
Abstract
Alteration of natural flow regime is considered a major threat to biodiversity in river floodplain ecosystems. Measurements of quantitative relationships between flow regime change and biodiversity are, however, incomplete and inconclusive. This hampers the assessment of human impact on riverine floodplain wetlands in global biodiversity evaluations. We systematically reviewed the scientific literature and extracted information from existing data sets for a meta-analysis to unravel a general quantitative understanding of the ecological consequences of altered flow regimes. From 28 studies we retrieved both ecological and hydrological data. Relative mean abundance of original species (mean species abundance, MSA) and relative species richness were used as effect size measures of biodiversity intactness. The meta-analysis showed that alteration of a natural flow regime reduces the MSA by more than 50 % on average, and species richness by more than 25 %. Impact on species richness and abundance tends to be related to the degree of hydrological alteration. These results can be used in strategic quantitative assessments by incorporating the relationships into global models on environmental change and biodiversity such as GLOBIO-aquatic.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Natural floodplains in river basins are among the most biologically diverse and productive ecosystems on the planet (Ward et al. 1999; Tockner and Stanford 2002). The natural river flow regime is considered the key factor driving ecological functioning and biodiversity patterns in floodplain ecosystems (Junk et al. 1989; Bunn and Arthington 2002). Flood pulses actively shape the floodplain surface, creating a spatio-temporal heterogeneous environment supporting a great variety of habitats (Ward et al. 1999; Tockner et al. 2010; Davidson et al. 2012).
Like other freshwater wetlands, river floodplains are among the most threatened ecosystems and over the past 30 years species diversity has declined faster here than in terrestrial or marine ecosystems (Loh and Wackernagel 2004; MEA 2005; Revenga et al. 2005). Alteration of the flow regime is generally considered one of the most serious and continuing human threats to the ecological intactness of these ecosystems (Poff et al. 1997; Postel and Richter 2003; Poff et al. 2007).
Everywhere, rivers have been regulated for various purposes, including public water supply, irrigation, navigation, flood mitigation and electricity generation, thereby contributing considerably to human development and economic prosperity (Dynesius and Nilsson 1994). As a consequence, over 60 % of global river systems have been affected by altered stream flows (Revenga et al. 2000), and this figure is projected to increase (World Wildlife Fund 2004). Damming is one of the most common types of water flow management, and has profound effects on river hydrology (Middleton 2002), affecting both the total discharge and the variability (i.e. frequency, duration, timing, magnitude and rate of change) of flow (Poff et al. 1997).
Several studies have described the potential consequences of river regulation for riverine ecology in a ‘qualitative’ manner (e.g. Kingsford 2000; Nilsson and Berggren 2000; Bunn and Arthington 2002; Poff et al. 2009; Tockner et al. 2010). Attempts to quantify relationships between river regulation and biodiversity have, however, been so far incomplete or inconclusive (Poff and Zimmerman 2010). Therefore it is difficult to incorporate impacts of river regulation on biodiversity in strategic environmental assessments, which frustrates sound decision making (Arthington et al. 2006; Jackson et al. 2001; Stone 2008).
At a global level, human impacts on river systems are qualitatively assessed, using expert models or direct correlations between species distribution data and abiotic data (Sala et al. 2000; Xenopoulos et al. 2005). Quantitative assessments using empirically based relationships between flow alteration and ecological impact and state of the art models are necessary for more reliable impact assessments to support policy makers at both a regional and global level. Global models of environmental change and biodiversity, such as IMAGE (MNP 2006) and GLOBIO (Alkemade et al. 2009, 2011; Netherlands Environmental Assessment Agency 2010), are important tools for exploring policy options and scenarios for biodiversity conservation (e.g. within the framework of the Convention on Biological Diversity or the Ramsar Convention on Wetlands). The GLOBIO model comprises a module for terrestrial (Alkemade et al. 2009), and (recently) for inland freshwater ecosystems, referred to as GLOBIO-aquatic (Alkemade et al. 2011; Janse et al. unpublished manuscript). Considering the relatively high biodiversity value and associated conservation importance of floodplains, the effects of alteration of their natural flow regime need to be implemented in GLOBIO-aquatic.
The GLOBIO model is based on empirically based cause–effect relationships between environmental drivers and biodiversity impacts, formulated through meta-analyses of published data (Alkemade et al. 2009). The impact on biodiversity is expressed in terms of the mean species abundance (MSA) of original species, relative to their abundance in an undisturbed control situation (Alkemade et al. 2009; Secretariat of the Convention on Biological Diversity 2006), which is a measure of intactness of biodiversity similar to the Biodiversity Integrity Index (Majer and Beeston 1996) and the Biodiversity Intactness Index (Scholes and Biggs 2005).
