
Abstract
Most countries sharing the Amazon basin have signed the Ramsar Convention on Wetlands of International Importance but still lack complete wetland inventories, classification systems, and management plans. Amazonian wetlands vary considerably with respect to hydrology, water and soil fertility, vegetation cover, diversity in plant and animal species and primary and secondary productivity. Here, we propose a classification system of major natural habitats of Amazonian white-water river floodplains (várzeas) based on hydrological, water and soil chemistry and biological parameters. The Amazonian várzea is one of the largest Amazonian wetlands. It is exceptionally rich in plant and animal species and plays important roles in landscape history, evolution, hydrology and biogeochemical cycles of the Amazon basin. Most of Amazonia’s rural population lives in or along the várzea, emphasizing the economic importance of its natural resources. Our classification indicates five major systems, which are subdivided into 10 main habitats and up to 40 functional (vegetation) units of which the most important mesohabitats are described. We understand this classification as a dynamic system, as it is open to the inclusion of future research attempts and habitats without affecting the entire classification system. Our classification may be used for scientific purposes, such as comparative studies on biomass, productivity, biogeochemical cycles and biodiversity. Also, because the classification builds on habitat types and/or vegetation and functional units already distinguished by the local population it may be especially useful in guiding intelligent use of várzea habitat for specific management activities, such as agriculture, animal husbandry, forestry, fisheries, and conservation.
Similar content being viewed by others
Avoid common mistakes on your manuscript.
Introduction
In recent years, worldwide interest on the Amazon basin has grown because of its role in the global climate. Discussions about hydrological and biogeochemical cycles, carbon budget and methane emission as well as on the maintenance of biodiversity have generally concentrated on the upland rain forest and have generally neglected the extended wetland areas (Sousa Jr. et al. 2011) which make up about 30 % of the Amazon basin. In 1993, Brazil signed the Ramsar Convention, which requires a national policy for the wise use and protection of wetlands and their components. However, the conduction of inventories has been slow (Diegues 1994, 2002) and the scientific basis for a classification remains poor. Only in 2011, Junk et al. presented a classification of the major Amazonian wetland types. Each of them covers tens of thousands square kilometers and contains a large number of different habitats, which have not yet been classified.
Of major importance are the extended floodplains which accompany the large Amazonian lowland rivers. Amazonian rivers are classified according to their water quality ranging from nutrient-rich white-water rivers to nutrient-poor clear water and black-water rivers (Sioli 1950; Junk et al. 2011). The floodplains along the large white-water rivers, locally and in scientific literature called várzeas, cover an area of about 400,000 km2 (Melack and Hess 2010). The várzeas are of great economic importance. According to the last census of the Brazilian population (IBGE 2010), 3.1 million people constitute the rural population of the States of Amazonas and Pará. From this amount, more than 60 % live in the várzea. Most timber (Schöngart 2008; Wittmann and Oliveira Wittmann 2010) and also part of the beef, fruit and vegetable consumed in urban centers of the Amazon derive from the várzea (Junk et al. 2000). The Amazonian inland fishery plays an important role in the protein supply of the local population (Junk et al. 2000) and 70–80 % of the catch derives from nutrient-rich white-water rivers and their várzeas.
In recent times, an increasing pressure on várzea resources has created conflicts between different stake-holder groups and has put in risk the ecological integrity of the entire system. Therefore, a classification system based on scientific criteria that subdivides várzea habitats into easily distinguishable units and allows stakeholders to address specific regulations for specific habitats is urgently required for efficient management and conservation purposes. This article presents the first classification of the major natural habitats of the Amazonian várzeas using hydrological, soil- and water-chemical and botanical parameters for habitat characterization.
Genesis and ecological characterization of large Amazonian white-water river floodplains (várzeas)
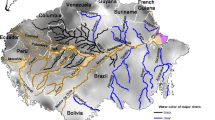
Várzeas accompany the main stem of the Amazon River and its large white-water tributaries Madeira, Purús, Juruá and Japurá (Fig. 1). These rivers transport large amounts of dissolved and suspended matter from the Andes to the Atlantic Ocean. Changes in dissolved and suspended load are also observed during the hydrological cycle (Fig. 1). At a sampling station downriver of the central Amazonian City of Manacapuru concentrations of suspended matter are highest at low-water stages (300–450 mg l−1) and lowest during falling water levels (110–150 mg l−1) (Meade 1994). Seasonal changes in storage and remobilization of suspended sediment in the Amazon main stem have been demonstrated between Manacapuru and the city of Óbidos. During early and middle rising stages sediments are temporarily deposited, while near peak and falling stages sediments are re-suspended (Meade et al. 1985).

Distribution of central Amazonian white-water river floodplains (várzeas), and hydrographs of major white-water rivers
The development of the recent floodplain of the Amazon River started about 15,000 years ago at the end of the last glacial period. At that time, the sea level was about 130 m lower than today and the Amazon River had eroded a deep valley into the soft sediments which had been deposited during former interglacial periods (paleo-várzeas) (Irion et al. 1997, 2010). Because of the sea-level rise the river filled the valley with recent sediments, a process that is still ongoing as indicated by the large number of large floodplain lakes at the lower Amazon and the formation of internal deltas in these lakes by inflowing river water. The recent floodplain is flooded during high-water levels and is dry during low-water levels. The zone of periodic flooding can span laterally several tens of kilometers on either side of the river channel and is called the aquatic terrestrial transition zone (ATTZ; Junk et al. 1989).
