
Abstract
Proper management techniques on moist-soil wetlands provide methods for enhancement of established wetlands, restoration of former wetlands, and creation of new wetland habitat. These techniques also create suitable wetland habitat for non-breeding waterfowl and other wetland dependent species during winter. To understand moist-soil managed wetland vegetative patterns, aspects such as plant species distribution, reproductive strategy, seed bank composition and viability should be thoroughly characterized. We investigated soil seed bank potential of moist-soil managed wetlands on Richland Creek Wildlife Management Area, Texas to determine which treatment (i.e., drawdown or flooded) produced the most desirable moist-soil plants. A total of 27 species germinated, producing 3,731 and 3,031 seedlings in drawdown and flooded treatments, respectively. There were also differences in stem densities between treatments of desirable and non-desirable species. Drawdown treatments had more seedlings germinate than flooded treatments, validating the notion that drawdown treatments provide favorable conditions for seed germination. Drawdown and flooding techniques, when properly timed, will allow managers to drive and directly influence managed wetland plant communities based on seed bank composition and response to presence or absence of water during the germination period.
Similar content being viewed by others
Avoid common mistakes on your manuscript.
Introduction
Moist-soil managed wetlands are typically shallow water areas impounded by levees, allowing for flooding (i.e., water addition) during fall and winter, and drawdown (i.e., water removal) in spring and summer (Fredrickson and Taylor 1982; Smith et al. 1989; Haukos and Smith 1993; Lane and Jensen 1999; Anderson and Smith 2000; Strader and Stinson 2005). Plant species recruitment typically occurs during drawdown periods, when the moist-soil managed wetland is free of standing water, but the substratum remains moist (Lane and Jensen 1999). During drawdown, persistent seeds within the seed bank will germinate in response to favorable conditions such as varying temperature, light, and oxygen regimes (Leck 2003). Short duration drawdowns promote germination and growth of annual wetland plants that produce high quality seeds, tubers, and structure for aquatic invertebrates, waterfowl, shorebirds, and other waterbirds (Lane and Jensen 1999). Conversely, extended periods of flooding promotes lower quality seed producing perennial aquatic and emergent wetland plants (Howard and Mendelssohn 1995), but provide important foraging habitat, substrate, and cover for migrating and wintering waterfowl and other wetland dependent species (Fredrickson and Taylor 1982). By manipulating water levels within moist-soil managed wetlands, managers can target specific plant community development, seed germination, growth and subsequent seed production, and promote desirable moist-soil plants (van der Valk and Davis 1978).
Generally, the overarching objectives of moist-soil management are to increase wetland productivity and waterfowl use, where moist-soil management techniques maximize production of naturally occurring wetland plants (Strader and Stinson 2005). Moist-soil management promotes production of naturally occurring desirable wetland vegetation by emulating and manipulating natural wetland functions (e.g., hydrology and successional stage) (Lane and Jensen 1999). Hydrology is a dominant factor controlling development of spatial variation in wetland plant communities and is responsible for horizontal zonation of adult plants, seeds, and seedlings in both natural and moist-soil managed wetlands (Baldwin et al. 2001). Manipulations in hydrology will influence plant species composition during patterns of emergence from the seed bank (Casanova and Brock 2000; Baldwin et al. 2001). For example, van der Valk and Davis (1978) established relationships between periodic drawdown, emergence from the seed bank, vegetative growth, and inundation in prairie pothole wetlands which share hydrology regimes similar to moist-soil managed wetlands for waterfowl throughout the southern US (Strader and Stinson 2005). Annual plants are important components of both wetland types and their presence is due to favorable drawdown and soil exposure conditions for seed germination and seedling growth (Leck and Simpson 1993). Conversely, flooding can reduce seed germination and severely reduce seedling survival (Galinato and van der Valk 1986; Baldwin et al. 1996). Consequently, water manipulation is important in controlling temporal variation in plant species composition of moist-soil managed wetlands (Baldwin et al. 2001) and to maximize habitat availability and utilization with depth and timing of flooding and/or drawdown should be well planned (Lane and Jensen 1999).
To maintain, promote, or change moist-soil plant populations, production, and floristic diversity, aspects such as species distribution, reproductive strategy, seed bank composition and viability should be known and quantified for a given managed wetland (Leck and Graveline 1979). As such, examining seed bank potential can help managers (1) maintain a persistent and desirable plant community and (2) understand temporal seed bank dynamics, particularly as related to providing quality habitat to migrating and wintering waterfowl (Fredrickson and Taylor 1982). Such research on wetland seed banks is significant to plant population ecology, as well as many applied fields, such as conservation, restoration, and success of managed wetland communities (Araki and Washitani 2000; Middleton 2003; Bossuyt and Honnay 2008).
When moist-soil managed wetlands are specifically created to provide wintering waterfowl habitat, seed bank dynamics can ultimately influence success and failure of any management objectives, even under proper water manipulation strategies. Successful seed bank development requires a basic understanding of existing seed bank composition (van der Valk et al. 1992). Specifically, temporal changes in seed bank size, species composition, and persistence will provide insight into the importance of the seed bank to the overall management objectives (Leck 2003). Indeed, in newly created wetlands, existing seed banks may greatly influence initial plant species composition, where non-desirable plant communities may be enhanced or promoted depending upon hydrology and/or basin manipulations (Galinato and van der Valk 1986; Baldwin et al. 1996; Leck 2003).