This study contributes to the GLOBIO model by exploring the consequences of river regulation on the biodiversity intactness of flood-dependent wetlands. To uncover and quantify any general patterns at the regional and global level, we systematically searched the scientific literature for existing datasets relating community composition to hydrological change. We calculated the MSA to quantify effect size. As the MSA is not frequently used in meta-analyses and does not completely cover the complex biodiversity concept, we compared the MSA to a second indicator: the relative change of species richness (Faith et al. 2008). Meta-analyses were used to combine effect sizes across all studies and test the overall significance of flow regime alteration. Linear mixed effect models were used to explore potential effect modifiers.
Methods
Systematic search and data extraction
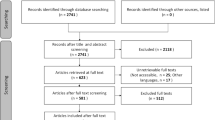
In February 2011, relevant published peer-reviewed articles on the impact of hydrological alterations on the biota in floodplain wetlands were searched using the electronic databases ISI Web of Knowledge and Scopus (Table 1). The setup of the systematic search is based on the guidelines provided by the Collaboration for Environmental Evidence ( 2013).
Studying the relationship between hydrology and ecology can typically be approached from either a hydrological or an ecological perspective. We used concepts from both disciplines for the search terms, and compiled search strings for extracting relevant articles from the databases as effectively as possible (Table 2). No grey literature was used in this systematic review. Possible bias resulting from this exclusion was verified.
Titles and abstracts of all returned hits were checked and judged against the aim of the review. From the papers that were not discarded we selected those that met the following criteria:
-
Relevant subject: Community composition in any floodplain wetland. Studies were included irrespective of habitat or spatial scale, except for experimental studies with a focus on micro scale processes.
-
Types of intervention: Human induced alteration of the natural flow regime, described in quantitative units.
-
Types of comparator: Undisturbed reference conditions in space or time (Before-After or Control-Impact studies).
-
Types of outcome: A quantitative comparison in terms of community composition (species richness, abundance or density).
The search query resulted in 686 unique articles, of which 28 fulfilled the selection criteria and contained usable data. 20 papers reported on community composition in terms of species abundance and 19 papers reported on local species richness (11 studies reported on both). All 28 articles reported on at least some differences in species composition between regulated (control) and unregulated (impact) sites. In several cases the differences were reported to be non-significant and were attributed to natural variation between sites. However, most studies showed differences that could be interpreted as effects of hydrological alterations, although not all changes had similar direction. The majority of the papers presenting data on species richness reported a reduction in species richness (16 out of 19). Studies on species abundance revealed a more complex picture and reported on shifts (positive and negative) rather than general decreases in the number of individuals. Table 3 shows which taxonomic groups were represented by the 28 papers, along with the types of hydrological regulation and the (terrestrial) biomes indigenous to where the studies took place (See Online Resource 1 for a short description of each study).
The available data on species richness and abundance, along with sample variance (if available) was extracted from the selected publications. Java Plot Digitizer 2.5.1 software was used to extract data from figures if values were not presented in tables. Abundance may have been recorded as density (number of individuals) or relative cover, depending on the taxonomic group studied. We preferred averaged data over cumulative numbers and, if presented, used transformed or rarified data to correct for differences in sampling size. To assess the hydrological alteration, we included the type and degree of hydrological alteration in the database by means of a short qualitative description and extraction of quantitative data of any hydrological variable provided. Data from disparate locations or different degrees of hydrological alteration presented in one article were treated as different datasets and considered independent. This resulted in 29 datasets for which we could calculate the MSA, and 32 datasets for which we could calculate the relative species richness. Additionally, we recorded system characteristics that could be regarded as cofactors, such as location (coordinates) and taxonomic details of the species studied.
Data synthesis and presentation
Ecological change
We used the MSA and the relative species richness as metrics of effect size for a quantitative comparison of datasets. To estimate the MSA of a dataset, we calculated the relative abundance of each species represented in the dataset (Eq. 1a). Effect sizes of all species were then averaged to calculate the Mean Species Abundance (Eq. 1b):
where R is is the ratio between the density of species i in the disturbed site d and its abundance in the undisturbed reference situation c in study s, for A isc >0. MSA s is the mean species abundance estimated in study s. Ns is the number of species in study s. If the abundance of a species in the disturbed site was higher than in the undisturbed floodplain, the ratios were truncated to 1. The variance of the MSA value for each study was estimated by calculating the variance of the external or the internal error (cf. Benítez-López et al. 2010; see Online Resource 2 for details). Taking a conservative approach, the larger of the two variances was used in the meta-analysis (DerSimonian and Laird 1986).