Dendritic Ria-lakes at the edges of the recent várzea are remnants of the glacial low sea-level stages that are carved deeply into the adjacent upland. They represent drowned valleys of black-water and clear-water rivers that have not yet filled with recent sediments. In contrast, paleo-várzeas are remnants of interglacial periods with sea-levels even higher than today. Some of them reach levels several meters above the recent floodplain (Irion et al. 2010).
Erosion and deposition processes lead to a complex, small-scaled mosaic of hydro-morphological structures that determine the current várzea landscape. The dynamics of channel migration increase from East to West with increasing declivity. At the foothills of the Andes, the rivers reshape their floodplains completely in a few hundred years (Kalliola et al. 1992). In central Amazonia the position of the main channels is relatively stable, despite visible erosion of many shores and a few sedimentation hot spots, e.g. near the city of Manaus, upriver of the confluence with the Negro River. Sediment dating in the central lake of Marchantaria Island near Manaus revealed a maximum age of the lake of somewhat more than 5,000 years (Irion et al. 1997). Potteries found in archeological sites of Careiro Island in front of Manaus were dated with 1,000 to 2,000 years (Sternberg 1960). We assume that low declivity and rocky outcrops in the river valley hinder major channel migration in this central Amazonian region.
Deposition of coarse sediments occurs mostly in sedimentation areas along the main channel, at the entrance of the channels into the lakes, or, during very high water levels, on the natural levees that are formed along the channels (Peixoto et al. 2009). Large floodplain areas are protected by the levees and receive only very low amounts of fine particles during the floods (Mertes 1994). These hydro-morphological processes are very important from an ecological point of view, because they lead to the formation of new habitats by sediment deposition and a setback of climax vegetation to earlier successional stages by erosion.
The sediment load contains considerable amounts of smectite, illite and kaolinite. Smectite has a high ion-exchange capacity and illite releases potassium during the weathering process, which results in a relatively high fertility of the alluvial soils. The water is slightly acidic to neutral (pH values 6–7), dominated by the alkali-earth metals Ca and Mg and by carbonates (Furch and Junk 1997). Electric conductivity decreases from the Andes in the West to the mouth in the East by dilution through tributaries with electrolyte-poor clear and black waters, originating from the Precambrian Shields in the North and South of the central Amazon basin. Near the Andes it is about 120–140 μS cm−1, at the lower course about 30–50 μS cm−1 during the wet and dry season, respectively (Gibbs 1967). Near the coast, electric conductivity can rise because of entrance of marine water and aerial salt deposition.
High nutrient content of water and sediments stimulates primary production. Aquatic habitats are colonized by algae and aquatic and semi-aquatic macrophytes. Low-lying areas of the ATTZ are colonized during the dry phase by annual and perennial grasses, sedges and herbaceous plants. Várzea areas covered at average less than 8 months per year with water are covered by different types of floodplain forests (see section Areas covered by different types of forest). Early-successional forest stages and forests on disturbed areas are often colonized by annual and perennial vines that can be considered as indicators of increased fertility. The change between terrestrial and aquatic phases during the annual cycle leads to the occupation of the same area by both aquatic and terrestrial plant communities at the different times of the year increasing primary production and nutrient dynamics (Junk and Piedade 1997).
Parameters used for the habitat classification
The new classification is based on the flood-pulse concept (Junk et al. 1989; Junk and Wantzen 2004; Junk 2005). Parameters used are hydrology, water and soil chemistry and species distribution and community structure of higher vegetation as described by Junk et al. (2011). The Amazon River and its large tributaries show a monomodal, predictable flood pulse with high amplitude and a sinusoidal shape (Fig. 1). Floodplain habitats are subjected to this pulse to different degrees depending on their position along the flooding gradient. The highest habitats on levees are flooded every couple of years for a few weeks while the lowest habitats are dry every couple of years during a few weeks only. Most habitats fall completely dry during low-water period, but some are waterlogged during the entire year because of hydro-morphologic peculiarities. Therefore, length and depth of flooding are important parameters for habitat differentiation (Junk et al. 2011).
Most várzea habitats have a high nutrient status that is determined by the amount of dissolved and suspended nutrients from the periodic inflow of river water. Duration of connectivity with the main river and habitat-internal cycles modify the nutrient status, but do not change the characteristic chemical attributes of soils and water of várzea habitats (Furch and Junk 1997). Therefore chemical parameters of soils and water have low importance for the classification of different várzea habitats. Differences exist only in habitats at the edges of the várzea, where water from the surrounding nutrient-poor uplands (terra firme) enters the várzea by tributaries, e.g. in Ria-lakes (Junk et al. 2011). An exception is soil granulometry. Sandy soils in deposition areas suffer increased drought stress during the terrestrial phase, but provide better oxygenation after flooding because of high porosity. Clay soils have a higher water retention capacity during the terrestrial phase, but suffer greater oxygen deficiency during water logging and flooding because of low porosity. The impact on forest communities is shown by habitats 4.3.1.1 and 4.3.1.2. In comparison with other wetlands, however, chemical and physical parameters of water and soils have great importance. Nutrient-rich várzeas are separated from nutrient-poor black- and clear-water floodplains by chemical parameters, and the impact of these parameters on primary production, secondary production, and management is shown by Junk et al. (2011).
Because of their longevity and their stationary behavior, higher plants best represent the environmental conditions of their respective habitats. Herbaceous plants indicate past conditions at temporal scales of months or years while trees at decades and centuries. Occurrence of species, composition of plant communities, biomass and NPP are important parameters used to characterize habitats. Animals are not considered in this classification because of their mobility and an insufficient data base about their habitat requirements. However, they may be included in further attempts to expand the present classification for other biological groups.