Commonly recognized as a primary limiting physical factor that varies along elevation gradients in many wetland habitats, water depth will have negative impacts on moist-soil plant survival, at both long and short temporal scales (Howard and Mendelssohn 1995). Emergent herbaceous moist-soil species vary in response to flooding or submergence, which is generally regarded as inhibitory to plant growth (Howard and Mendelssohn 1995; Flynn et al. 1999; Casanova and Brock 2000; Nicol et al. 2003; Capon 2007). Flooding and/or submergence typically promotes the growth of non-desirable wetland plant species (Fredrickson and Taylor 1982; Lane and Jensen 1999). In such conditions, subsequent management efforts may be hindered, particularly if non-desirable plants are not controlled or effectively removed from the seed bank (e.g., interrupt desirable seed production). The primary objective of this portion of the research was to quantify seed bank expression of managed and unmanaged moist-soil wetlands at the Richland Creek Wildlife Management Area (RCWMA) in east-central Texas. Specifically, the effects of experimentally simulated drawdown and flooded conditions on seed bank expression were examined over time for moist-soil managed wetlands at the RCWMA.
Study area
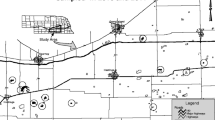
Research was conducted on the RCWMAs North Unit moist-soil managed wetland cells 1–4 and unmanaged moist-soil wetland cells named the Triangle, Gut, and DU marsh (Fig. 1). The RCWMA (31°13′N, 96°11′W) is located 40 km southeast of Corsicana in Freestone and Navarro Counties, Texas, along US highway 287 and FM 488 between Richland-Chambers Reservoir and the Trinity River encompassing 6,271 ha in the ecotone separating the Post Oak Savannah and Blackland Prairie ecological regions (TPWD Texas Parks and Wildlife Department 2005). Local climate is subtropical with mild winters and warm humid summers, an average daily summer temperature of 34 °C and winter temperature of 5 °C, a growing season of 246 days, and average rainfall of 101.6 cm per year (Natural Resource Conservation Service, NRCS 2002). Rainfall is typically distributed evenly throughout the year. Soils on the area are predominately of the Trinity series, which are fine, montmorillonitic, thermic, very haplaquolls, and mollisol soils (NRCS 2002).

Locations of wetlands used to collect seed bank samples on Richland Creek Wildlife Management Area, Freestone County, Texas 2005
The North Unit contains the moist-soil managed and unmanaged wetland cells, which are large non-forested areas characterized by a diverse herbaceous community. The typical water management within the managed moist-soil wetlands is a strategy that consists of slow drawdown (i.e., removal of water) starting late March–early April and lasting until mid-August. Inundation (i.e., flooding) begins in late August and lasts throughout fall and winter until drawdown the following spring. Water management within the unmanaged moist-soil wetlands is through over bank river flooding, precipitation events, and other passive water delivery. These actions produce common species such as barnyard grass (Echinochloa crusgalli), erect burhead (Echinodorus spp.), delta duck potato (Sagittaria spp.), square-stem spike rush (Eleocharis quadrangulata), wild millet (Echinochloa walteri), and water primrose (Ludwigia peploides).
Methods
Seed bank sample collection
Seed bank samples were collected from four managed moist-soil wetland(s) cell 1 (n = 17), cell 2 (n = 21), cell 3 (n = 25), and cell 4 (n = 12) as well as from three unmanaged moist-soil wetlands named the Triangle (n = 15), Gut (n = 15), and DU marsh (n = 15), respectively, a week prior to, or during, initial drawdown during late March 2005. The number of samples collected in the four managed moist-soil wetlands was determined by the number of established permanent plots (Collins 2012). While the three older moist-soil managed wetlands did not have established plots the number of samples collected was consistent among these three managed wetlands. Permanent plots were established using line transects within the four newly created moist-soil managed wetland that were systematically located lengthwise running east–west within each wetland. Facing west on the middle transect in each moist-soil managed wetland every 50 m a two-digit number was removed from a random number generator. The number determined how many paces were walked in the approximate cardinal direction (i.e., north or south) off the middle transect (ex. 42 = 42 paces). If the number was odd, the plot was placed to the south the appropriate number of paces, and if the number was even, the plot was placed to the north of the transect the appropriate number of paces. Once at the established plot location seed bank sample collection occurred in the approximate southeastern corner of all plots.
Seed bank samples were collected using a manual soil corer (10 cm long × 5.5 cm diameter), following Kadlec and Smith (1984) and Haukos and Smith (2001). Once removed, all samples were placed into labeled plastic bags on dry ice, and stored in a dark walk-in refrigerator (4 °C) where samples remained for ≤3 weeks before they were deployed in seed bank expression experiments. While this temperature could have an adverse effect on germination of seedlings it is consistent with procedure followed in other relevant seed bank research (Galinato and van der Valk 1986; Moore and Keedy 1988; Shipley et al. 1989; Shipley and Parent 1991; Budlesky and Galatowitsch Budelsky and Galatowitsch 1999; Boedeltje et al. 2002; Gurnell et al. 2006; Li et al. 2008; Hopfensperger et al. 2009).
We followed Kadlec and Smith (1984) and Haukos and Smith (2001) who used the direct application seedling emergence method for our germination experiments. We used the direct application seedling emergence method rather than the bulk reduction method (ter Heerdt et al. 1996) or substrate saturation method (Boedeltje et al. 2002) because the ease of experimental setup, appropriateness for large-scale studies (McFarland and Shafer 2011) and was considered suitable for determining the composition of the germinable seed bank and detecting temporal trends in germination (Capon and Brock 2006). While this method does have its disadvantages it tended to corroborate field level vegetative data being collected simultaneously (Collins 2012).
