et al. 2003). The procedure requires isolation and sterilization of AM fungal spores, making the technology lengthy, time-consuming and uneconomical for large scale transplantations. In contrast, the methodology followed during the present investigation is a single-step process, involving simply laying of a crude consortium of AM fungal spores multiplied on a suitable trap plant, at the time of transplantation of micropropagated plantlets. This technology may also be extended to other such medicinally important plants which require conservation efforts.
Abbreviations
- AM:
-
Arbuscular mycorrhiza
- GG:
-
Glomus geosporum
- GM:
-
Glomus microcarpum
- MC:
-
Mixed consortium
References
Allen SE (1989) Chemical analysis of ecological materials. 2nd edn. Blackwell Scientific Publishers, UK
Arnon DJ (1949) Copper enzyme in isolated chloroplasts polyphenol oxidase in Beta vulgaris. Plant Physiol 24:1–15
Bafna AR, Mishra SH (2005) Actividad antioxidante in vitro del extracto de methanol de los rizomas de Curculigo orchioides Gaertn. Ars Pharm 46(2):125–138
Brundrett M, Juniper S (1995) Non-destructive assessment of spore germination of VAM fungi and production of pot cultures from single spores. Soil Biol Biochem 27:85–91
Clapperton MJ, Reid DM (1992) A relationship between plant growth and increasing VA mycorrhizal inoculum density. New Phytol 120:227–234
Estrada-Luna AA, Davies FT (2003) Arbuscular mycorrhizal fungi influence water relations, gas exchange, abscissic acid and growth of micropropagated chile ancho pepper (Capsicum annum) plantlets during acclimatization and post-acclimatization. J Plant Physiol 160:1073–1083
Estrada-Luna AA, Davies FT Jr, Egilla JN (2000) Mycorrhizal fungi enhancement of growth and gas exchange of micropropagated guava (Psidium guajava L.) during ex vitro acclimatization and plant establishment. Mycorrhiza 10:1–8
Gaur A, Adholeya A (1999) Mycorrhizal effects on the acclimatization, survival, growth and chlorophyll of micropropagated Syngonium and Draceana inoculated at weaning and hardening stages. Mycorrhiza 9:215–219
Gerdemann JW, Nicolson TH (1963) Spores of mycorrhizal species extracted from soil by wet sieving and decanting method. Trans Brit Mycol Soc 46:235–244
Giri B, Kapoor R, Mukerji KG (2007) Improved tolerance of Acacia nilotica to salt stress by arbuscular mycorrhiza, Glomus fasciculatum is partly related to elevated K: Na ratios in root and shoot tissues. Micro Ecol (in press)
Giri B, Kapoor R, Mukerji KG (2005) Effect of arbuscular mycorrhiza Glomus fasciculatum and G. macrocarpum on the growth and nutrient content of Cassia siamea in semi-arid Indian wasteland soil. New Forest 29:63–73
Grout BWW, Millan S (1985) Photosynthetic development of micropropagated strawberry plantlets following transplanting. Ann Bot 55:129–131
Hiscox JD, Israelstam CP (1979) A method for extraction of chlorophyll from leaf tissue without maceration. Can J Bot 57:1331–1334
Jaizme-Vega MC, Rodriguez-Romero AS, Hermoso CM et al (2003) Growth of micropropagated bananas colonized by root-organ culture produced arbuscular mycorrhizal fungi entrapped in Ca-alginate beads. Plant Soil 254:329–335
Kapoor R, Bhatnagar AK (2007) Attenuation of cadmium toxicity in mycorrhizal Celery (Apium graveolens L.). World J Microbiol Biotechnol DOI 10.1007/s11274-006-9337-8
Kapoor R, Giri B, Mukerji KG (2002) Glomus macrocarpum: a potential bioinoculant to improve essential oil quality and concentration in dill (Anethum graveolens L.) and carum (Trachyspermum ammi (Linn.) Sprague). World J Microbiol Biotechnol 18:459–463
Kapoor R, Giri B, Mukerji KG (2004) Improved growth and essential oil yield and quality in Foeniculum vulgare Mill on mycorrhizal inoculation supplemented with P-fertilizer. Bioresource Technol 93:309–311
Munkrold L, Kjoller R, Vestberg M et al (2004) High functional diversity within species of arbuscular mycorrhizal fungi. New Phytol 164:357–364
Murashige T, Skoog F (1962) A revised medium for rapid growth and bioassays with tobacco tissue cultures. Physiol Plant 15:473–497
Ning J, Cumming JR (2001) Arbuscular mycorrhizal fungi alter phosphorus relations of broomsedge (Andropogon virginicus L.) plants. J Exp Bot 52:1883–1891
Parke JL, Kaeppler SW (2000) Effects of genetic differences among crop species and cultivars upon the arbuscular mycorrhizal symbiosis. In: Kapulnik K, Douds DD (eds) Arbuscular mycorrhizas: physiology and function. Kluwer, The Netherlands, pp 131–146
Prajapati HA, Patel DH, Mehta SP et al (2003) Direct in vitro regeneration of Curculigo orchioides Gaertn., an endangered anticarcinogenic herb. Curr Sci 84:747–749
Rai MK (2001) Current advances in mycorrhization in micropropagation. In Vitro Cell Dev Biol-Plant 37:158–167
Ravnskov S, Jakobsen I (1995) Functional compatibility in arbuscular mycorrhizas measured as hyphae P transport to the plant. New Phytol 129:611–618
Schenck NC, Perez Y (1990) Manual for the Identification of VA Mycorrhizal fungi
Smith FA, Jakobsen I, Smith SE (2000) Spatial differences in acquisition of soil phosphate between two arbuscular mycorrhizal fungi in symbiosis with Medicago truncatula. New Phytol 147:357–366
Sylvia DM, Williams SE (1992) Vesicular-arbuscular mycorrhizae and environmental stress. In: Bethlenfalvay GJ, Linderman RG (eds) Mycorrhizae in sustainable agriculture. Special Publication 54, ASA, Madison, Wisc, pp 101–124
Taiz L, Zeiger E (1998) Plant Physiology. Second ed., Sinauer Assoc. Inc. Pub., USA
Vaidya GK, D’Cruz L, Saxena OP et al (2005) Effect of various factors on plantlet regeneration of Curculigo orchioides Gaertn. Plant Cell Biotech Mol Biol 6:29–34
van der Heijen MGA, Klironomos JN, Moutoglis P et al (1998) Mycorrhizal fungal diversity determines plant biodiversity, ecosystem variability and productivity. Nature 396:69–72
Wala BB, Jasrai YT (2003) Micropropagtion of an endangered medicinal plant: Curculigo orchioides Gaertn. Plant Tissue Cult 13:13–19
Wang H, Parent S, Gosselin A et al (1993) Study of vesicular-arbuscular mycorrhizal peat-based substrates on symbiosis establishment, acclimatization and growth of three micropropagated species. J Am Soc Hort Sci 118:896–901
Acknowledgements
This research was funded by Department of Science and Technology, New Delhi, India. The authors are thankful to Prof. S. R. Yadav for helping in collection of the plant material.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Sharma, D., Kapoor, R. & Bhatnagar, A.K. Arbuscular mycorrhizal (AM) technology for the conservation of Curculigo orchioides Gaertn.: an endangered medicinal herb. World J Microbiol Biotechnol 24, 395–400 (2008). https://doi.org/10.1007/s11274-007-9488-2
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11274-007-9488-2