
Abstract
Wetland restoration can mitigate aerobic decomposition of subsided organic soils, as well as re-establish conditions favorable for carbon storage. Rates of carbon storage result from the balance of inputs and losses, both of which are affected by wetland hydrology. We followed the effect of water depth (25 and 55 cm) on the plant community, primary production, and changes in two re-established wetlands in the Sacramento San-Joaquin River Delta, California for 9 years after flooding to determine how relatively small differences in water depth affect carbon storage rates over time. To estimate annual carbon inputs, plant species cover, standing above- and below-ground plant biomass, and annual biomass turnover rates were measured, and allometric biomass models for Schoenoplectus (Scirpus) acutus and Typha spp., the emergent marsh dominants, were developed. As the wetlands developed, environmental factors, including water temperature, depth, and pH were measured. Emergent marsh vegetation colonized the shallow wetland more rapidly than the deeper wetland. This is important to potential carbon storage because emergent marsh vegetation is more productive, and less labile, than submerged and floating vegetation. Primary production of emergent marsh vegetation ranged from 1.3 to 3.2 kg of carbon per square meter annually; and, mid-season standing live biomass represented about half of the annual primary production. Changes in species composition occurred in both submerged and emergent plant communities as the wetlands matured. Water depth, temperature, and pH were lower in areas with emergent marsh vegetation compared to submerged vegetation, all of which, in turn, can affect carbon cycling and storage rates.
Similar content being viewed by others


Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
Wetland restoration is used for a variety of purposes, such as filtering and storing nutrients and organic matter, providing habitat for a variety of species, and re-establishing natural buffer zones for flood control (Zedler 2003). In the Sacramento-San Joaquin Delta, CA, organic soils more than 15 m deep in some areas formed in historic marshes, where organic matter accumulation and land surface elevation gains kept pace with sea level rise during the late Holocene (Atwater 1980). These organic soils were drained for agricultural use beginning in the mid-nineteenth century. Their subsequent compaction and oxidative loss have resulted in extensive subsidence, and some areas more than 6 m below sea level still have several meters of organic material remaining (Deverel and Rojstaczer 1996; Deverel et al. 1998). Continuing subsidence poses an increasing threat to levee stability because it increases the hydraulic gradient and pressures the levees must withstand (Mount and Twiss 2005). Moreover, subsidence endangers this important source of freshwater for about two-thirds of the population of California because catastrophic levee failure in this seismically sensitive region could result in salt water intrusion and contamination for months to years (Mount and Twiss 2005). Restoration of Delta wetlands can mitigate the loss of organic soil from aerobic decomposition, and recreate the conditions that provide for carbon storage (Miller et al. 2000).
Carbon is stored in the environment when primary production exceeds losses. In temperate regions, marshes are highly productive systems (Penfound 1956; Westlake 1963), and prolonged periods of flooding can limit decomposition (Lee 1990; Battle and Golladay 2001), and make them potentially favorable areas for carbon storage (Craft and Richardson 1993; Frockling et al. 2001; Chimner et al. 2002). Flooding duration and depth affect plant species composition and productivity in wetlands (Keddy and Ellis 1985; Casanova and Brock 2000; Fraser and Kernezis 2005). The effect of water depth on plant species distribution and growth provides important information for understanding ecosystem processes such as the biogeochemical cycling of carbon and the potential carbon storage capacity of wetlands (Gosselink and Turner 1978).
To investigate use of permanent shallow flooding to mitigate aerobic decomposition of old soil carbon substrates, and to assess the effect of water depth on carbon storage in restored wetlands, the United States Geological Survey shallowly flooded a deeply subsided agricultural field in the Delta. This study assessed the effects of permanent flooding at two water depths (25 and 55 cm) on plant species composition and productivity to estimate annual carbon inputs to the wetlands for 9 years following flooding. Changes in environmental conditions concomitant with wetland re-establishment that can affect the carbon storage potential of re-established Delta wetlands over time also were studied.
Materials and methods
Site description
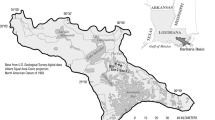
The study site was a 6-ha agricultural field located near the center of Twitchell Island in the Sacramento-San Joaquin River Delta of California (38°6′N, 121°39′W; Fig. 1). The climate is Mediterranean, with cool, rainy winters and hot, dry summers and a growing season that can extend from February to November. The soil is a Rindge mucky silt loam. Botanical analyses of peat deposits in the region showed it primarily comprises Schoenoplectus acutus and Phragmites (Atwater 1980), indicating peat in the delta developed in vast emergent marshes. Approximately 4–6 m of peat underlay the study site.

Location of Twitchell Island in the Sacramento-San Joaquin Delta in California
The field was leveled at ~4.5 m below sea level, and split into two similarly sized areas using a berm. In the fall of 1997, the site was permanently flooded to different water depths (~25 cm in the ‘shallow wetland’ and 55 cm in the ‘deep wetland’) to study the effect of water depth on carbon storage potential in re-established wetlands in the region. To ensure that S. acutus (tules) were able to colonize the restored wetlands, S. acutus stems and rhizomes, collected from stands on the island, were planted in five 10 × 25 m blocks, spaced every 2 m, in the eastern half of both areas shortly before flooding. Typha spp. (cattails) in the surrounding ditches provided windborne seed to the site.