Seed bank expression experiments
Individual seed bank samples were homogenized, divided in half, and each half placed into an individual 4 × 10 × 20 cm plastic dish lined with 2 cm of sterilized potting soil. Each dish was uniquely labeled using a wooden tongue depressor. Each half of each seed bank sample was randomly assigned into a simulated drawdown or flooding treatment (van der Valk and Davis 1978; Kadlec and Smith 1984). Samples, in dishes, were then randomly arranged on four germination tables in the greenhouse at Stephen F. Austin State University (SFASU). To maintain similar environmental conditions on both sample halves (i.e., drawdown and flooding) samples were randomly placed on the same table, but no two half-samples were positioned next to one another. Dishes exposed to simulated drawdown treatments were monitored daily, and watered as needed with distilled water to maintain moist-soil conditions without standing water (Kadlec and Smith 1984; van der Valk and Davis 1978). Dishes exposed to the simulated flooding treatments were also monitored daily, and watered as needed to maintain ≥3 cm of standing water (van der Valk and Davis 1978; Fredrickson and Taylor 1982; Kadlec and Smith 1984; Casanova and Brock 2000; Haukos and Smith 2001; Capon 2007; Hopfensperger et al. 2009). Fredrickson and Taylor (1982) reported that in managed moist-soil wetlands that plants should attain a height of 10–15 cm before wetlands are reflooded as complete submergence of recently germinated annual moist-soil plants for longer than 2–3 days can retard their growth, therefore a depth of 4 cm was maintained in our flooded treatment. Dishes were monitored from 25 April to 31 October 2005, corresponding with the growing season in Navarro and Freestone Counties (NRCS 2002).
Soil seed bank assessment followed the seedling emergence technique (Smith and Kadlec 1983; Pederson and Smith 1988; Haukos and Smith 2001), whereas germinated seedlings were identified and counted once monthly generating seedling density (seedlings/m2). Once identified, seedlings were carefully removed to prevent soil disturbance. Unidentified seedlings were transplanted to individual containers and grown until identified. Nomenclature followed Correll and Johnston (1979) and seedlings were verified by voucher specimens at the SFASU Herbarium.
Seedling classification
Seedlings were classified as desirable or non-desirable, respectively, based upon their known value for waterfowl, following Fredrickson and Taylor (1982). Desirable plants were defined as those that provide energy or some other nutritive requirement to migrating and wintering waterfowl (Fredrickson and Taylor 1982; Stutzenbaker 1999; Strader and Stinson 2005). Non-desirable species were defined as those that provide neither high quantity nor high quality seed, and tend to dominate later successional stages (Fredrickson and Taylor 1982; Strader and Stinson 2005). Non-desirable species may, however, provide aquatic invertebrate substrate(s), or perform some other wetland functions (Fredrickson and Taylor 1982) and may not necessarily be non-desirable wetland plants, but are not considered direct food or food producers for wintering waterfowl (see Fredrickson and Taylor 1982).
After classification as desirable or non-desirable, seedlings were assigned to plant groups commonly used by the NRCS within the National Plant Database (i.e., annual introduced grass [aig], perennial native forb [pnf], annual native grass [ang], etc.) (USDA 2011). The following group assignments were used to indicate a combination of growth habit (grass, forb, shrub, vine, or grass-like), life cycle (annual or perennial), and source (native or introduced): ang, aig, perennial introduced forb, annual native forb, annual perennial native, perennial native, perennial native grass-like, pnf, annual perennial native subshrub, and annual native vine (USDA 2011). When both annual and perennial are indicated for one species, this indicates that the individual plant species can have growth durations as either annual or perennial.
Data analyses
Each dish was considered an experimental unit (Smith and Kadlec 1983). A suite of diversity indices (i.e., niche overlap, Simpson’s diversity index, Shannon–Wiener diversity index, species evenness) were calculated for both treatments (i.e., drawdown or flooded), moist-soil managed wetlands (i.e., specific managed wetland from which seed bank samples were removed), treatment × moist-soil managed wetlands, and treatment over time (i.e., 30-day periods). Percent similarity (i.e., niche overlap) was calculated using the relative abundance of all species summed to 100 %. This index is calculated by:
where P is the percentage similarity between sample 1 and 2, p 1i is the percentage of species i in community sample 1, p 2i is the percentage of species i in community sample 2.
This index ranges from 0 (no similarity) to 100 (complete similarity) allowing for comparison between units of interest (i.e., treatments, managed wetland, etc.) (Krebs 1999).
χ 2 analysis was used to examine differences in stem density (i.e., seedlings/m2) among (1) desirable and non-desirable moist-soil plants between simulated treatments, (2) desirable and non-desirable moist-soil plants over time (i.e., 30-day increments), (3) treatments among plant groups, (4) simulated treatments among managed and unmanaged moist-soil wetlands, (5) managed/unmanaged moist-soil wetlands among desirable/non-desirable plant species, (6) managed/unmanaged moist-soil wetlands between time periods, and (7) managed/unmanaged moist-soil wetlands between desirable/non-desirable plant species and time period. A repeated measure, three-way multivariate analysis of variance (MANOVA) was also used to examine differences in stem density between desirable and non-desirable plant species, among time periods and simulated treatments; between simulated treatments and species groups, moist-soil managed wetlands, and managed to unmanaged moist-soil wetlands between treatments as well as time period × desirable/non-desirable plant species, desirable/non-desirable plant species, and time periods. If differences (P < 0.05) occurred in MANOVA, subsequent univariate analysis of variance (ANOVA) were used, followed by least square mean separation if differences (P < 0.05) occurred in ANOVA.