Humans have utilized the resources of the várzea since millennia (Roosevelt 1999). Today, subsistence agriculture, cattle ranching and timber exploitation have dramatically modified the vegetation cover (Goulding et al. 1996; Junk et al. 2000). For instance, undisturbed high-várzea forests are found only in remote areas or in protected reserves such as the Mamirauá Sustainable Development Reserve near Tefé or in indigenous reserves. Many areas, covered today by different communities of annual and perennial grasses, sedges and herbaceous plants or early-successional forest stages, have been formerly covered by different types of mature floodplain forests.
The new classification of várzea habitats
Rationale
Our classification system differentiates between five major categories: permanently terrestrial, permanently aquatic, periodically terrestrial, periodically aquatic and swamp habitats. Permanently aquatic habitats are included because of the exchange of water, dissolved and particulate material, and biota between the river, permanent lakes and the ATTZ. Permanently terrestrial habitats are included because of their influence on the species composition of plant communities inside the várzea, mainly on the floodplain forests as shown by Wittmann et al. (2010). Swamp systems differ from the others by representing permanent or multiannual waterlogged conditions.
Habitats in pulsing systems which pass through both terrestrial and aquatic phases often present difficulties in delineation and resist clear-cut classifications. Depending on the viewpoint of the observer, they can be classified as periodically aquatic or periodically terrestrial ones. Our classification system bases on communities and successional stages of higher plants. Most of the plant species can be considered as terrestrial despite of many adaptations to periodic life in water (e.g., trees and terrestrial herbaceous grasses, sedges and herbs, or semiaquatic (e.g. palustric herbaceous grasses, sedges and herbs). Therefore we denominate habitats covered by these species as periodically terrestrial habitats. However, a few important habitats, such as channels and many open water areas fall dry for short periods and do not show characteristic higher plant communities for the classification. Others, such as beds of floating aquatic macrophytes play an important role in the aquatic phase because of large extension and high productivity, and therefore require habitat status. But the same area is occupied by terrestrial grasses and herbaceous plants during the terrestrial phase and plays an important role as a terrestrial habitat. In this case, it is logical to list them twice in the classification, as periodically aquatic and periodically terrestrial habitat with the respective species (categories 2.2.2 and 4.2.2).
A synthesis of the new classification system of the habitats of Amazonian várzeas is given in Table 1. If necessary, the classification allows the introduction of new habitats without affecting the entire system.
General description of major functional units
Lakes
Lakes can be formed between bars (scroll lakes), in abandoned river meanders (oxbow lakes, Fig. 2) in floodplain depressions (depression lakes) and in drowned valleys of tributaries (Ria-lakes) (Fig. 3). Melack (1984) analyzed shape and fetch of 8,050 lakes in the várzea of the Amazon River, 5,010 lakes had a round to oval form and corresponded to the depression lakes in our classification. They are most common in the middle and lower reaches of the Amazon River. 1,530 lakes were levee lakes with an elongated shape and corresponded to the scroll lakes in our classification. They are most common in the upper and middle reaches. 830 lakes had a dendritic form, which is characteristic for Ria-lakes. They are most frequent in the middle reaches. The majority of the 270 oxbow lakes are found in the upper reaches.

Lower Juruá River floodplain with oxbow lakes of different connectivity levels. Source: Landsat5 TM 001 062 (10.06.1997), Instituto Nacional de Pesquisas Espaciais-INPE

Amazon várzea near the confluence of the Solimões and Japurá Rivers (Mamirauá Sustainable Development Reserve), with characteristic hydro-morphological features. 1 = upland; 2 = Ria lakes embedded in the upland; 3 = depression lakes; 4 = ridges and swales with scroll lakes; 5 = river channel; 6 = long floodplain-internal channels (paranas); 7 = shortcuts (furos). Source: Landsat5 TM 001 062 (10.06.1997), Instituto Nacional de Pesquisas Espaciais-INPE
Scroll lakes are elongated, narrow and mostly covered by aquatic macrophytes. Oxbow lakes can have steep shores and less macrophyte cover. Large parts of depression lakes fall dry during the low-water period. Aquatic macrophytes develop in large quantities with rising water level but become uprooted by wind and wave action and drift away with increasing water level. Ria-lakes receive electrolyte-rich white water from the main river and electrolyte-poor black or clear water from the tributaries. During rising water level white water advances into the lake and during falling water level black or clear water advances to the mouth from the tributaries. The occurrence of perennial white water specific grasses such as Echinochloa polystachya, Paspalum repens and P. fasciculatum and some white-water indicator tree species such as Cecropia latiloba and Pseudobombax munguba indicate substantial intrusion of sediment-laden river water into Ria-lakes.
Most lakes are shallow with a maximum depth at high-water level in the western Amazon basin up to 12 m and in the eastern part up to 6 m. Some Ria lakes can have a greater depth. During day-time a thermal stratification develops with surface water temperatures of up to 33 °C and about 28 °C in the hypolimnion at a depth of 3–5 m. High amounts of organic material and high decomposition rates lead to hypoxic or even anoxic conditions in the hypolimnion during the high-water period. Benthos is impoverished. The size and fetch of the lakes determine water circulation and oxygen concentration in the hypolimnion (Melack 1984). In scroll lakes, oxbow lakes and narrow Ria-lakes oxygen concentrations in the hypolimnion are especially low because the lakes are protected against wind-induced turbulences. In permanent lakes with steep shores, dense floating islands, locally called matupá, can develop because permanent water logging retards the decomposition of the organic layer. Water circulation and oxygen availability is better at the bottom of the larger and shallower depression lakes in eastern Amazonia as shown by the occurrence of bivalves in the sediment (Junk unpubl.).