For each dataset on species richness, the relative species richness (LR) was calculated by dividing the number of species Jx found in the disturbed (impacted) sites by the number of species Jo found in undisturbed reference (control) sites. The logarithm of the ratio was used as effect size for its statistical properties (Hedges et al. 1999):
The variance of each individual effect size was calculated as:
where Jx and Jo are the species number and \(\sigma_{{J_{x} }}^{2}\) and \(\sigma_{{J_{o} }}^{2}\) are sample variances of the species number found in impacted sites (x) and undisturbed reference sites (o) respectively. In case Jx = 0 the data point was omitted.
Hydrological change
Ideally, the relationship between flow alteration and ecological variables would be expressed as a simple quantitative ratio (i.e. % ecological change in terms of % flow alteration) (Poff et al. 2009). However, the natural flow regime comprises numerous interacting components that are hypothesized to drive ecological processes. Moreover, measures of flow modification are mostly inconsistently reported in the literature (Olden and Poff 2003; Poff and Zimmerman 2010). Poff et al. (2009) remarked that ecological changes may also be formalised and empirically tested when they are expressed as categorical responses. In that vein, we decided to group hydrological alterations into three subclasses on an ordinal scale (low, medium, high), referred to as disturbance classes, to formalize the degree of alteration. If presented, formal indices of hydrological alteration were used to subdivide the cases over the three categories. Otherwise, the proportional changes of the primary hydrological variables that were presented were used for classification, with reference to the qualitative descriptions given by the authors (see Online Resource 2 for details).
Meta-analysis
For both metrics a random effect meta-analysis was used to derive a weighted mean for all datasets and test the global mean effect of flow regulation on the biodiversity of floodplains. Sampling variances were used to calculate weights and included in the analysis to give studies with relatively good precision more weight (Osenberg et al. 1999). Publication bias was assessed by inspecting funnel plots of asymmetry along with formal regression tests (Egger et al. 1997; Viechtbauer 2010), assuming that studies with large sample sizes would be near the average, and small studies (with more variance) would be spread on either side of the average. Heterogeneity was assessed by inspection of forest-plots and formal tests of heterogeneity Q and I2 (Borenstein et al. 2009). A significant Q (P < 0.05) denotes that the variance among effect sizes is greater than expected from sampling error and indicates that different studies do not share a common effect size. The I2 index quantifies the degree of heterogeneity and is a measure of the signal to noise ratio across the observed effect estimates (Huedo-Medina et al. 2006). To elaborate on the degree of hydrological alteration, separate meta-analyses were performed for each of the disturbance classes containing non-duplicated independent datasets. The meta-analyses were performed with the ‘metafor’ package available in R 2.15.1 software (Viechtbauer 2010).
To assess whether the relationship between hydrological alteration and ecological impact is influenced by other variables, we explored sources of heterogeneity by building linear mixed-effect (LME) models with several potential effect modifiers as fixed effects and the source article as a random effect. As well as the degree of hydrological alteration (disturbance classes), the factors considered were the percentage of non-natural land use in the catchment area, biome-type and taxon. We used the coordinates of each study site to retrieve the percentage of non-natural land use (NNLU) in the catchment area upstream of the study site from the IMAGE database (MNP, 2006). We subdivided the studies over three categories: low (<10 %), medium (10–50 %) and high (>50 %) percentage of NNLU. The IMAGE framework was also used to assign biome types to the studies according to their location (cf. Benítez-López et al. 2010). Furthermore, species identity was used to divide the datasets over different taxonomic groups: mammals, birds, fish, invertebrates and plants, as these groups are expected to present distinct differences in their response to alterations. Models were compared by means of Akaike’s information criterion (AIC) and Akaike weights.
Results
Meta-analysis
Our analysis showed that on average, alteration of the natural flow regime has a negative impact on biodiversity in floodplain wetlands. The meta-analysis of all MSA datasets results in a pooled effect size of 0.482 (Table 4)—a significant divergence from the no effect level (P < 0.0001). Publication bias was low (regression test, P > 0.5), and the funnel plots showed no asymmetry (forest plots and funnel plots are presented in Online Resource 3). The high values for Q and I2 indicated that the datasets did not share a common effect size (P(Q) < 0.0001; Table 4), and this was confirmed by inspection of the funnel plot, which showed large deviations in the effect size of datasets with small variances. The high fail-safe number indicated that a large number of studies reporting neutral effects would be needed to overturn the significance of the pooled effect size, and therefore the pooled effect size can be considered a reliable estimate of the mean effect.