Results
A total of 6,802 seedlings (\( \bar{x} \) = 28.3 seedlings/m2) representing 27 species were identified. Seedlings represented 14 families, 13 plant groups, and 6 standardized plant groups. Of the 27 species identified, only one (marsh flatsedge: Cyperus pseudovegetus) was not recorded during field transects (see Collins 2012). A total of 11 desirable (n = 5,127 individuals; \( \bar{x} \) = 21.4 seedlings/m2) and 16 non-desirable species (n = 1,675 individuals; \( \bar{x} \) = 6.9 seedlings/m2) were identified (Table 1). Approximately 75 % of all individual seedlings were desirable, regardless of experimental moist-soil treatment (i.e., drawdown or flooded; Table 1). Within experimental drawdown treatments, most germination occurred within the first 60 days, while germination within flooded treatments was more evenly distributed among the four 30-day time periods. Greater than 80 % of desirable plant species germinated within the first 60 days of experimental drawdown conditions. The first two 30-day time periods were dominated by least >50 % desirable moist-soil plant species germination, whereas the final two 30-day time periods were dominated by >50 % non-desirable moist-soil plant species.
Overall, Simpson’s and Shannon–Wiener species diversity indices were similar between experimental drawdown and flooded treatments, which ranged from 2.01 to 5.14 for drawdown treatment and 1.18–4.38 for flooded treatment (Table 2). Over time (0–30, 31–60, 61–90, 91–120 days) both diversity indices ranged between 1.18 and 5.14, respectively (Table 2). There was relatively high species similarity (32.7 %) between experimental drawdown and flooded treatments. Niche overlap estimates were comparable to the similarity estimates, as 39 % of species in drawdown treatments were also found in flooded treatments, and 42 % of species identified in flooded treatments were found in drawdown treatments. However, plant species evenness was skewed towards two desirable moist-soil plant species. Red-rooted flatnut sedge (Cyperus erythrorhizos) accounted for 36 % of all individual seedlings and 48 % of all desirable plant seedlings. Similarly, toothcup (Ammannia coccinea) accounted for 24 % of all individual seedlings and 31 % of all desirable plant seedlings. Although erect burhead and water primrose only accounted for 8 % of all individual seedlings, they accounted for 34 and 33 % of all non-desirable seedlings, respectively.
A total of 2,342 (\( \bar{x} \) = 31.2 seedlings/m2) and 1,114 (\( \bar{x} \) = 14.8 seedlings/m2) desirable seedlings germinated from seed bank samples collected in the managed moist-soil wetlands (cells 1–4) exposed to simulated drawdown and flooded treatments, respectively, and 305 (\( \bar{x} \) = 4.1 seedlings/m2) and 643 (\( \bar{x} \) = 8.6 seedlings/m2) non-desirable seedlings were identified from the same wetlands exposed to simulated drawdown and flooded treatments, respectively (Table 3). In the three unmanaged moist-soil wetland cells (i.e., Triangle, Gut, and DU marsh) a total of 780 (\( \bar{x} \) = 17.3 seedlings/m2) and 890 (\( \bar{x} \) = 19.7 seedlings/m2) desirable seedlings germinated from seed bank samples exposed to simulated drawdown and flooded treatments, respectively (Table 3), while a total of 304 (\( \bar{x} \) = 6.7 seedlings/m2) and 424 (\( \bar{x} \) = 9.4 seedlings/m2) non-desirable seedlings germinated from seed bank samples exposed to simulated drawdown and flooded treatments, respectively (Table 3). Desirable red-rooted flatnut sedge and nodding smartweed (P. lapathifolium) germinated at the greatest numbers under simulated moist conditions, while desirable toothcup germinated at the greatest numbers under simulated flooded treatment conditions. Non-desirable water primrose, frog fruit (Phyla lancelota), and waterhemp (Amaranthus tuberculatus) germinated at the greatest number under simulated drawdown conditions, while erect burhead, water primrose, and duck potato (S. lancifolia) germinated in the greatest numbers under flooded treatment conditions.
Stem densities varied between treatments and desirable and non-desirable moist-soil plants (χ 2 = 2,271.5, P < 0.001), where desirable plant species had greater stem densities in both simulated moist and flooded treatment. Stem densities also varied between desirable and non-desirable moist-soil plants across time periods (χ 2 = 544.6, P < 0.001), where desirable plant species had greater densities than during the first three time periods. Similarly, stem densities varied between treatments and moist-soil plant groups (χ 2 = 1,876.5, P = <0.001), where stem densities were typically greater in the simulated moist treatment. Stem densities varied (χ 2 = 731.9, P < 0.001) among individual moist-soil managed wetlands, but both desirable and non-desirable stem densities were greater in the simulated drawdown treatment. Stem densities varied between simulated treatments and among managed and unmanaged moist-soil wetland cells (χ 2 = 342.7, P < 0.001), where greatest densities occurred in managed wetlands, regardless of treatment. Desirable seedlings reached greater densities in both managed and unmanaged moist-soil wetlands (χ 2 = 278.5, P < 0.001). Stem densities for all seedlings were greatest from seed bank samples collected from managed wetlands during all four 30-day temporal periods (χ 2 = 137.4, P < 0.001). Finally, seedling stem densities were greatest for both desirable and non-desirable species in managed wetlands during all four 30-day temporal periods (χ 2 = 1,136.60, P < 0.001) and (χ 2 = 251.58, P < 0.001).