The degree of connectivity between the river and lakes is important for the exchange of water, dissolved and solid substances and the biota. Large lakes are connected to the river most of the time through small channels locally called paranás (long channels) or furos (shortcuts) (Fig. 3), because the large amount of inflowing and out-flowing water keeps the channels open. Small lakes may be isolated for several months and become connected to the river only at high water level. Some white-water tributaries of the Amazon River such as the Purus and Juruá have oxbow lakes embedded in the paleo-várzea that are connected to the river only during extreme floods. With decreasing connectivity, lakes tend to establish their own specific nutrient cycles (Furch and Junk 1997). Lakes isolated at low water are important refuges for the aquatic fauna.
River channels and intermittent water distribution systems
For hydrologists, river channels serve for the transport of water and dissolved and solid substances. For biologists they are habitats, passive dispersal routes and migration pathways for plants and animals. The large river channels transport sandy bed-load in form of giant ripples up to 12 m high (Sioli 1965; Mertes and Meade 1985). At the shores, vegetation belts, mainly of the robust aquatic and semi-aquatic grasses Paspalum fasciculatum, P. repens and E. polystachya provide food and/or habitats for the aquatic fauna and stabilize the sediments favoring the first stages of succession by woody plants. Log jams provide specific habitats at the shore.
Inside the floodplain, water is distributed by minor channels that connect the main river channels with the floodplain lakes. Water flux in these paranás and furos can change direction, transporting water with rising water level from the main channels into the floodplain and from the floodplain to the major river channels with falling water level. The channels are also important active migration- or passive transport-pathways for aquatic plants and animals between the rivers and their floodplains. Along the border of the floodplains adjacent to the uplands there are long paranás, which transport the run-off from the upland to the river (Fig. 3).
Areas covered by different types of grass- and herbaceous plant communities
Low-lying parts of the ATTZ and higher lying areas disturbed by the floods or by human activity are colonized by grasses, sedges and herbaceous plants (Fig. 4). Junk and Piedade (1993) list 388 herbaceous species in the várzea near Manaus, of which 44 can be considered aquatic, 20 palustric and 17 aquatic with a terrestrial phase. Numbers of aquatic and palustric macrophytes is low because high water-level fluctuations and low water transparency obstruct the growth of submersed and emergent plants. 28 species are free-floating (Junk and Piedade 1993). These numbers will certainly increase when other várzea areas are sampled. Areas exposed to dry conditions for a few weeks are colonized by annual species with a short life cycle. Many species are ruderal plants and agricultural weeds, which also colonize disturbed moist upland areas along roads (Seidenschwarz 1986). Specimens are recruited from the seed bank in the sediment and/or from seeds transported by the water.

Different habitats and plant communities in the várzea. From bottom to top: Mud and sand flats, annual and perennial terrestrial herbaceous vegetation, pioneer forest on newly build low lying levees and forest of early-secondary stage on slightly higher levees
Along the river, current and wave action hinder the growth of aquatic macrophytes. In lakes annual aquatic species such as Luziola spruceana, Oryza perennis and P. repens develop, interspersed with free-floating species such as Eichhornia crassipes, Salvinia spp., Azolla caroliana and others. After a few weeks, with rising water, these communities become uprooted and drift away. Higher lying areas are dominated by perennial grass species such as E. polystachya, P. repens and P. fasciculatum. The first two species have a preference for aquatic conditions and grow up with the rising water. P. fasciculatum becomes inundated and survives under water. The free-water area above the drowned stands is colonized by free-floating species (Junk and Piedade 1997; Piedade and Junk 2000).
The perennial species maintain a high biomass during the terrestrial phase, which hinders the establishment of trees. Therefore large monospecific stands of these grasses can be observed in new sedimentation areas which are already high enough along the flood gradient to be colonized by pioneer trees. Nonetheless, as these grasses retain sediments, they help in consolidating new areas for eventual colonization of woody species and are important elements in the successional pattern of várzea forests (Wittmann et al. 2010).
Under permanently wet conditions, dead organic material accumulates amongst tightly knit root networks of free-floating plants and floating islands develop. This habitat type contributes an additional nine palustric species, which are not affected by water-level fluctuations. With increasing density of the organic layer, founder species become restricted to the border of the island and are substituted by swamp species such as Scirpus cubensis and other sedges and ferns in the center. When the tree-like Montrichardia arborescens or M. linifera invade the community, floating islands (matupás) can become very thick and be colonized by shrubs and trees (Junk and Piedade 1997).
High primary productivity is an inherent attribute of várzea habitats because of permanent nutrient input from the river and internal nutrient cycles, which accumulate nutrients in the system. Aquatic species capture dissolved nutrients from the water and maintain them in the ATTZ. Terrestrial species take up nutrients from the sediments and, when decomposing, deliver them to the water, increasing the nutrient concentration in the water of the várzea lakes and the ATTZ. Therefore the várzea functions on a higher trophic level than expected by the amounts of nutrients transported into the floodplain by the inflowing river water (Junk 1997). Perennial C4 grasses show a primary production of up to 100 t ha−1 year−1 and a biomass of up to 80 t ha−1 year−1 dry matter (Piedade et al. 1991, 2001). Annual terrestrial and aquatic C3 plants produce up to 50 t ha−1 year−1 dry matter because of the consecutive colonization of the same area by different plant communities during the hydrological cycle (Junk and Piedade 1997). Floodplain forests are also highly productive as shown later.