When considering the datasets on species richness, the all-encompassing meta-analysis also showed a significant effect (P < 0.01; Table 5), with a pooled LR of −0.140 (i.e. a 28 % reduction). There was no evidence for publication bias and, in contrast to the MSA, there was no evidence for heterogeneity (P(Q) > 0.1; Table 5); the low values for Q and I2 indicated that the variance in effect size values was not higher than expected from natural variation and the studies might share a common ‘true’ effect size.
Subdividing datasets over three classes of hydrological alteration did not lead to significant differences between the classes (Welch’s t tests; P > 0.1), and the pooled effect sizes per disturbance class did not all differ significantly from the no-effect level, as was shown by subgroup meta-analyses (Tables 4, 5). However, the disturbance class representing the strongest disturbance did show the largest pooled effect size, while lesser disturbance led to smaller (non-significant) effects, which was consistent for the MSA (Fig. 1) and the LR (Fig. 2). Increase in the intensity of hydrological alteration tends to lead to more drastic deviation from the reference state.

Outcome of the meta-analysis; MSA values for the three disturbance classes ±1 SEM (dist. class ‘low’, n = 4; dist. class ‘medium’, n = 10; dist. class ‘high’, n = 15). MSA = 1 indicates ‘no effect’. Asterisks denote denotes a significant difference (P < 0.001) compared to the no-effect level

Outcome of the meta-analysis; average effect size of the log ratio of species richness for the three disturbance classes ±1 SEM (dist. class ‘low’, n = 4; dist. class ‘medium’, n = 10; dist. class ‘high’, n = 18). The single asterisk denotes a significant difference (P < 0.05) compared to the no-effect level
Heterogeneity
For both metrics the degree of hydrological alteration (i.e. classes of disturbance) was revealed to be a relevant factor in explaining variance among the effect sizes (Table 6). Examination of the linear mixed-effect (LME) models revealed, however, that the degree of disturbance is not the only relevant factor. In fact, the most comprehensive LME models turned out to be the most parsimonious in explaining heterogeneity i.e. they presented the lowest AIC values (Table 6). For the MSA, the smallest contributive factor is the percentage of NNLU in the catchment area, as deduced from comparison with the second best explanatory model, while for the LR the type of biome is the least additive.
Discussion
The qualitative review of the selected articles that resulted from the literature search had already revealed that 84 % of the articles reported a decrease in species, corroborating previous summaries by Poff and Zimmerman (2010), who also reported 84 % of studies that presented negative ecological responses, and Lloyd et al. (2003), who found that 86 % of the studies presented an negative ecological effect (the latter also included in-channel responses). Although such an overview makes it clear that flow modification does affect biodiversity in floodplain wetlands in most cases, the responses of individual studies are variable to some extent. Hence, such a summary does not permit a quantitative estimate of the average effect and its significance. Here we provide evidence showing that river regulation has a significant overall negative effect on biodiversity intactness in floodplain wetlands, and present a reliable quantitative estimate of the mean response: on average, alteration of the natural flow regime reduces the mean abundance of the original species (MSA) by >50 %, and species richness by >25 %.
It has until now been difficult to detect general, transferable quantitative relationships between individual measures of flow alteration and ecological response (Poff and Zimmerman 2010). This is partly due to the fact that there is a large variety of natural flooding regimes, which is also reflected in our database. Another important factor here is inconsistent reporting on the myriad hydrological variables in the literature, which frustrates possible identification of any relationships. In the search for generalities and greater quantitative understanding at the regional and global level, we circumvented these difficulties by deliberately including different flooding regimes, and categorizing the responses of hydrological variables (both qualitative and quantitative) into three broad categories to formalize the degree of hydrological alteration. Although this method allows for some arbitrariness and the resulting comparisons are only semi-quantitative, we argue that the method does provide some valuable information on the degree and effect of alteration and an opportunity to test it.
Subgroup analysis revealed a pattern indicating an effect of the degree of flow modification for both MSA and LR; the studies that were classified as lightly disturbed presented on average the least ecological response, whereas the studies assigned to the highest disturbance category showed on average the strongest ecological response. The importance of degree of flow modification in explaining heterogeneity was confirmed by the LME models.
If one compares indices of biodiversity impact for the low degree of hydrological alteration, it appears that species richness is more robust than MSA, indicating that the abundance of species is decreasing while species richness is not affected per se, revealing a complementarity between the two indicators. On longer timescales, however, reduced abundances can have serious consequences when only juveniles are impacted (Braatne et al. 2007) and community regeneration is severed. In general, (severe) flow modification showed a net decrease in species richness (including the arrival of new species), implying that the MSA does not provide an exact picture of the average decrease in abundance of any species as it must also be influenced by the disappearance of original species.