Stem density for all species combined did not vary between desirable and non-desirable species (Wilks’ λ = 0.99, P = 0.228); however, there was an interaction (Wilks’ λ = 0.96, P < 0.001) between treatment and plant status (desirable/non-desirable). Densities of desirable seedlings were nearly double those of non-desirable seedlings in drawdown treatments, while the converse was true for the flooded treatment. Drawdown treatments produced greater desirable stem densities and flooded treatments produced greater non-desirable stem densities (Table 4).
Stem density varied between plant status and time period (Wilks’ λ = 0.98, P < 0.001), where subsequent ANOVAs (F = 7.24, P < 0.001) demonstrated that germination was similar between desirable and non-desirable species during the first 30 days (Table 5). However, irrespective of simulated treatment, stem densities of desirable seedlings was greatest during the second 30-day period, while non-desirable seedling stem densities were greatest during the last two 30-day periods (Table 5). Interactions also occurred between simulated treatments and time period (Wilks’ λ = 0.99; P = 0.036). Subsequent ANOVAs demonstrated that seedling densities varied among time periods (F = 2.86, P = 0.036). Seedling germination was greatest during the first two time periods for the drawdown treatment and germination was greatest through the first three time periods for the flooded treatment (Table 5). Stem density varied between treatment and moist-soil managed wetlands (Wilks’ λ = 0.9751, df = 6, P < 0.001) and interactions were found where treatment had an effect on stem density within each moist-soil managed wetland (Table 6).
Stem densities varied between treatment and managed and unmanaged moist-soil wetlands (Wilks’ λ = 0.9914, df = 1, P < 0.0007), where stem densities varied between managed and unmanaged wetlands (F = 11.63, P < 0.007). Stem density in managed moist-soil wetlands under drawdown were more similar to unmanaged flooded moist-soil managed wetlands, while moist-soil managed wetland under flooded conditions were similar to unmanaged moist-soil wetlands under drawdown conditions (Table 7). Stem density varied between time periods and managed/unmanaged moist-soil wetland (Wilks’ λ = 0.9970, df = 3, P < 0.2711) (Table 7), where stem density differences in desirable and non-desirable species, varied among time periods (Table 8). Desirable species had the greatest stem densities in the first two time periods while the first three time periods produced the greatest stem densities for non-desirable species (Table 8).
Discussion
Drawdown and flooded treatments had approximately 32 % of their species in common, slightly higher than van der Valk and Davis (1978), who reported that drawdown and flooded treatments had only approximately 25 % species similarity. As little as 2 cm of standing water may significantly influence seed germination (van der Valk and Davis 1978), where all available seeds contained within the seed bank may not germinate under either treatment condition (van der Valk and Davis 1978). However, the drawdown treatment had more seedlings germinate throughout the entire study, similar to Smith and Kadlec (1983) who found that more species germinated in moist than submerged treatments and suggested there is greater potential for species composition change under moist field conditions. Several factors may influence species composition change under moist field conditions. Seeds may respond to favorable varying temperatures, light, oxygen regimes as well as in soil, lack of canopy, and drawdown conditions that provide suitable germination conditions to be exploited (Leck 2003). Baldwin et al. (2001) also found that twice as many species and five times greater individual seedlings emerged from drawdown conditions than under flooded conditions. Therefore moist-soil conditions (i.e., drawdown) should be created as early as mid-March to produce annual emergent desirable species for continual seed bank renewal.
Fredrickson and Taylor (1982) suggested that slow early season drawdowns will produce a more desirable, dense, and diverse vegetative community that results in greater seed production. This greater seed production promotes desirable plant species expansion and provides essential food resources for migrating and wintering waterfowl a goal of many wetland wildlife managers (Fredrickson and Taylor 1982; Lane and Jensen 1999; Strader and Stinson 2005). Therefore utilizing moist-soil techniques should maximize production of naturally occurring wetland vegetation. By emulating and manipulating natural wetland functions (e.g., hydrology and successional stage) via precise control of hydrology and manipulation of plant succession, wildlife managers can achieve desired plant communities and provide habitat requirements for a variety of wildlife species throughout their annual cycles (Lane and Jensen 1999). In the playas of Texas, Haukos and Smith (1993) suggested moist-soil conditions should be created as early as possible in April to allow for desirable plant species germination, such as smartweeds and annual grasses, and reported that plants germinating early in April had greater overall seed production.
As there was a rapid response from early and continuous germinators in the drawdown treatment, drawdowns should promote establishment of desirable wetland plant species such as pink smartweed (P. pensylvanicum), nodding smartweed, curly dock (Rumex crispus), and barnyard grass (Haukos and Smith 2001). Early and continuous germinators are species that germinate rapidly after exposure to drawdown conditions and then proceed with low germination rates (i.e., early) during the remainder of the growing season or produce seedlings at the same rate (i.e., continuous) throughout the growing season under drawdown conditions (Haukos and Smith 2001) such as barnyard grass and smartweeds which can produce 1,350 kg/ha of seeds (Fredrickson and Taylor 1982; Laubhan and Fredrickson 1992; Gray et al. 1999; Sherfy and Kirkpatrick 1999; Bowyer et al. 2005). Many early and continuous germinators are considered desirable to waterfowl managers due to their ability to provide food for wintering and migrating waterfowl (Fredrickson and Taylor 1982).