Fresh water was piped in continuously from the San Joaquin River and entered the site through inlets on either end of the south side of the two wetland restoration areas. Outflows were weirs in the center of the north side of each restoration area (Fig. 2). The approximate water depths in each wetland were maintained during the study period by raising outlet weirs to compensate for the accumulation of organic material. Each wetland had six piers, three on the north side and three on the south, projecting into the wetlands to access permanent sampling sites (Fig. 2). To facilitate sampling throughout the wetlands without disturbing the vegetation or substrate, four boardwalks were constructed in each wetland. Boardwalks were used to access permanent transects for assessing plant colonization, growth, spread, and changes in species composition. Allochthonous biomass inputs to the wetlands were not considered because these were essentially closed wetland systems, surrounded by agricultural fields.

Diagram of the wetland site. The west wetland is, and the east wetland is 55 cm deep. Letters represent piers and sampling platforms. Lines represent boardwalks and sampling transects
Transect measurements of plant cover
A point-quadrat method was used to follow temporal changes in species composition of the wetland plant community on four permanent transects ranging from about 100 to 150 m long in each wetland. For each transect sampling event, data were collected from 200 to 300 points and 40 to 53 quadrats in the shallow wetland, and from 150 to 200 points and 30 to 40 quadrats in the deep wetland. There were fewer data points in the deeper wetland because transects in the deep wetland were shorter than those in the shallow wetland. During the first two growing seasons sampling was conducted in June and October (1998 and 1999); thereafter, peak biomass was sampled biennially from 2000 to 2006.
At every 2.5 m interval along each transect algal or plant species presence or absence below the water surface, at the water surface, and above the water surface were recorded. Frequency tables of species presence were compiled to assess changes in percent species cover through time, and by water depth, at different zones (below, at, and above the water surface) in the wetland. Because point transect plant cover sampling provided discrete, categorical data differences in vegetation coverage and species composition were assessed with chi-square analysis of frequency distributions between water depths and over time. The three zones were analyzed both separately and in combination to accurately assess percent cover by the different plant communities.
At every fifth point, or 12.5 m, all live plants growing in a 33 × 33 cm (0.11 m²) quadrat were counted and identified. This provided representative sampling of species composition, spatially, and plant density (Giroux and Bedard 1988). Beginning in 2000, the height, diameter, number of leaves, and flower presence or absence of all emergent vegetation in these sample quadrats were recorded for allometric correlation biomass estimates. Differences in the density of emergent vegetation over time and by water depth were analyzed using SAS general linear model (GLM) and non-parametric comparisons of means, where appropriate.
Allometric biomass models
Because the emergent marsh dominants, S. acutus and Typha spp., contributed the majority of biomass inputs to the wetlands, allometric models to estimate plant biomass were developed by harvesting plants throughout the growing season from both water depths and correlating the measurements of morphologic parameters to plant dry weights. During 1999 and 2001, approximately 800 S. acutus plants of all sizes were collected, and the height of each plant and the diameter at 0.5 m from the top of the shoot base measured. Almost 2,200 Typha spp. plants of all sizes were collected between 2000 and 2002. The tallest leaf and the stem diameter at 0.5 m from the top of the shoot base were measured, the leaves were counted, and flower presence noted. All plants were dried at 70°C for 48–72 h to a constant weight and weighed. The morphologic variables were related to plant biomass measurements, performing transformations of the parameters, as necessary, to meet assumptions of normal distribution and to establish linear relationships between the variables. Plants from the destructive biomass harvests were not used to develop the models. Neither the time of sample collection or water depth affected the allometric relationships.
Destructive biomass harvests in 2000 and 2001 included measurements of plant height, diameter, and leaf number of all individual plants in the samples before they were dried and weighed to allow for comparisons between measured and predicted biomass values. SAS correlation, regression, and GLM analyses were used to test the models by comparing estimated and measured biomass from the destructive biomass harvests.
Measurements of plant biomass
Destructive biomass harvests were used to estimate productivity of the dominant plant species for the first 5 years of the study. Above and belowground biomass of dominant emergent marsh species was measured with destructive harvests in Oct 1998, July and August 1999 and May and July 2000. In July 2001, only aboveground biomass was harvested. In early May, late July and early October 2002, only belowground biomass in the shallow wetland was measured. During 1998 to 2000, samples of floating and submerged vegetation were collected in both wetlands, but no samples of floating and submerged vegetation were collected after 2000, despite changes in species composition in submerged vegetation.
Between 1998 and 2000 all live aboveground emergent marsh vegetation biomass was harvested from 0.11 m² quadrats at ~20 regularly spaced plots along transects in each wetland. In 2001, about 40 samples were collected from each wetland. All belowground structures extracted from two 10 cm diameter, 20 cm deep soil cores from the harvested quadrats were used to estimate belowground biomass (Giroux and Bedard 1988). Soil was washed from the roots, and live roots were separated from dead ones based on color and turgidity. All biomass was dried at 70°C to a constant weight, and weighed. Subsamples of dried shoots and roots were ground with a Tecater mill (1093), ashed at 500°C for 4 h to determine organic matter content, and analyzed for carbon (C) and nitrogen (N) content with a CHNS/O analyzer (Perkin/Elmer series 2 2400).
Beginning in July 2002, destructive sampling of aboveground biomass was replaced with non-destructive methods using data collected in the mid season transect data sampling as a measure of peak season standing live biomass. Plant size measurements from quadrats were used in allometric biomass models developed for this study. The effects of water depth, plant species and sample time on above and belowground biomass measurements were analyzed using SAS GLM.