The LME models indicated that the relationship between flow modification and biodiversity impact is not determined solely by the degree of alteration to the river; other factors also influence the impact on biodiversity. One factor we considered was the percentage of non-natural land use (NNLU) in the catchment area. NNLU itself may impact the biodiversity of streams (e.g. Weijters et al. 2009) as land use covaries with riparian degradation (Walsh et al. 2007), the inflow of nutrients (Crosbie and Chow-Fraser 1999) and other (toxic) pollutants. Although the studies used in our analyses control for environmental conditions other than the hydrological treatment, the reference conditions do not always reflect pristine reference conditions, and vulnerable species may have already disappeared. Indeed, inspection of our data suggests that areas with a low percentage of NNLU are more sensitive to river regulation than areas with high NNLU. Furthermore, the relationship seems to be determined by the biome in which the wetland is located, and the type of taxa studied. Preliminary analysis of our data indicates that in terms of the MSA, animals tend to have a stronger response to flow modification than plants, while in terms of species richness plants appear more sensitive. Poff and Zimmerman (2010) also mentioned variation in response for different taxonomic groups, showing that fish are especially sensitive to flow alteration. Although animals are more mobile than plants, the latter may show greater adaptive phenotypic plasticity and benefit from the presence of a seed bank (Jansson et al. 2000). Nonetheless, the effect on plant species may simply be delayed if plant communities respond slowly to new environmental conditions and sudden shifts in dominance may appear at a later point in time (Braatne et al. 2007).
These factors (and probably many others) should therefore be taken into account when further investigating and quantifying the relationship between flow modification and biodiversity impact, particularly for the MSA for which heterogeneity was found to be high in the dataset. For species richness, however, there was no evidence for heterogeneity, indicating that the underlying ecological response is more robust and may be easier to predict in a field situation.
Generally, a paucity of data limits a more detailed analysis (Lloyd et al. 2003). There were already only a few data points available to test the effects of a low degree of hydrological alteration, indicating that there is a bias in the scientific literature towards studies reporting on severely disturbed flow regimes (Poff and Zimmerman 2010). Also, there was a geographical bias in our dataset, with most studies conducted in North America, Europe and Australia. Moreover, most studies concentrated on impacts on vegetation, with a disproportionate interest in cottonwood forest ecosystems in the American West. By expanding the search query, checking more electronic databases and including ‘grey’ literature, the number of usable datasets could possibly be increased, allowing greater quantification of the relationship between river regulation and biodiversity impact.
Despite these limitations, our quantitative meta-analysis reveals that flow modification has an overall significant negative effect on biodiversity intactness in floodplain wetlands, and provides an important first indication of the effect size in terms of mean species abundance and species richness. Moreover, our analyses suggest that impact on biodiversity is positively related to the degree of hydrological alteration. These insights may now be incorporated into the IMAGE–GLOBIO framework (Online Resource 4; Janse et al. unpublished manuscript), and used for policy decision making (Poff et al. 2003), so facilitating more effective and wise natural resource management (Stanford et al. 1996).
References
*References of studies included in the meta-analysis
Alkemade R, van Oorschot M, Miles L, Nellemann C, Bakkenes M, Ten Brink B (2009) GLOBIO3: a framework to investigate options for reducing global terrestrial biodiversity loss. Ecosystems 12:374–390
Alkemade R, Janse J, van Rooij W, Trisurat Y (2011) Applying GLOBIO at different geographical levels. In: Trisurat Y, Shrestha RP, Alkemade R (eds) Land use, climate change and biodiversity modelling: perspectives and applications. Information Science Reference, Hershey