Areas covered by different types of forest
All várzea areas that show a mean aquatic phase of less than 8 months year−1 can be colonized by woody species that form floodplain forest (Fig. 4). Some shrubs tolerate even 9 months of flooding. With more than 1,000 flood-tolerant species, Amazonian várzea forests are the most species-rich floodplain forests in the world (Wittmann et al. 2006). Increasing flood stress from the highest levees to low-lying areas leads to species distributions according to their flood tolerance. Wittmann et al. (2002, 2004) differentiates between high-várzea forest that develops on areas flooded on average less than 3 months and to a depth of less than 3 m, and low-várzea forests in higher flooded areas. High-várzea forests colonize the highest natural levees that represent only a small percentage of the total várzea area. In the Sustainable Development Reserve of Mamirauá near the city of Tefé, only about 15 % belong to the high várzea (Wittmann et al. 2002).
Only about 30 % of all tree species occur in both high and low várzea forest types. High-várzea forests show a greater affinity to adjacent terra firme forests of which they share about 30 % of the tree species. Species of low-várzea forests are well adapted to long and deep flooding. Consequently, low-várzea forests share only about 10 % of all species with the adjacent uplands, and they may contain elevated degrees of endemic tree species (Wittmann et al. 2010). Tree species richness in both forest types is lower than in adjacent terra firme forests (up to 250 species ha−1; Ter Steege et al. 2006), and amounts to up to 90 species ha−1 in late-successional low-várzea forests and up to 140 species ha−1 in high-várzea forests (Wittmann et al. 2010). Species composition in both forest types changes considerably from East to West. Western Amazonian várzea is generally richer in tree species than the eastern region (Wittmann et al. 2006). This trend of increasing tree species diversity from the East to the West was also described for non-flooded terra firme forest, and is interpreted as being the response to increasing precipitation, decreasing precipitation seasonality, and increasing soil fertility (Ter Steege et al. 2006; Hoorn et al. 2010).
In addition to the flood stress, sedimentation and erosion processes constantly affect the várzea forest. Forest succession in sedimentation areas near the river channel starts with the colonization of Salix martiana and Alchornea castaneifolia, and in western Amazonia, with Tessaria integrifolia. These species tolerate long-term flooding and high sedimentation rates (Wittmann and Parolin 2005). In lakes with little sediment deposition flood tolerant shrubs such as Eugenia spp. and Symmeria paniculata are pioneer species. In areas with low sedimentation rates a low-várzea forest develops that may be stable for millenia, because the low sediment input raises the soil surface only very slowly (Fig. 5). In high sedimentation areas the low-várzea forest passes to a high-várzea forest that is the final successional forest stage in the várzea. A specific forest type is the swamp forest, locally called chavascal. It is a species-poor forest in habitats that are waterlogged also during low-water periods. Extended swamp forest areas in the várzea occur when the floodplain is exceptionally large, as near the confluence of white-water rivers in the Central Amazon (e.g., Solimões and Japurá Rivers—Mamirauá Sustainable Development Reserve; Solimões and Purus Rivers—Piagaçu-Purus Sustainable Development Reserve). In addition, swamp forests occur along the distal part of the várzea because the terra firme provides water during the low-water period, and at the lower Amazon because of the smaller water-level fluctuations. Small swamp forest patches can also develop in lakes on floating islands. The different successional stages and a time frame for the different types of plant successions in várzea floodplains are presented in Fig. 5.

Development of várzea floodplain forests in different environments (Wittmann et al. 2010)
Along the gradient of forest succession aboveground-woody biomass stocks increase from 18 ± 3 Mg ha−1 (early successional shrub communities on areas with high sedimentation rates) to 117 ± 9–261 ± 10 Mg ha−1 (intermediate successional forests) (Schöngart et al. 2010). Late-successional forests in the low várzea have accumulated woody-biomass stocks of 230 ± 9 to 239 ± 11 Mg ha−1. In the low várzea aboveground NPP declines along the successional gradient from 28.2 to 31.8 Mg ha−1 year−1 in the intermediate successional forests to 13.3 Mg ha−1 year−1 in the late-successional forests (Schöngart et al. 2010). For high-várzea forests, swamp forest and early-successional shrub communities on areas with low sedimentation rates no data on biomass stocks and NPP are available for the Central Amazonia. Primary production of the high-várzea forest is three times higher than that of adjacent upland forests and four times higher than that of nutrient-poor black-water inundation forests (high igapó forest) (Schöngart et al. 2010).
Vegetation-free habitats
River channels and the central parts of lakes are free of higher vegetation. In lakes, phytoplankton is rare in the turbid river channels. During the terrestrial phase, vegetation-free habitats in the ATTZ cover only small areas because herbaceous plant growth is very vigorous in newly available terrestrial habitat. Major habitats are steep river banks, sand banks and rocky outcrops and are important for certain plant and animal species. For example, dry, steep river banks are used by king fishers during the terrestrial phase and during the aquatic phase freshwater crabs and armored catfishes build their nesting hollows. Sand bars are used by river turtles and some shore birds for nesting (Petermann 1997).
There is little information about the occurrence of bed rocks in the middle and lower Amazon main stem but they certainly influence the channel dynamics of the Amazon and its tributaries. The relative stability of the central Amazon main stem may be explained in part by the existence of obstacles of laterite and sand stone that constrain the free meandering behavior of the river. Laterites can be observed at low water at the shores near Manaus and sand stones can be observed at the mouth of the Paraná de Curari. The rapids in the Madeira River at Porto Velho and the reservoirs under construction there are important obstacles for migrating fishes and other aquatic animals, however they can also provide specific habitats for rheophilic plant and fish species.