Seasonal plant growth and mortality turnover assessment
Stem and leaf turnover of the dominant emergent marsh species were estimated using methods based on those of other researchers (Smith and Kadlec 1985; Morris and Haskin 1990; de Leeuw et al. 1996). Ten permanent quadrats were randomly placed in S. acutus stands at each water depth in March 1998 and 1999. S. acutus shoots have a single sheath leaf at the base of the plant, and do not lose and replace leaves over their lifespan. For this reason, annual biomass turnover in S. acutus stands can be reasonably represented by the total number of stems that grow in a given area throughout the year in relation to the number of stems present at the peak of the growing season, and, similarly, by cumulative plant heights or weights. Plant height and number were recorded throughout the growing season to assess the total seasonal growth in a given area. Each plant was tagged and numbered, and its height was measured monthly.
Similarly, in March of 1999 ten random quadrats were placed in Typha stands at each water depth. Typha plants have leaves that are shed as the plant matures, and grow new leaves further up on the shoot. Thus, in Typha stands, turnover of biomass includes both the successive growth and loss of leaves on individual plants, and the successive replacement of old plants by the growth of new plants over a growing season. Turnover of Typha was assessed by tagging, numbering and measuring the length of each leaf on every plant in the quadrat once a month. Flower spikes, and mortality, and final leaf length of newly senesced leaves and plants were also recorded.
The allometric biomass models were used to estimate standing and cumulative biomass from the plant size parameters recorded throughout the growing season for each quadrat. Plant biomass turnover was estimated from growth and mortality through the season by comparing the end of the season cumulative biomass to the mid-season standing biomass mean. Estimates of biomass turnover were compared to turnover estimated by stem density and mortality in both emergent marsh species. Emergent marsh growth and turnover data were analyzed with SAS ANOVA and Mixed Model ANOVA procedures. Annual biomass turnover of aquatic floating and submerged vegetation was not measured.
Annual primary production
Annual C inputs from emergent vegetation to these wetlands were estimated from measurements of mid season standing biomass, annual biomass turnover rate, and average plant biomass C content of shoots and roots. Because root biomass was not measured in all years, root biomass estimates for years when belowground biomass samples were not collected were based on the average root:shoot ratio in relation to annual aboveground production measurements (0.8 ± 0.1), and therefore do not compensate for lower aboveground production with higher aboveground production rates. Also, root turnover in clonal dominants is usually not as high as that of the aboveground vegetation as a result of winter conservation of root biomass for spring growth, in contrast to nearly complete winter die-off of aboveground plant biomass (Garver et al. 1988; Gill and Jackson 2000). Therefore, annual root turnover values were estimated to be half of measured aboveground turnover rates, or 1.5 ± 0.1 times and 2.0 ± 0.1 times mid season live biomass, respectively.
Measurements of environmental parameters
Specific conductance, pH, and dissolved oxygen were measured using YSI probes and meter (556 MPS) at the six piers in each wetland at regular intervals between 1998 and 2004. At each sampling, several water depth measurements were taken to the nearest 0.5 cm, and soil temperatures were measured using permanently placed thermocouple sensors and Omega meters (450 AKT). Discrete measurements of surface water temperatures were made using the Omega thermocouples and meters. Beginning in 2005, water temperatures in the deep wetland were measured continuously in eight locations representing different plant communities, using permanently placed temperature sensors and Hobo data loggers (H8 Outdoor Industrial 4-Channel). At each site, temperature sensors were placed at the bottom of the water column near the wetland sediment surface, midway between the sediment and water surfaces, and floating just beneath the water surface. Statistical analyses of differences in environmental parameters by wetland plant community were conducted using SAS mixed model ANOVA to account for seasonal effects. Relationships among the environmental factors were assessed using SAS correlation analyses.
Results
Plant community composition and cover
Though plant colonization was rapid in both wetlands (Fig. 3a, b), with almost complete cover by the second growing season, emergent marsh vegetation colonization and spread, including planted S. acutus, were slower in the deep wetland than the shallow wetland (χ² < 0.0001). The most rapid spread of emergent marsh vegetation occurred in the first 2 years after flooding, when emergent marsh vegetation colonized nearly 90% of the shallow wetland, and nearly 50% of the deep wetland. In both wetlands, emergent marsh vegetation first colonized to the south and southeast, where winter winds appeared to congregate seeds and vegetative propagules. After the rapid colonization of the first 2 years, cover by emergent marsh vegetation in the deep wetland increased slowly, and the area covered by submerged vegetation decreased.

a Plant community composition in the 25 cm deep wetland based on frequency measures of plant species along transects represented as percent cover from the first year after flooding in September 1997 to July 2006. b Plant community composition in the 55 cm deep wetland based on frequency measures of plant species along transects represented as percent cover from the first year after flooding in September 1997 to July 2006
In the shallow marsh, T. latifolia cover declined from 12% of emergent vegetation in 2000 to 1% of emergent vegetation in 2006. Percent cover of T. angustifolia and T. domingensis varied from year to year, though it tended to be greater for T. angustifolia. In the deep wetland, where emergent vegetation slowly spread throughout the study period, T. latifolia consistently composed 8–9% of the emergent marsh species, while T. angustifolia and T. domingensis showed greater variability. The percentage of Typha with mixed characteristics, indicative of hybridization between the species, slowly increased in both water depths (data not shown).
Following emergent marsh colonization, Lemna (duckweed) and Azolla, small floating aquatic plants, also became established and spread through the wetlands, often under the emergent marsh canopy. However, in the shallow wetland, the area covered by Lemna and Azolla was halved from the maximum cover of 85% recorded in 2000 to the last measurement in 2006; while Lemna cover in the 55 cm wetland was <60% after slowing increasing throughout the study period (data not shown). During this time, the species composition of submerged vegetation also changed as the initial colonizer, Myriophyllum, was replaced by Egeria, Ceratophyllum, and Potamogeton sp. over time (data not shown).