Arthington AH, Bunn SE, Poff NLR, Naiman RJ (2006) The challenge of providing environmental flow rules to sustain river ecosystems. Ecol Appl 16:1311–1318
*Beauchamp VB, Stromberg JC (2008) Changes to herbaceous plant communities on a regulated desert river. River Res Appl 24:754–770
*Beauchamp VB, Stromberg JC, Stutz JC (2007) Flow regulation has minimal influence on mycorrhizal fungi of a semi-arid floodplain ecosystem despite changes in hydrology, soils, and vegetation. J Arid Environ 68:188–205
Benítez-López A, Alkemade R, Verweij PA (2010) The impacts of roads and other infrastructure on mammal and bird populations: a meta-analysis. Biol Conserv 143:1307–1316
Borenstein M, Hedges LV, Higgins JPT, Rothstein HR (2009) Introduction to meta-analysis. Wiley & Sons, New York
Braatne JH, Jamieson R, Gill KM, Rood SB (2007) Instream flows and the decline of riparian cottonwoods along the Yakima river, Washington, USA. River Res Appl 23:247–267
Bunn SE, Arthington AH (2002) Basic principles and ecological consequences of altered flow regimes for aquatic biodiversity. Environ Manag 30:492–507
*Chauhan M, Gopal B (2005) Vegetation structure and dynamics of a floodplain wetland along a subtropical regulated river. River Res Appl 21:513–534
Collaboration for Environmental Evidence (2013) Guidelines for systematic review and evidence synthesis in environmental management. Version 4.2. Environmental Evidence. www.environmentalevidence.org/Documents/Guidelines/Guidelines4.2.pdf
Crosbie B, Chow-Fraser P (1999) Percentage land use in the watershed determines the water and sediment quality of 22 marshes in the Great Lakes basin. Can J Fish Aquat Sci 56:1781–1791
Davidson TA, Mackay AW, Wolski P, Mazebedi R, Murray-Hudson M, Todd M (2012) Seasonal and spatial hydrological variability drives aquatic biodiversity in a flood-pulsed, sub-tropical wetland. Freshw Biol 57:1253–1265
DerSimonian R, Laird N (1986) Meta-analysis in clinical trials. Control Clin Trials 7:177–188
Dynesius M, Nilsson D (1994) Fragmentation and flow regulation of river systems in the northern third of the world. Science 266(6186):753–762
Egger M, Smith GD, Schneider M, Minder C (1997) Bias in meta-analysis detected by a simple, graphical test. BMJ 315:629
*Ellis LM, Crawford CS, Molles MC Jr (2001) Influence of annual flooding on terrestrial arthropod assemblages of a Rio Grande riparian forest. Regul Rivers 17:1–20
Faith DP, Ferrier S, Williams KJ (2008) Getting biodiversity intactness indices right: ensuring that ‘biodiversity’ reflects ‘diversity’. Glob Change Biol 14:207–217
*Falck MJ, Wilson KR, Andersen DC (2003) Small mammals within riparian habitats of a regulated and unregulated arid land river. West North Am Nat 63:35–42
*Franklin SB, Kupfer JA, Pezeshki SR, Gentry R, Smith RD (2009) Complex effects of channelization and levee construction on western Tennessee floodplain forest function. Wetlands 29:451–464
*Gergel SE, Dixon MD, Turner MG (2002) Consequences of human-altered floods: levees, floods, and floodplain forests along the Wisconsin river. Ecol Appl 12:1755–1770
*Gubiani ÉA, Gomes LC, Agostinho AA, Okada EK (2007) Persistence of fish populations in the upper Paraná river: effects of water regulation by dams. Ecol Freshw Fish 16:191–197
Hedges LV, Gurevitch J, Curtis PS (1999) The meta-analysis of response ratios in experimental ecology. Ecology 80:1150–1156
Huedo-Medina TB, Sanchez-Meca J, Marin-Martinez F, Botella J (2006) Assessing heterogeneity in meta-analysis: Q statistic or I2 index? Psychol Methods 11(2):193–206
*Hyslop EJ (1988) A comparison of the composition of the juvenile fish catch from the Sokoto-Rima floodplain, Nigeria in years preceding and immediately after upstream dam completion. J Fish Biol 32:895–899
Jackson RB, Carpenter SR, Dahm CN, McKnight DM, Naiman RJ, Postel SL et al (2001) Water in a changing world. Ecol Appl 11:1027–1045
*Jansson R, Nilsson C, Dynesius M, Andersson E (2000) Effects of river regulation on river-margin vegetation: a comparison of eight boreal rivers. Ecol Appl 10:203–224
*Jenkins KM, Boulton AJ (2007) Detecting impacts and setting restoration targets in arid-zone rivers: aquatic micro-invertebrate responses to reduced floodplain inundation. J Appl Ecol 44:823–832
Junk WJ, Bayley PB, Sparks RE (1989) The flood pulse concept in river-floodplain systems. Canadian special publication of fisheries and aquatic sciences 106:110–127