More than half of all desirable species germinated within the first 30 days of exposure and >80 % within the first 60 days, which mirrors germination rates in playas, where germination was initiated within the first 30 days and after 90 days of exposure 63 and 77 % of seedlings germinated in moist and flooded conditions, respectively (Pederson 1983; Haukos and Smith 1997; Haukos and Smith 2001). Similarly, Welling et al. (1988) found that nearly all seed bank germination occurred in the first 2 months of exposure to drawdown treatments in the prairie pothole region. As desirable species will typically germinate within 60 days of drawdown conditions, specific plant species composition can be achieved in managed wetlands via strategic drawdown and flooding treatments. However, managers should keep in mind that non-desirable species germinated under flooded conditions on a consistent basis while under the drawdown condition they will not germinate until later in the growing period. For example, in this study, >50 % of non-desirable species germinated in the last two 30-day time periods. Managers should be conscious of water depth, as Baldwin et al. (2001) reported that higher water levels negatively influenced vegetation growth and seed germination in field, greenhouse, and seed-bank experiments and subsequently stated that shallow flooding for a month early in the growing season was a more important determinant of community composition than later flooding even if it occurred longer as well as Capon (2007) who also found that duration of submergence, rate of drawdown, and seasonal timing all have the potential to influence soil seed bank response. Although 27 of 57 known species to occur on the study area were actually recorded in these germination experiments (see Collins 2012), not all species growing on the site will be represented in its seed bank (van der Valk et al. 1992). Seed bank experiments not only reflect last year’s vegetation, but also, to a limited extent, the immediate past vegetation (Leck and Simpson 1987). Field level data collected on the moist-soil managed wetlands on RCWMA found 50 species were present over the 3 years of data collection, and typically averaged 25 species per calendar year of data collection which is consistent with 27 observed during germination trials (Collins 2012). Compositional changes will occur due to differences in germination environment, effects of management practices such as disking and flooding duration and depth, as well as maturation of managed moist-soil wetlands. If germination of a certain targeted species assemblage is desired, knowledge of seed bank composition and expression studies will drive management decisions towards treatments to promote germination and growth of desired species (Smith and Kadlec 1983). Moist-soil wetlands on RCWMA are relatively new, so many annual seed producing moist-soil plant species were present both in field vegetative transect data (Collins 2012) and seed bank data. Generally, the most prolific seed producers and desirable plants for waterfowl were annuals that dominate early successional seral stages (i.e., new wetlands) (Strader and Stinson 2005). Therefore, proper germination conditions were met for many of the species both in the greenhouse and in field conditions. Desirable species were present in greater densities under moist conditions and within the first 60 days of exposure to moist conditions. Also managed moist-soil wetlands had greater moist-soil plant densities under moist conditions than unmanaged moist-soil wetlands. Managed moist-soil wetlands had greater mean seedling germination than the, unmanaged wetlands, for which Triangle, Gut water control is greatly reduced. These results should be encouraging to wetland managers because it shows that managers can produce desirable annual moist-soil plant species through water manipulations (i.e., frequent drawdown and flooding conditions), even on newly created moist-soil managed wetlands.
Baldwin et al. (2001) documented negative impacts on vegetation due to greater water depths in both field and greenhouse conditions. Inhibitory effects of flooding on vegetative growth and seedling recruitment have been widely documented (Galinato and van der Valk 1986; McKee and Mendelssohn 1989; Baldwin et al. 2001). As the managed moist-soil wetlands age, greenhouse seed bank expression experiments and field scale transect data should look similar in species composition, as dominant species persist under consistent water management. However, if management practices are inconsistent or objectives vary annually, seed bank and field scale composition may diverge. For example, wild millet widely occurred within moist-soil managed wetlands (Collins 2012), but was never recorded in the seed bank expression experiments, perhaps due to lack of proper germination conditions. Conversely, toothcup was a dominant species in both seed bank expression experiments and vegetation transects during 2004 (Collins 2012), but its relative density and dominance dropped to extremely low and irrelevant quantities in 2005 and 2006 (Collins 2012), due to longer inundation periods and greater water depths in the field during the 2005 growing season. This was also true for vegetation data from August 2004 which detected red-rooted flatnut sedge as a dominant species found in all four moist-soil wetlands, and the seed bank data also reflects this concordance (Collins 2012). However, it was not detected again on field scale transects during the next 2 years, which indicates that germination conditions were only met in 2004 germination for red-rooted flatnut sedge. For example, Smith and Kadlec (1983) found that germination conditions were not met for Tamarix (Tamarix pentandra), curly-leaf pondweed (Potamogeton crispus), and fennel pondweed (P. pectinatus) in seed bank trials, although they occurred in the field. They postulated that few seeds were present in their samples, perhaps due to poor seed recruitment and germination under field conditions experienced in that study. van der Valk and Davis (1978) also reported this same phenomenon for seeds of both bur-reed (Sparganium) and river bulrush (Scirpus fluviatilis), where discrepancies were observed between field and seed bank samples.