Emergent vegetation tended to grow more densely in the shallow wetland than the deep marsh (Fig. 4; P = 0.0202). Stem density of emergent marsh vegetation showed significant changes with time in the shallow wetland (P < 0.0001), but in the deeper wetland annual differences in plant stand density were not significant (Fig. 4; P > 0.05). Because Typha spp. dominate these wetlands, temporal changes in stem density measurements primarily reflect Typha densities, as the number of samples containing S. acutus was limited, especially in the earlier years. However, stands of S. acutus generally grew more densely than Typha when they occurred (mean stem densities 83.4 ± 12.1 and 53.7 ± 1.7, respectively; P < 0.0001).

Density of emergent marsh vegetation in 25 and 55 cm deep wetlands from the first year after flooding in September 1997 to July 2006. Vertical bars represent standard error
Allometric biomass models
For S. acutus natural log (Ln) transformations of plant height and diameter at 50 cm, showed the strongest linear relationship to log transformed weight variables: log 10 weight = (0.5028 × ln height) + (0.3471 × ln diameter) − 1.7654 (r² = 0.924). Without the inclusion of shoot diameter model estimates were less constrained (log 10 weight = 0.7947 × ln height − 3.2177, r² = 0.824).
Of the 2,200 individual Typha plants collected, about 1,200 were T. latifolia, and 600 were in flower. While biomass of Typha spp. was most strongly correlated to plant height for all plants, the allometric relationships differed depending on Typha species, and whether or not the plant was flowering. T. latifolia plants tended to be heavier than T. angustifolia and T. domingensis plants of the same height, especially when in flower, while T. angustifolia and T. domingensis plants showed similar allometric correlations to one another both with and without flowers. Natural log transformations of the size variables compared to log transformed weights were used to model the correlations because they showed the strongest linear relationships.
Separating plants by species type, where T. latifolia was separated from the other Typha species, and by flowering, SAS GENMOD was used to generate a logistic regression to fit the data: log 10 weight = (−2.188) + (0.601 × ln height) + (0.2128 × ln diameter) + (0.2721 × ln leaf number) − 0.484 (if species is T. angustifolia or T. domingensis) − 0.2677 (if the plant has no flower). The model showed slightly better fit to T. angustifolia or T. domingensis (r² = 0.92) than to T. latifolia (r² = 0.9). The use of leaf number helped to constrain the model, particularly with regard to estimating T. latifolia biomass.
Overall, predicted weights of emergent marsh vegetation were 6% greater than the measured biomass of samples from destructive harvests. Estimates of emergent marsh biomass in the deep water were one percent lower than measured, but were 10% greater than measured biomass in the 25 cm deep water. The correlation coefficient of predicted and measured biomass values from biomass harvests in 2000 and 2001 was 92.4%, with somewhat stronger correlation in the 25 cm wetland (97%) than in the 55 cm deep water (88%). The correlation coefficient from the S. acutus two-parameter model was 93%; and, predicted weights were within 1% of measured values. However, the Typha allocorrelation model tended to overestimate weight compared to measured values by 4.4 ± 0.7%. For Typha, allometric biomass model precision was greater in shallower water depths, but accuracy improved with increasing water depth. Beginning in 2002, aboveground standing biomass estimates for emergent vegetation were based on allometric biomass correlation equations.
Standing plant biomass
Destructive harvests conducted between 1998 and 2000 showed the ash-free dry weight of standing live aboveground emergent marsh vegetation was an order of magnitude greater than that of standing crop biomass measurements of submerged and floating vegetation in the wetlands (P < 0.0001; Table 1). Emergent vegetation also had significantly higher C:N ratios than aquatic vegetation in the wetlands (P < 0.0001).
Mid-season standing aboveground biomass of emergent marsh vegetation ranged from 924 to 2,353 g m−² (Fig. 5). Measurements of standing aboveground biomass of emergent marsh vegetation showed significant inter-annual variability (P < 0.0001), but no significant difference between the deep and shallow wetlands (P > 0.05).

Measurements of peak standing live above- and below-ground biomass of emergent marsh vegetation in the 25 and 55 cm deep wetlands from 1998 to 2006. Vertical bars represent standard error
Root biomass measurements also showed significant inter-annual variation (Fig. 5; P = 0.0022), but did not show differences by water depth when both wetlands were sampled (P > 0.05). However, S. acutus root biomass was greater than root biomass in Typha stands (P = 0.0018). Root:shoot ratios also showed significant inter-annual differences (P = 0.0322), but did not significantly differ by emergent marsh species (S. acutus = 0.9 ± 0.1; Typha = 0.8 ± 0.1; P > 0.05) or water depth (deep wetland = 0.7 ± 0.1; shallow wetland = 0.9 ± 0.1; P > 0.05). The lowest root:shoot ratios were measured in 1999 (0.7 ± 0.1), and highest in 2000 (1.2 ± 0.3). Root:shoot ratios varied more widely in S. acutus (0.6 ± 0.2 to 1.7 ± 0.4) than in Typha stands (0.7 ± 0.1 to 1.0 ± 0.3), and appeared to respond more to differences in water depth than Typha (1.0 ± 0.2, deep and 0.7 ± 0.2, shallow in S. acutus; 0.8 ± 0.1 at both water depths in Typha; P > 0.5). There were significant inter-annual differences in ash-free root C:N ratios (P = 0.0002), but no significant differences between emergent marsh species. In both root and shoot samples, ash-free N content was highest in the first growing season, which resulted in significantly lower plant tissue C:N ratios in 1998 than all other years of the study (data not shown).