Adjacent upland systems: paleo-várzeas and tertiary terra firme
The uplands adjacent to the várzeas cannot be considered as wetlands because they do not show wetland attributes. They are, however, important for the várzea because there is an intensive active and passive exchange of plant and animal species between both systems. Paleo-várzeas were built up on sediments at or above the recent floodplain level that were deposited by the Amazon River and its white-water tributaries during Pliocene/Pleistocene interglacial high-water periods. The deposition pattern is complex and still little understood (Irion et al. 2010). All paleo-várzeas show typical hydromorphic sedimentation patterns and mineralogical peculiarities of strongly weathered sediments of Andean origin, including elevated soil fertility. Soils of uplands of tertiary origin are strongly weathered. They contain high amounts of caolinite and sand and are extremely nutrient-poor. Upland tree species become pre-adapted to floodplain conditions in riparian forests along terra firme streams and small rivers and can later colonize the large river floodplains (Wittmann et al. 2010).
Discussion
Classification systems depend on the objectives and the parameters used by the authors. The problems in elaborating a general classification system were discussed by Finlayson and Van der Valk (1995). Those authors pointed out the need to resolve differences among existing systems in the definition of a wetland and how wetland types are defined. Furthermore, they called for the standardization of data collection, storage, and dissemination techniques in order to generate more extensive international inventories.
Many international classification systems were formulated decades ago and often do not satisfy modern scientific requirements and national peculiarities. For instance, despite the existence of large riverine floodplains along the Mississippi, Ohio, and Missouri Rivers, the US Fish and Wildlife Service (USFWS) classification system (Cowardin et al. 1979) does not treat floodplains as a specific wetland category nor does it consider the large habitat diversity of these systems. The classification system of the Scientific Committee on Problems of the Environment (SCOPE) includes the category “floodplains” but does not distinguish minor sub-units (Gopal et al. 1990). Amazonian várzeas include subunits that can be classed with all riverine and lacustrine and some of the palustrine subunits of the Ramsar Classification System (RCS, 16/11/2011). Our classification system does not pretend to substitute other already existing ones, but to complement them.
We consider large and complex wetlands as landscape units that include a wide variety of functional subunits and major habitats, which interact with each other. Our classification system subdivides the landscape-unit “várzea” in minor subunits according to defined parameters concerning hydrology, water- and soil-chemistry, and plant community characteristics. It is an ecological classification aimed at overcoming the following problems: (1) the lack of fixed boundaries between many habitats, (2) the changes in habitat conditions in the ATTZ throughout the year due to the flood pulse, and (3) the large-scale changes in habitat conditions and the related changes in vegetation cover due to multi-annual changes in discharge and precipitation. It can be used for scientific purposes, such as comparative studies on biomass, productivity, biogeochemical cycles and biodiversity, as well as specific management purposes such as agriculture, husbandry, forestry, fisheries, and protection measures.
Our approach makes use of the habitat types and/or vegetation units already distinguished by the local population, similar to the system devised by Gopal and Sah (1995) in their classification of wetlands of the Indian subcontinent and by Nunes da Cunha and Junk (2011) of the habitats of the Pantanal of Mato Grosso, Brazil. Local denominations of habitats and vegetation units are the result of long-term observations of specific geological, hydrological, and biological characteristics by humans living there, and they provide insight into specific ecosystem functions. Local denominations also facilitate acceptance of a classification system by the local population, which is of fundamental importance in the development and implementation of protection measures. Functional diversity and species diversity are related to habitat diversity, and habitat diversity is threatened by inadequate management strategies. Today’s battle between stake-holders and environmental protection is often not with respect to eco-regions but on a much smaller scale, to habitats and plant communities and their functions in the landscape. As mentioned before, humans have already dramatically changed the vegetation cover of the várzea. The new habitat classification will allow managers to pinpoint threatened habitats and implement specific management and protection measures to reduce negative impacts as well as permit restoration of degraded habitats.
Our classification is open to add additional habitats and habitat characteristics without modifying the entire system or major parts of it. This will certainly occur, because the size of the várzea and the difficulty in accessing some of its regions did not allow us to survey it in its entirety or to study all of its habitats in detail. Climate, hydrology, physical and chemical parameters of soil and water, and communities of higher plants are parameters that can be used for habitat classification in large inland pulsing wetlands worldwide. First, we will extend our classification to other Brazilian river floodplains such as those at Paraná, Araguaia and Guaporé Rivers. This will close a large gap of knowledge about the structure and adequate utilization of Brazilian wetlands. Second, we will analyze, based on available literature, the applicability of our classification systems on wetlands in other parts of the globe. This will facilitate comparison between wetlands and their habitats at a global scale. First attempts of a desk study to transform the very detailed vegetation classification of the Everglades (Rutchey et al. 2006) for direct comparison with the habitats of the Pantanal (Nunes da Cunha and Junk 2011) have been very encouraging.