Within many greenhouse experiments, some species might not germinate due to competition, allelopathy, poor germination conditions, and limited number(s) of seeds in samples. Keddy (1999, 2000) suggested that prediction of the presence and abundance of a particular species would require foresight regarding how these various variables (i.e., hydrology, competition, allelopathy, and disturbance) would act on germination and other life history traits. One possible way to corroborate seed bank and field transect data would be to use growth chambers in which the environment can be controlled and allow a longer growing period to express the seed bank to its full potential.
The successional model proposed by van der Valk (1981) for freshwater wetlands dependent on periodic changes in hydrology (i.e., water level) can be applied to all the moist-soil wetlands on RCWMA. van der Valk (1981) postulated that wetland floristic composition normally results from (1) destruction of all or some of the existing vegetation by pathogens, herbivores, or man, (2) changes in the physical or chemical habitat conditions (i.e., change in water or nutrient levels) that favor the growth of some species over others, (3) interactions among plants (i.e., competition, allelopathy), or (4) the invasion and establishment of new species. Destruction of existing vegetation and physical conditions occurred during field-level drawdown and flooding allow for annual seedlings to germinate and begin the process of establishment. Specifically, changes in water levels should allow for nutrient cycling, plant senescence, and subsequent decomposition of the plant litter, allowing new seedlings to germinate during the growing season when drawdown occurs (van der Valk and Davis 1978; van der Valk 1981; van der Valk et al. 1992; Collins 2012).
References
Anderson JT, Smith LM (2000) Invertebrate response to moist-soil management of playa wetlands. Ecol Appl 10:550–558
Araki S, Washitani I (2000) Seed dormancy/germination traits of seven Persicaria species and their implication in soil seed-bank strategy. Ecol Res 15:33–46
Baldwin AH, McKee KL, Mendelssohn IA (1996) The influence of vegetation, salinity, and inundation of seed banks of oligohaline coastal marshes. Am J Bot 83:470–479
Baldwin AH, Egnotovich MS, Clarke E (2001) Hydrological change and vegetation of tidal freshwater marshes: field, greenhouse, and seed-bank experiments. Wetlands 21:519–531
Boedeltje G, ter Heerdt GNJ, Bakker JP (2002) Applying the seedling-emergence method under waterlogged conditions to detect the seed bank of aquatic plants in submerged sediments. Aquat Bot 72:121–128
Bossuyt B, Honnay O (2008) Can the seed bank be used for ecological restoration? An overview of seed bank characteristics in European communities. J Veg Sci 19:875–884
Bowyer MW, Stafford JD, Yetter AP, Hine CS, Horath MM, Havera SP (2005) Moist-soil plant seed production for waterfowl at Chautauqua National Wildlife Refuge, Illinois. Am Midl Nat 154:331–341
Budelsky RA, Galatowitsch SM (1999) Effects of moisture, temperature, and time on seed germination of five wetland Carices: implications for restoration. Restor Ecol 7:86–97
Capon SJ (2007) Effects of flooding on seedling emergence from the soil seed bank of a large desert floodplain. Wetlands 27:904–914
Capon SJ, Brock MA (2006) Flooding, soil seed bank dynamics and vegetation reliance of a hydrologically variable desert floodplain. Freshw Biol 51:206–223
Casanova MT, Brock MA (2000) How do depth, duration and frequency of flooding influence the establishment of wetland plant communities? Plant Ecol 147:237–250
Collins DP (2012) Moist-soil managed wetlands and their associated vegetative, aquatic invertebrate, and waterfowl communities in east-central Texas. PhD Dissertation, Stephen F. Austin State University, Nacogdoches
Correll DS, Johnston MC (1979) Manual of the vascular plants of Texas. University of Texas at Dallas Press, Richardson
Flynn KM, Mendelssohn IA, Wilsey BJ (1999) The effects of water level management on the soils and vegetation of two coastal Louisiana marshes. Wetl Ecol Manag 7:193–218
Fredrickson LH, Taylor TS (1982) Management of seasonally flooded impoundments for wildlife. United States Fish and Wildlife Service, Resource Publication 148
Galinato ML, van der Valk AG (1986) Seed germination traits of annuals and emergents recruited during drawdowns in the Delta Marsh, Manitoba, Canada. Aquat Bot 26:86–102
Gray MJ, Kaminski RM, Weerakkody G (1999) Predicting seed yield of moist-soil plants. J Wildl Manag 63:1261–1268
Gurnell AM, Boitsidis AJ, Thompson K, Clifford NJ (2006) Seed bank, seed dispersal and vegetation cover: colonization along a newly created river channel. J Veg Sci 17:665–674
Haukos DA, Smith LM (1993) Moist-soil management of playa lakes for migrating and wintering ducks. Wildl Soc Bull 21:288–298
Haukos DA, Smith LM (1997) Common flora of playa lakes. Texas Tech University Press, Lubbock
Haukos DA, Smith LM (2001) Temporal emergence patterns of seedlings from playa wetlands. Wetlands 21:274–280
Hopfensperger KN, Engelhardt KAM, Lookingbill TR (2009) Vegetation and seed bank dynamics in a tidal freshwater marsh. J Veg Sci 20:767–778
Howard RJ, Mendelssohn IA (1995) Effect of increased water depth on growth of a common perennial freshwater-intermediate marsh species in coastal Louisiana. Wetlands 15:82–89