In 2002, live S. acutus root biomass was greatest in the October sample, when dead root biomass was lower than in May and July samples (Fig. 6). Live Typha root biomass did not show seasonal changes, but mid-season Typha root samples from the July harvest had a large amount of adventitious root growth compared to other sample dates, and the lowest ash-free root N concentration (0.87 ± 0.06) and highest dead root biomass of the season. In both emergent marsh species, ash-free root N concentrations were highest in the May sample (P < 0.0001; when S. acutus = 1.5 ± 0.13 and Typha = 1.28 ± 0.12). S. acutus roots had greater ash (0.096 ± 0.004; P = 0.0051) and ash-free N content (1.21 ± 0.06; P = 0.0171) than Typha roots (ash content = 0.085 ± 0.0003; N = 1.03 ± 0.06). Similarly, ash-free root C content was significantly greater in S. acutus (45.6 ± 0.2) than Typha (44.2 ± 0.1; P < 0.0001).

Live and dead root biomass of Typha and Scoenoplectus acutus in the shallow wetland in the 2002 growing season. Vertical bars represent standard error
Plant biomass turnover
Assessments of plant turnover based on both stem number and biomass did not differ significantly from one another in S. acutus stands, nor did the measures of turnover show significant effects of water depth. Annual stem turnover from quadrats in both water depths averaged 1.9 ± 0.1 times the mid season live stem number in 1998 and 1.7 ± 0.1 times in 1999; and, estimated annual biomass turnover was 1.9 ± 0.1 times the mid-season standing biomass measurement. These measurements indicated that standing biomass measurements from mid-season sampling represent a little more than half of S. acutus annual production (Fig. 7).

Cumulative seasonal biomass and live standing biomass of Typha and Schoenoplectus acutus in 1999. Vertical bars represent standard error
Water depth also did not significantly affect turnover estimates of Typha, so average annual Typha turnover measurements were estimated using quadrats from both water depths. Annual turnover of Typha stems was 1.6 ± 0.01 times the mid season live stem number. Stem turnover under represented Typha production because it did not include leaf turnover that occurred with stem growth. Annual Typha turnover based on biomass estimates was 2.1 ± 0.1 (Fig. 7), and did not differ significantly from turnover estimates based on measurements of leaf length production over the course of a year. These turnover measurements showed that mid season standing live Typha biomass represented a little less than half of the annual biomass production of Typha. Plant emergence and mortality, or annual stem turnover, represented ~30% of biomass turnover, and the shedding and replacement of leaves by Typha accounted for about 25% of seasonal biomass.
Estimates of annual biomass turnover of Typha and S. acutus did not differ significantly, and were pooled to estimate an average annual turnover of emergent marsh vegetation in these wetlands of 2.0 ± 1.0 times mid season standing live biomass. Therefore, on average, mid season aboveground standing biomass in these marshes represented about half of their total annual biomass production. Root turnover was not measured.
While Typha spp. and S. acutus did not show significant differences in mid-season standing biomass or seasonal turnover rate, Typha spp. senesced earlier in the year than S. acutus. So at the end of the growing season, S. acutus had significantly greater live standing biomass than Typha spp. (P > 0.0001; Fig. 7).
Annual primary production
Estimates of annual C inputs from emergent vegetation showed large inter-annual variation, ranging from 1.2 kg to more than 3 kg m−² (Table 2), with average annual inputs >2 kg C m−2. Water depth did not significantly affect the C inputs from stands of established emergent marsh vegetation, but it did affect the percent cover of emergent marsh vegetation (Table 2).
Wetland environment
The emergent and submerged plant communities showed significant differences in wetland environmental parameters. Water under emergent marsh vegetation had significantly lower pH than in areas of submerged vegetation and algae (Fig. 8; P = 0.0347). Dissolved oxygen concentrations in water also showed significant differences by plant community (P = 0.0187; data not shown), and were strongly correlated to pH (correlation coefficient = 0.7689; P < 0.0001). Also, areas of emergent marsh vegetation had lower water temperatures compared to areas of aquatic vegetation with no plant canopy shading the water surface (Fig. 9; P = 0.0252).

Water pH in emergent and submerged vegetation from April 1999 to October 2002. Vertical bars represent standard error

Average surface and subsurface water temperatures recorded in half hour intervals in four emergent and four submerged vegetation sites in the 55 cm deep wetland between 30 April 2006 and 2 May 2006
Discussion
Because biomass estimates from emergent vegetation were much greater than inputs from submerged and floating vegetation, the emergent wetland plant community dominated C inputs to these wetlands. Biomass estimates for submerged and floating aquatic vegetation at our site were comparable to estimates of standing biomass of these aquatic plants in other studies (Adams and McCracken 1974; Best and Visser 1986; Ennabili et al. 1998; Squires and Lesack 2003; Carillo et al. 2006). However, C inputs from submerged and floating aquatic vegetation made up a relatively small and fairly labile proportion of the carbon entering the system, as indicated by their low C:N ratios (Taylor et al. 1989; Szumigalski and Bayley 1996).
Carbon inputs from emergent marsh plants are significantly greater than inputs from submerged and floating aquatic vegetation, and decompose more slowly than submerged plant species (Rodgers, et al. 1983), which may be important to peat development in warm climates (Bridgham and Richardson 1992). Measurements of plant species cover in the different depth wetlands indicate areas with differences in both inputs and potential losses; and, therefore, are important to estimates of potential C storage in these wetlands. In this study, estimates of C inputs from plant biomass contributing to C storage comprise C from emergent vegetation only because the highly labile aquatic vegetation was unlikely to play an important role in wetland C storage.