References
Cowardin LM, Carter V, Golet FC, LaRoe ET (1979) Classification of wetlands and deepwater habitats of the United States. US Department of the Interior, Fish and Wildlife Service, Washington, DC
Diegues ACS (1994) An inventory of Brazilian wetlands. IUCN—The World Conservation Union, Gland
Diegues ACS (2002) Povos e Águas. Núcleo de Apoio à Pesquisa sobre Populações Humanas e Áreas Úmidas Brasileiras, 2nd edn. NUPAUB, São Paulo
Finlayson CM, Van der Valk AG (1995) Wetland classification and inventory: a summary. Vegetatio 118:185–192
Furch K, Junk WJ (1997) Physicochemical conditions in floodplains. In: Junk WJ (ed) The Central Amazon floodplain: ecology of a pulsing system. Ecological studies, vol 126. Springer, Berlin, pp 69–108
Gibbs RJ (1967) The geochemistry of the Amazon River: part I. The factors that control the salinity and the composition and concentration of the suspended solids. Geol Soc Am Bull 78:1203–1232
Gopal B, Sah M (1995) Inventory and classification of wetlands in India. Vegetatio 118:39–48
Gopal B, Kvet J, Löffler H, Masing V, Patten BC (1990) Definition and classification. In: Patten BC (ed) Wetlands and shallow continental water bodies. SPB Academic Publishing, The Hague, pp 9–15
Goulding M, Smith NHJ, Mahar DJ (1996) Floods of fortune: ecology and economy along the Amazon. Columbia University Press, New York
Hoorn C, Wesselingh FP, Ter Steege H, Bermudez MA, Mora A, Sevink J, Sanmartín I, Sanchez-Messeguer A, Anderson CL, Figueiredo JP, Jaramillo C, Riff D, Negri FR, Hooghiemstra H, Lundberg J, Stadler T, Särkinen T, Antonelli A (2010) Amazonian through time: andean uplift, climate change, landscape evolution, and biodiversity. Science 330:927–931
IBGE (2010) Censo Demográfico 2000–2010. Censo. http://www.censo2010.ibge.gov.br/primeiros_dados_divulgados/index.php?uf=13. Accessed 16 Nov 2011
Irion G, Junk WJ, de Mello JASN (1997) The large Central Amazonian river floodplains near Manaus: geological, climatological, hydrological, and geomorphological aspects. In: Junk WJ (ed) The Central Amazon floodplain. Ecology of a pulsing system. Springer, Berlin, pp 23–46
Irion G, de Mello JASN, Morais J, Piedade MTF, Junk WJ, Garming L (2010) Development of the Amazon valley during the middle to late quaternary: sedimentological and climatological observations. In: Junk WJ, Piedade MTF, Wittmann F, Schöngart J, Parolin P (eds) Ecology and management of Amazonian floodplain forests. Ecological studies, vol 210. Springer, Heidelberg, pp 27–42
Junk WJ (1997) Structure and function of the large Central Amazonian River floodplains: synthesis and discussion. In: Junk WJ (ed) The Central Amazon floodplains. Ecology of a pulsing system. Ecological studies, vol 126. Springer, Berlin, pp 455–472
Junk WJ (2005) Flood pulsing and the linkages between terrestrial, aquatic, and wetland systems. Proc. Int. Assoc. Theor. Appl. Limnol. 29(1):11–38
Junk WJ, Piedade MTF (1993) Herbaceous plants of the Amazon floodplain near Manaus: species diversity and adaptations to the flood pulse. Amazoniana 12(3/4):467–484
Junk WJ, Piedade MTF (1997) Plant life in the floodplain with special reference to herbaceous plants. In: Junk WJ (ed) The Central Amazon floodplain: ecology of a pulsing system. Ecological studies, vol 126. Springer, Berlin, pp 147–186
Junk WJ, Wantzen KM (2004) The flood pulse concept: new aspects, approaches, and applications—an update. In: Welcomme RL, Petr T (eds) Proceedings of the 2nd international symposium on the management of large rivers for fisheries, vol 2. Food and Agriculture Organization & Mekong River Commission. FAO Regional Office for Asia and the Pacific, Bangkok, Cambodia, pp 117–149
Junk WJ, Bayley PB, Sparks RE (1989) The flood pulse concept in river-floodplain-systems. Can Special Publ Fish Aquatic Sci 106:110–127
Junk WJ, Ohly JJ, Piedade MTF, Soares MGM (eds) (2000) The central Amazon floodplain: actual use and options for a sustainable management. Backhuys Publishers, Leiden
Junk WJ, Piedade MTF, Schöngart J, Cohn-Haft M, Adeney JM, Wittmann F (2011) A classification of major naturally occurring Amazonian lowland wetlands. Wetlands 31:623–640
Kalliola R, Salo J, Puhakka M, Rajasilta M, Häme T, Neller RJ, Räsänen ME, Arias WAD (1992) Upper Amazon channel migration. Naturwissenschaften 79:75–79
Meade RH (1994) Suspended sediments of the modern Amazon and Orinoco Rivers. Quat Int 21:29–39
Meade RH, Dunne T, Richey JE, Santos UM, Salati E (1985) Storage and remobilization of suspended sediment in the lower Amazon River of Brazil. Science 228:488–490
Melack JM (1984) Amazon floodplain lakes: shape, fetch, and stratification. Verh Internat Verein Limnol 22:1278–1282
Melack JM, Hess LL (2010) Remote sensing of the distribution and extent of wetlands in the Amazon basin. In: Junk WJ, Piedade MTF, Wittmann F, Schöngart J, Parolin P (eds) Ecology and management of Amazonian floodplain forests Ecological studies, vol 210. Springer, Berlin, pp 43–59
Mertes LAK (1994) Rates of floodplain sedimentation on the Central Amazon River. Geology 22:171–174
Mertes LAK, Meade RH (1985) Particle sizes of sands collected from the bed of the Amazon River and its tributaries during 1982–1984. U.S. Geological Survey Open-File Report 85-333, pp 1–16