Kadlec JA, Smith LM (1984) Marsh plant establishment on newly flooded salt flats. Wildl Soc Bull 12:388–394
Keddy P (1999) Wetland restoration: the potential for assembly rules in the service of conservation. Wetlands 19:716–732
Keddy P (2000) Wetland ecology principles and conservation. Cambridge University Press, Cambridge
Krebs CJ (1999) Ecological methodology, 2nd edn. Benjamin/Cummings, Menlo Park
Lane JJ, Jensen KC (1999) Moist-soil impoundments for wetland wildlife. US Army Corps of Engineers. Technical Report EL-99-11
Laubhan MK, Fredrickson LH (1992) Estimating seed production of common plants in seasonally flooded wetlands. J Wildl Manag 56:329–337
Leck MA (2003) Seed-bank and vegetative development in a created tidal freshwater wetland on the Delaware River, Trenton, New Jersey, USA. Wetlands 23:310–343
Leck MA, Graveline KJ (1979) The seed bank of a freshwater tidal marsh. Am J Bot 66:1006–1015
Leck MA, Simpson RL (1987) Seed bank of a freshwater tidal wetland: turnover and relationship to vegetation change. Am J Bot 74:360–370
Leck MA, Simpson RL (1993) Seeds and seedlings of the Hamilton Marshes, a Delaware River tidal freshwater wetland. Proc Acad Nat Sci Phila 144:267–281
Li EH, Liu GH, Li W, Yuan LY, Li SC (2008) The seed-bank of a lakeshore wetland in Lake Honghu: implications for restoration. Plant Ecol 195:69–76
McFarland DG, Shafer DJ (2011) Protocol considerations for aquatic plant seed bank assessment. J Aquat Plant Manag 49:9–19
McKee KL, Mendelssohn IA (1989) Response of freshwater marsh plant community to increased salinity and increased water level. Aquat Bot 34:301–316
Middleton BA (2003) Soil seed banks and the potential restoration of forested wetlands after farming. J Appl Ecol 40:1025–1034
Moore DRJ, Keedy PA (1988) Effects of water-depth gradient on the germination of lakeshore plants. Can J Bot 66:548–552
Natural Resource Conservation Service (2002) Soil survey of Freestone County, Texas. United States Department of Agriculture
Nicol JM, Ganf GG, Pelton GA (2003) Seed banks of a southern Australian wetland: the influence of water regime on the final floristic composition. Plant Ecol 168:191–205
Pederson RL (1983) Abundance, distribution, and diversity of buried seed populations in the Delta Marsh, Manitoba, Canada. PhD Dissertation, Iowa State University, Ames
Pederson RL, Smith LM (1988) Implications of wetland seed bank research: a review of Great Britain and prairie marsh studies. In: Wilcox DA (ed) Interdisciplinary approach to freshwater wetland research. Michigan State University Press, East Lansing
Sherfy MH, Kirkpatrick RL (1999) Additional regression equations for predicting seed yield of moist-soil plants. Wetlands 19:709–714
Shipley B, Parent M (1991) Germination responses of 64 wetland species in relation to seed size, minimum time to reproduction and seedling relative growth rate. Funct Ecol 5:111–118
Shipley B, Keddy PA, Moore DRJ, Lemky K (1989) Regeneration and establishment strategies of emergent macrophytes. J Ecol 77:1093–1110
Smith LM, Kadlec JA (1983) Seed banks and their role during drawdown of a North American marsh. J Appl Ecol 20:673–684
Smith LM, Pederson RL, Kaminski RM (eds) (1989) Habitat management for migrating and wintering waterfowl in North America. Texas Tech University Press, Lubbock, 560 pp
Strader RW, Stinson PH (2005) Moist-soil management guidelines for the U.S. Fish and Wildlife Service Southeast Region
Stutzenbaker CD (1999) Aquatic and wetland plants of the Western Gulf Coast. Texas Parks and Wildlife Press, Austin
ter Heerdt GNJ, Verweij GL, Bekker RM, Bakker JP (1996) An improved method for seed-bank analysis: seedling emergence after removing the soil by sieving. Funct Ecol 10:144–151
Texas Parks and Wildlife Department (2005) Texas comprehensive wildlife conservation strategy. Texas Parks and Wildlife Department, Austin
USDA, NRCS (2011) The PLANTS database. National Plant Data Team, Greensboro. http://plants.usda.gov. Accessed 23 June 2011
van der Valk AG (1981) Succession in wetlands: a Gleasonian approach. Ecology 62:688–696
van der Valk AG, Davis CB (1978) The role of seed banks in the vegetation dynamics of prairie glacial marshes. Ecology 59:322–335
van der Valk AG, Pederson RL, Davis CB (1992) Restoration and creation of freshwater wetlands using seed banks. Wetl Ecol Manag 1:191–197
Welling CH, Pederson RL, van der Valk AG (1988) Recruitment from the seed bank and the development of zonation of emergent vegetation during a drawdown in a prairie wetland. J Ecol 76:483–484
Acknowledgments
Financial and logistical support for this research was provided by the Texas Parks and Wildlife Department and the Arthur Temple College of Forestry and Agriculture (MacIntire-Stennis) at Stephen F. Austin State University. We thank Eric Woolverton, Edwin Bowman, Gary Rhodes, Kevin Kraai, and Matt Symmank for field and logistical support. We appreciate comments by C. Comer, B. Oswald, D. Scognamillo, G. Esslinger, and B. Howe on earlier drafts of this manuscript.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Collins, D.P., Conway, W.C., Mason, C.D. et al. Seed bank potential of moist-soil managed wetlands in east-central Texas. Wetlands Ecol Manage 21, 353–366 (2013). https://doi.org/10.1007/s11273-013-9307-5
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11273-013-9307-5