Standing biomass measurements of emergent marsh vegetation in these wetlands displayed large inter-annual variability but were comparable to standing biomass measurements for Typha and Schoenoplectus in other wetlands (Klopatek and Stearns 1978; Brinson et al. 1981; Smith and Kadlec 1985; Davis 1990; Tanner 1994). Large inter-annual variation in standing biomass measurements is common in emergent marsh communities (Pearsall and Gorham 1956; Morris and Haskin 1990; Neill 1990). Productive years can leave a large amount of standing dead plant material that may impose light limitations on the following year’s production, which can significantly affect plant growth dynamics (Garbey et al. 2006; Rocha et al. 2008).
Standing crop biomass measurements can severely underestimate net primary production because they do not include seasonal biomass lost to leaf and shoot mortality prior to sampling, or growth occurring afterwards (Bradbury and Hofstra 1976; Kirby and Gosselink 1976; Linthurst and Reimold 1978a; Whigham et al. 1978). This unmeasured production can be substantial in regions with long growing seasons (Pratolongo et al. 2005).
Measurements of emergent marsh biomass turnover in these marshes showed that annual primary production was approximately twice mid season standing live biomass. This is comparable to biomass turnover measurements in other emergent marshes, though measurements of seasonal plant biomass turnover show differences according to plant species, environment, length of growing season, and year of measurement (Kirby and Gosselink 1976; Linthurst and Reimold 1978b; Davis 1990; Morris and Haskin 1990; de Leeuw et al. 1996). In some plant species, such as T. domingensis and Spartina alterniflora, higher turnover occurs at times and locations with greater nutrient availability (Kirby and Gosselink 1976; Davis 1990). However, S. alterniflora can also show lower biomass turnover in years with greater peak standing biomass production (Morris and Haskin 1990), indicating that a variety of factors can influence plant biomass turnover rates. In these wetlands, biomass turnover estimates of S. acutus were similar between the two sampling years, even though significant differences in plant tissue nitrogen concentrations suggest high nitrogen availability during the first growing season after flooding.
In a fertilization study significant differences in aboveground biomass of S. acutus among treatments were balanced by shifting root:shoot ratios, so that overall biomass production was not significantly higher with fertilization (Neill 1990). Similarly, in the restored wetlands in this study, the total plant (root and shoot) biomass was very similar in years when both were measured, despite significant inter-annual differences in both shoot and root biomass. Root:shoot ratios of S. acutus during in the first two growing seasons after flooding were similar to the low root:shoot ratios in S. validus found in high nutrient treatments in agricultural wastewaters, but increased significantly in 2000. Typha root:shoot ratios in these wetlands did not show significant inter-annual variation.
Factors that potentially affect root production, root:shoot ratios, or plant biomass allocation can be important to C storage in marshes because belowground biomass composes a large fraction of the organic matter that is preserved in wetlands, indicating that belowground production is important to rates of C storage in wetlands (Chen and Twilley 1999; Middleton and McKee 2001). Belowground structures are not exported from the wetland, can decompose more slowly, and are less subject to herbivory than aboveground biomass (van der Valk and Attiwill 1984). The importance of roots to organic matter accumulation in wetlands primarily is attributed to slow decomposition, which can be caused by a variety of factors, including oxygen limitations, a build up of toxic metabolites in the soil, and lower tissue quality of roots compared to aboveground tissues (Middleton and McKee 2001; Chimner and Ewel 2005).
Water depth can affect biomass allocation between roots and shoots in some plant species (Karagatzides and Hutchinson 1991; Squires and van der Valk 1992). A study in Manitoba found that S. lacustrus spp. allocated more biomass to roots in both the shallower and deeper ends of their water depth ranges, but T. glauca did not (Squires and van der Valk 1992). However, in deeper water T. angustifolia was found to produce thicker rhizomes (Sharma et al. 2008), and a larger proportion of vertical rhizomes than in shallower water (Asaeda et al. 2008). Allometric investigation of biomass allocation above and belowground in T. domingensis and Cladium jamaicense showed no response to nutrient gradients, but did show size-dependent biomass allocation such that smaller plants had greater root biomass relative to larger plants (Miao et al. 2008). In another study, S. fluviatilis showed greater biomass allocation to roots in smaller plants, but relatively constant root:shoot ratios in T. latifolia at all sizes (Whigham and Simpson 1978).
In this study, both belowground production and stem density were higher in S. acutus than in Typha. Also, plant density was higher in the shallower wetland than the deeper wetland, though overall aboveground biomass estimates did not significantly differ by water depth. In permanently flooded conditions, T. glauca and S. lacustrus spp. glaucus, demonstrated the ability to increase shoot length with increasing water depth, which compensated for decreasing plant densities with increasing water depth so that standing biomass was relatively constant (Squires and van der Valk 1992; Waters and Shay 1992).
Spatial and temporal variation in vegetative cover in natural systems requires a large number of samples to adequately assess biomass inputs (Dickerman et al. 1986; Thursby et al. 2002); but, allometric relationships between size and weight of plants allow primary production to be assessed with rapid, non-destructive sampling of a site (Morris and Haskin 1990; Thursby et al. 2002). This is particularly useful in marshes dominated by a few species of clonal emergent macrophytes. Typha allocorrelation models developed for this study showed the northern and southern cattails, T. angustifolia and T. domingensis, respectively, had more similar morphological relationships to biomass than T. latifolia, which is more widely distributed. Also, S. acutus tended to have taller shoots and a longer growing season than Typha spp. The continuing spread of S. acutus into areas previously colonized by Typha spp. may be related to the differences in seasonal growth among these marsh species.