Nunes da Cunha C, Junk WJ (2011) A preliminary classification of habitats of the Pantanal of Mato Grosso and Mato Grosso do Sul, and its relation to national and international wetland classification systems. In: Junk WJ, da Silva CJ, Nunes da Cunha C, Wantzen KM (eds) The Pantanal: ecology, biodiversity and sustainable management of a large neotropical seasonal wetland. Pensoft, Sofia, pp 127–141
Peixoto JMA, Nelson BW, Wittmann F (2009) Spatial and temporal dynamics of alluvial geomorphology and vegetation in central Amazonian white-water floodplains by remote-sensing techniques. Remote Sens Environ 113:2258–2266
Petermann P (1997) The birds. In: Junk WJ (ed) The Central Amazon floodplain. Ecology of a pulsing system. Ecological studies, vol 126. Springer, Berlin, pp 419–452
Piedade MTF, Junk WJ (2000) Natural herbaceous plant communities in the Amazon floodplain and their use. In: Junk WJ, Ohly JJ, Piedade MTF, Soares MGM (eds) The Central Amazon floodplain: actual use and options for a sustainable management. Backhuys Publishers, Leiden, pp 269–290
Piedade MTF, Junk WJ, Long SP (1991) The productivity of the C4 grass Echinochloa polystachia on the Amazon Floodplain. Ecology 72(4):1456–1463
Piedade MTF, Worbes M, Junk WJ (2001) Geoecological controls on elemental fluxes in communities of higher plants in Amazonian floodplains. In: McClain ME, Victoria RL, Richey JE (eds) The biogeochemistry of the Amazon Basin. Oxford University Press, Oxford, pp 209–234
RCS: http://en.wikipedia.org/wiki/Ramsar_Classification_System_for_Wetland_Type. Accessed 16 Nov 2011
Roosevelt AC (1999) Twelve thousand years of human-environment interaction in the Amazon Floodplain. In: Padoch C, Ayres JM, Pinedo-Vasquez M, Henderson A (eds) Várzea—diversity, development, and conservation of Amazonia’s Whitewater floodplains. Advances in economic botany, vol 13. The New York Botanical Garden Press, New York, pp 371–392
Rutchey K, Schall TN, Doren RF, Atkinson A, Ross M, Jones D, Madden M, Vilchek L, Bradlay KA, Snyder JR, Burch JN, Pernas T, Witcher B, Pyne M, White R, Smith III TJ, Sadle J, Smith CS, Patterson ME, Gann DE (2006) Vegetation classification for South Florida natural areas. United States Geological Survey, open-file report 2006-1240, Saint Petersburg
Schöngart J (2008) Growth-Oriented Logging (GOL): a new concept towards sustainable forest management in Central Amazonian várzea floodplains. For Ecol Manage 256:46–58
Schöngart J, Wittmann F, Worbes M (2010) Biomass and net primary production of Central Amazonian floodplain forests. In: Junk WJ, Piedade MTF, Wittmann F, Schöngart J, Parolin P (eds) Amazonian floodplain forests: ecophysiology, biodiversity and sustainable management Ecological studies, vol 210. Springer, Berlin, pp 347–388
Seidenschwarz F (1986) Pioniervegetation im Amazonasgebiet Perus. Ein pflanzensoziologischer Vergleich von vorandinem Flussufer und Kulturland. Monographs on agriculture and ecology of warmer climates, vol 3. Margraf, Triops Verlag, Langen
Sioli H (1950) Das Wasser im Amazonasgebiet. Forsch Fortschr 26:274–280
Sioli H (1965) Zur Morphologie des Flußbettes des des unteren Amazonas. Naturwissenschaften 52:104
Sousa PT Jr, Piedade MTF, Candotti E (2011) Brazil‘s forest code puts wetlands at risk. Nature 478:458
Sternberg HOR (1960) Radiocarbon dating as applied to a problem of Amazonian morphology. In: XVIII international geographical congress, no. 2, Rio de Janeiro, pp 399–423
Ter Steege H, Pitman N, Phillips OL, Chave J, Sabatier D, Duque A, Molino JF, Prévost MF, Spichiger R, Castellanos H, Hildebrand P, Vásquez R (2006) Continental scale patterns of canopy tree composition and function across Amazonia. Nature 443:444–447
Wittmann F, Oliveira Wittmann A (2010) Use of Amazonian floodplain trees. In: Junk WJ, Piedade MTF, Wittmann F, Schöngart J, Parolin P (eds) Amazonian floodplain forests: ecophysiology, biodiversity and sustainable management. Ecological studies, vol 210. Springer, Berlin, pp 389–418
Wittmann F, Parolin P (2005) Aboveground roots in Amazonian floodplain trees. Biotropica 37:609–619
Wittmann F, Anhuf D, Junk WJ (2002) Tree species distribution and community structure of central Amazonian várzea forests by remote sensing techniques. J Trop Ecol 18:805–820
Wittmann F, Junk WJ, Piedade MTF (2004) The várzea forests in Amazonia: flooding and the highly dynamic geomorphology interact with natural forest succession. For Ecol Manage 196:199–212
Wittmann F, Schöngart J, Montero JC, Motzer T, Junk WJ, Piedade MTF, Queiroz HL, Worbes M (2006) Tree species composition and diversity gradients in white-water forests across the Amazon basin. J Biogeogr 33:1334–1347
Wittmann F, Schöngart J, Junk WJ (2010) Phytogeography, species diversity, community structure and dynamics of Amazonian várzea forests. In: Junk WJ, Piedade MTF, Wittmann F, Schöngart J, Parolin P (eds) Amazonian floodplain forests: ecophysiology, biodiversity and sustainable management. Ecological studies, vol 210. Springer, Berlin, pp 61–102
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Junk, W.J., Piedade, M.T.F., Schöngart, J. et al. A classification of major natural habitats of Amazonian white-water river floodplains (várzeas). Wetlands Ecol Manage 20, 461–475 (2012). https://doi.org/10.1007/s11273-012-9268-0
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11273-012-9268-0