*Katz GL, Friedman JM, Beatty SW (2005) Delayed effects of flood control on a flood-dependent riparian forest. Ecol Appl 15:1019–1035
Kingsford RT (2000) Ecological impacts of dams, water diversions and river management on floodplain wetlands in Australia. Austral Ecol 25:109–127
*Kingsford RT, Jenkins KM, Porter JL (2004) Imposed hydrological stability on lakes in arid Australia and effects on waterbirds. Ecology 85:2478–2492
Ladson AR, White LJ, Doolan JA, Finlayson BL, Hart BT, Lake PS et al (1999) Development and testing of an index of stream condition for waterway management in Australia. Freshw Biol 41:453–468
Lloyd N, Quinn G, Thoms M, Arthington A, Gawne B, Humphries P et al (2003) Does flow modification cause geomorphological and ecological response in rivers? A literature review from an Australian perspective. CRC for Freshwater Ecology, Technical report, Canberra
Loh J, Wackernagel M (eds) (2004) Living planet report 2004. WWF, Gland
*Lovell JT, Gibson J, Heschel MS (2009) Disturbance regime mediates riparian forest dynamics and physiological performance, Arkansas river, CO. Am Midl Nat 162:289–304
Majer JD, Beeston G (1996) The biodiversity integrity index: an illustration using ants in Western Australia. Conserv Biol 10(1):65–73
*Mallik AU, Richardson JS (2009) Riparian vegetation change in upstream and downstream reaches of three temperate rivers dammed for hydroelectric generation in British Columbia, Canada. Ecol Eng 35:810–819
MEA (2005) Ecosystems and human well-being: current state and trends. World Resources Institute, Washington
*Merritt DM, Poff NLR (2010) Shifting dominance of riparian Populus and Tamarix along gradients of flow alteration in western North American rivers. Ecol Appl 20:135–152
Middleton BA (2002) The flood pulse concept in wetland restoration. Flood pulsing in wetlands: restoring the natural hydrological balance. Wiley, New York
MNP (2006) In: Bouwman AF, Kram T, Goldewijk KK (eds) Integrated modelling of global environmental change: an overview of image 2.4. Netherlands Environmental Assessment Agency (MNP), Bilthoven
Netherlands Environmental Assessment Agency (2010) Rethinking global biodiversity strategies: exploring structural changes in production and consumption to reduce biodiversity loss. Netherlands Environmental Assessment Agency, Bilthoven
Nilsson C, Berggren K (2000) Alterations of riparian ecosystems caused by river regulation. Bioscience 50:783–792
*Nilsson C, Ekblad A, Gardfjell M, Carlberg B (1991) Long-term effects of river regulation on river margin vegetation. J Appl Ecol 28:963–987
*Nilsson C, Jansson R (1995) Floristic differences between riparian corridors of regulated and free-flowing boreal rivers. Regul Rivers 11:55–66
Nilsson C, Svedmark M (2002) Basic principles and ecological consequences of changing water regimes: riparian plant communities. Environ Manag 30:468–480
Olden JD, Poff NLR (2003) Redundancy and the choice of hydrologic indices for characterizing streamflow regimes. River Res Appl 19:101–121
Osenberg CW, Sarnelle O, Cooper SD, Holt RD (1999) Resolving ecological questions through meta-analysis: goals, metrics and models. Ecology 80(4):1105–1117
*Paetzold A, Yoshimura C, Tockner K (2008) Riparian arthropod responses to flow regulation and river channelization. J Appl Ecol 45:894–903
Poff NL, Richter BD, Arthington AH, Bunn SE, Naiman RJ, Kendy E et al (2009) The ecological limits of hydrologic alteration (ELOHA): a new framework for developing regional environmental flow standards. Freshw Biol 55:147–170
Poff NL, Zimmerman JKH (2010) Ecological responses to altered flow regimes: a literature review to inform the science and management of environmental flows. Freshw Biol 55:194–205
Poff NLR, Allan JD, Bain MB, Karr JR, Prestegaard KL, Richter BD et al (1997) The natural flow regime. Bioscience 47:769–784
Poff NLR, Allan JD, Palmer MA, Hart DD, Richter BD, Arthington AH et al (2003) River flows and water wars: emerging science for environmental decision making. Front Ecol Environ 1:298–306
Poff NLR, Olden JD, Merritt DM, Pepin DM (2007) Homogenization of regional river dynamics by dams and global biodiversity implications. Proc Natl Acad Sci 104:5732
Postel S, Richter BD (2003) Rivers for life: managing water for people and nature. Island Press, Washington
*Quinn CH, Ndangalasi HJ, Gerstle J, Lovett JC (2005) Effect of the lower Kihansi hydropower project and post-project mitigation measures on wetland vegetation in Kihansi Gorge, Tanzania. Biodivers Conserv 14:297–308