Typha latifolia cover declined with time in the shallower marsh, but continued to make up a fairly constant proportion of the Typha community in the deep wetland, where emergent marsh vegetation continues to slowly colonize and spread. T. latifolia appears to be an early colonizer, well adapted to changing environments, while T. angustifolia and T. domingensis demonstrate greater stress tolerance (McNaughton 1975; Grace 1989). Also, T. angustifolia and T. domingensis hybrids are fertile, unlike hybrids with T. latifolia (Smith 1987).
Changes in wetland plant composition can affect the wetland environment in a variety of ways that can affect rates of C sequestration. Litter from different plant species decomposes at different rates (Davis and van der Valk 1978), and emergent marsh vegetation tends to decompose more slowly than submerged or floating vegetation. Vegetation can affect nutrient flows through wetlands (Simpson et al. 1978), which can affect rates of both growth and decomposition (Hietz 1992; Squires and Lesack 2003; Corstanje et al. 2006). Furthermore, this study showed differences in the wetland environment, associated with different plant communities, which could have important effects on biogeochemical processes.
Compared to areas with submerged vegetation and algae, emergent marsh vegetation was associated with lower water temperatures and pH, both of which can slow decomposition (Lee 1990; Murayama and Bakar 1996), and increase C storage, proportionately. Shading by emergent marsh vegetation appeared to decrease water temperatures, and to inhibit photosynthetic activity by submerged vegetation and algae. Photosynthesis by submerged vegetation and algae increases the oxygen concentration in water, which, in turn, can increase pH (Reddy 1981; Gordon and Sand-Jensen 1990). Increases in sediment elevation, or decreases in water depth, can be attributed, in part, to belowground plant biomass inputs in marshes (Reed 2002). Also, the presence of emergent marsh vegetation can slow water movement by increasing friction, which can increase sediment deposition, and, subsequently, decrease water depth (Brueske and Barrett 1994; Harter and Mitsch 2003).
Significant elevation gains from organic matter accumulation only occurred in emergent marsh areas of these wetlands; and, within emergent marsh areas, measurements of accretion varied significantly both between wetlands and by location within the wetlands (Miller et al. 2008), unlike measurements of emergent marsh primary production. Organic matter accumulation results from the balance of inputs and losses. Therefore, environments that minimize decomposition can be important to achieving maximum C storage rates in wetlands, and not high productivity alone.
Primary productivity in wetlands averages twice that of other terrestrial ecosystems (Schlesinger 1997), and this long-term study demonstrated that re-established marshes in the Sacramento-San Joaquin Delta can have annual C inputs averaging more than 2 kg C m−2 year−1. These estimates fall in the high end of the range of wetland production measurements (Mitsch and Gosselink 1993; Sharitz and Phillips 2006), showing that these wetlands are highly productive. Despite inter-annual variability, the high rate of primary productivity in these marshes indicates the potential for good carbon storage capacity.
The study also demonstrated that water depth affected plant establishment and spread, and that differences in environmental parameters occurred between emergent and submerged plant communities. This can affect many wetland functions, including C cycling. Changes in the wetland environment concomitant with colonization by emergent marsh vegetation, such as lower water temperatures, pH, and dissolved oxygen concentrations, suggest that not only do emergent marsh plants provide large annual C inputs, but the colonization and spread of the emergent marsh plant community have associated changes in the wetland environment that could slow decomposition, and, thereby, facilitate C storage in re-established Delta marshes.
References
Adams MS, McCracken MD (1974) Seasonal production of the Myriophyllum component of the littoral of Lake Wingra, Wisconsin. J Ecol 62(2):457–465. doi:10.2307/2258991
Asaeda T, Sharma P, Rajapakse L (2008) Seasonal patterns of carbohydrate translocation and synthesis of structural carbon components in Typha Angustifolia. Hydrobiologia 607:87–101. doi:10.1007/s10750-008-9369-1
Atwater BF (1980) Attempts to correlate the late quaternary climatic records between San Francisco Bay, the Sacramento-San Joaquin Delta, and the Mokelomne. Ph.D. dissertation, University of Delaware
Battle JM, Golladay SW (2001) Hydroperiod influence on breakdown of leaf litter in cypress-gum wetlands. Am Midl Nat 146:128–145. doi:10.1674/0003-0031(2001)146[0128:HIOBOL]2.0.CO;2
Best EPH, Visser HWC (1986) Seasonal growth of the submerged macrophyte Ceratophyllum demersum L. in mesotrophic Lake Vechen in relation to isolation, temperature and reserve carbohydrates. Hydrobiologia 148(3):231–243. doi:10.1007/BF00017526
Bradbury IK, Hofstra G (1976) Vegetation death and its importance to primary production measurements. Ecology 57:209–211. doi:10.2307/1936414