Revenga C, Brunner J, Henninger N, Kassem K, Payne R (2000) Pilot analysis of global ecosystems: freshwater systems. World Resources Institute, Washington
Revenga C, Campbell I, Abell R, De Villiers P, Bryer M (2005) Prospects for monitoring freshwater ecosystems towards the 2010 targets. Philos Trans R Soc B 360:397
*Robertson AI, Bacon P, Heagney G (2001) The responses of floodplain primary production to flood frequency and timing. J Appl Ecol 38:126–136
Sala OE, Chapin FS, Armesto JJ, Berlow E, Bloomfield J, Dirzo R et al (2000) Global biodiversity scenarios for the year 2100. Science 287:1770–1774
Scholes RJ, Biggs R (2005) A biodiversity intactness index. Nature 434:45–49
Secretariat of the Convention on Biological Diversity (2006) Global biodiversity outlook 2. Secretariat of the Convention on Biological Diversity, Montreal
*Shafroth PB, Stromberg JC, Patten DT (2002) Riparian vegetation response to altered disturbance and stress regimes. Ecol Appl 12:107–123
Stanford JA, Ward JV, Liss WJ, Frissell CA, Williams RN, Lichatowich JA et al (1996) A general protocol for restoration of regulated rivers. Regul Rivers 12:391–413
Stone R (2008) Three gorges dam: into the unknown. Science 321:628
*Stromberg JC, Beauchamp VB, Dixon MD, Lite SJ, Paradzick C (2007a) Importance of low-flow and high-flow characteristics to restoration of riparian vegetation along rivers in arid south-western united states. Freshw Biol 52:651
*Stromberg JC, Lite SJ, Marler R, Paradzick C, Shafroth PB, Shorrock D et al (2007b) Altered stream-flow regimes and invasive plant species: the Tamarix case. Glob Ecol Biogeogr 16:381–393
Tockner K, Pusch M, Borchardt D, Lorang MS (2010) Multiple stressors in coupled river–floodplain ecosystems. Freshw Biol 55:135–151
Tockner K, Stanford JA (2002) Riverine flood plains: present state and future trends. Environ Conserv 29:308–330
*Toner J, Farrell JM, Mead JV (2010) Muskrat abundance responses to water level regulation within freshwater coastal wetlands. Wetlands 30:211–219
*Uowolo AL, Binkley D, Carol Adair E (2005) Plant diversity in riparian forests in northwest Colorado: effects of time and river regulation. For Ecol Manag 218:107–114
*Van Geest GJ, Coops H, Roijackers RMM, Buijse AD, Scheffer M (2005) Succession of aquatic vegetation driven by reduced water-level fluctuations in floodplain lakes. J Appl Ecol 42:251–260
*Vaňhara J (1991) A floodplain forest mosquito community after man-made moisture changes (culicidae, diptera). Regul Rivers 6:341–348
Viechtbauer W (2010) Conducting meta-analyses in R with the metafor package. J Stat Softw 36(3):1–48
Walsh CJ, Waller KA, Gehling J, Mac Nally R (2007) Riverine invertebrate assemblages are degraded more by catchment urbanization than by riparian deforestation. Freshw Biol 52:574–587
Ward JV, Tockner K, Schiemer F (1999) Biodiversity of floodplain river ecosystems: ecotones and connectivity. Regul Rivers 15:125–139
Ward JV, Tockner K, Arscott DB, Claret C (2002) Riverine landscape diversity. Freshw Biol 47:517–539
Weijters MJ, Janse JH, Alkemade R, Verhoeven JTA (2009) Quantifying the effect of catchment land-use and water nutrient concentrations on freshwater river and stream biodiversity. Aquat Conserv 19:104–112
World Wildlife Fund (2004) Rivers at risk: dams and the future of freshwater ecosystems. Report. WWF, Gland
Xenopoulos MA, Lodge DM, Alcamo J, Märker M, Schulze K, Van Vuuren DP (2005) Scenarios of freshwater fish extinctions from climate change and water withdrawal. Glob Change Biol 11:1–8
Acknowledgments
We would like to thank Michiel Bakkenes, Michiel Jeuken, Richard Peters, Nynke Schulp, Richard Verweij and Maria Witmer for their technical support and helpful comments during the development of this paper. We also would like to thank Liz Barnes and two anonymous reviewers for proofreading.
Author information
Authors and Affiliations
Corresponding author
Electronic supplementary material
Below is the link to the electronic supplementary material.
Rights and permissions
About this article

Cite this article
Kuiper, J.J., Janse, J.H., Teurlincx, S. et al. The impact of river regulation on the biodiversity intactness of floodplain wetlands. Wetlands Ecol Manage 22, 647–658 (2014). https://doi.org/10.1007/s11273-014-9360-8
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11273-014-9360-8