Bridgham SD, Richardson CJ (1992) Mechanisms controlling soil respiration (CO2 and CH4) in southern peatlands. Soil Biol Biochem 24(11):1089–1099. doi:10.1016/0038-0717(92)90058-6
Brinson MM, Lugo AE, Brown S (1981) Primary productivity, decomposition, and consumer activity in freshwater wetlands. Annu Rev Ecol Syst 12:123–161. doi:10.1146/annurev.es.12.110181.001011
Brueske CC, Barrett CW (1994) Effects of vegetation and hydrologic load on sedimentation patterns in experimental wetland systems. Ecol Eng 3:429–447. doi:10.1016/0925-8574(94)00011-5
Carillo Y, Guarin A, Guillot G (2006) Biomass distribution, growth and decay of Egeria densa in a tropical high-mountain reservoir (NEUSA, Columbia). Aquat Bot 85:7–15. doi:10.1016/j.aquabot.2006.01.006
Casanova MT, Brock MA (2000) How do depth, duration and frequency of flooding influence the establishment of wetland plant communities? Plant Ecol 147:237–250. doi:10.1023/A:1009875226637
Chen R, Twilley RR (1999) A simulation model of organic matter and nutrient accumulation in mangrove wetland soils. Biogeochemistry 44:93–118
Chimner RA, Ewel KC (2005) A tropical freshwater wetland: II. Production, decomposition, and peat formation. Wetlands Ecol Manag 13:671–684. doi:10.1007/s11273-005-0965-9
Chimner RA, Cooper DJ, Parton WJ (2002) Modeling carbon accumulation in Rocky Mountain fens. Wetlands 22(1):100–110. doi:10.1672/0277-5212(2002)022[0100:MCAIRM]2.0.CO;2
Corstanje R, Reddy KR, Portier KM (2006) Typha latifolia and Cladium jamaicense litter decay in response to exogenous nutrient enrichment. Aquat Bot 84:70–78. doi:10.1016/j.aquabot.2005.07.013
Craft CB, Richardson CJ (1993) Peat accretion and N, P and organic C accumulation in nutrient-enriched and unenriched Everglades peatlands. Ecol Appl 3(3):446–458. doi:10.2307/1941914
Davis SM (1990) Sawgrass and cattail production in relation to nutrient supply in the Everglades. In: Sharitz RR, Gibbons JW (eds) Freshwater wetlands and wildlife. Office of Scientific and Technical Information, U.S. Department of Energy, Oak Ridge, pp 325–341
Davis CB, van der Valk AG (1978) The decomposition of standing and fallen litter of Typha glauca and Scirpus fluviatilis. Can J Bot 56:662–674. doi:10.1139/b78-073
de Leeuw J, Wielemaker A, de Munck W, Herman PM (1996) Net aerial primary production (NAPP) of the marsh macrophyte Scirpus maritimus estimated by a combination of destructive and non-destructive sampling methods. Hydrobiologia 123:101–108
Deverel SJ, Rojstaczer S (1996) Subsidence of agricultural lands in the Sacramento-San Joaquin Delta, California: role of aqueous and gaseous carbon fluxes. Water Resour Res 32:2359–2367. doi:10.1029/96WR01338
Deverel SJ, Wang B, Rojstaczer S (1998) Subsidence of organic soils, Sacramento-San Joaquin Delta, CA. In: Borchers JW (ed) Land subsidence histories and current research. Proceedings of the Dr. Joseph F. Poland symposium. Association of Engineering Geologist special publication no. 8. Star Publishing Co., Belmont, pp 489–502
Dickerman JA, Stewart AA, Wetzel RG (1986) Estimates of net annual aboveground production: sensitivity to sampling frequency. Ecology 67(3):650–659. doi:10.2307/1937689
Ennabili A, Ater M, Radoux M (1998) Biomass production and NPK retention in macrophytes from wetlands of the Tingitan Peninsula. Aquat Bot 62:45–56. doi:10.1016/S0304-3770(98)00075-8
Fraser LH, Kernezis JP (2005) A comparative assessment of seedling survival and biomass accumulation for fourteen different wetland plant species grown under minor water-depth differences. Wetlands 25(3):520–530. doi:10.1672/0277-5212(2005)025[0520:ACAOSS]2.0.CO;2
Frockling S, Roulet NT, Moore TR, Richard PJH, Richard PJH, Lavoie M, Muller SD (2001) Modeling northern peatland decomposition and peat accumulation. Ecosystems (N Y, Print) 4(5):479–498. doi:10.1007/s10021-001-0105-1
Garbey C, Thiebaut G, Muller S (2006) An experimental study of the plastic responses of Ranunuculus peltatus Schrank to four environmental parameters. Hydrobiologia 570:41–46. doi:10.1007/s10750-006-0159-3
Garver FG, Dubbe DR, Pratt DC (1988) Seasonal patterns in accumulation and partitioning of biomass and macronutrients in Typha spp. Aquat Bot 32:115–127. doi:10.1016/0304-3770(88)90092-7
Gill RA, Jackson RB (2000) Global patterns of root turnover for terrestrial ecosystems. New Phytol 147:3–31. doi:10.1046/j.1469-8137.2000.00676.x
Giroux JF, Bedard J (1988) Estimating above- and below-ground macrophyte production in Scirpus tidal marshes. Can J Bot 66:368–374. doi:10.1139/b88-059
Gordon DM, Sand-Jensen K (1990) Effects of O2, pH and DIC on net-O2 evolution by marine macroalgae. Mar Biol (Berl) 106:445–451. doi:10.1007/BF01344325
Gosselink JG, Turner RE (1978) The role of hydrology in freshwater wetland ecosystems. In: Good RE, Whigham DF, Simpson RL (eds) Freshwater wetlands: ecological processes and management potential. Academic Press, New York
Grace JB (1989) Effects of water depth on Typha latifolia and Typha domingensis. Am J Bot 76(5):762–768. doi:10.2307/2444423
Harter SK, Mitsch WJ (2003) Patterns of short-term sedimentation in a freshwater created marsh. J Environ Qual 32:325–334
Hietz P (1992) Decomposition and nutrient dynamics of reed (Phragmites austalis (Cav.) Trin. Ex Steud.) litter in Lake Neusiedl, Austria. Aquat Bot 43:211–230. doi:10.1016/0304-3770(92)90068-T
Karagatzides JD, Hutchinson I (1991) Intraspecific comparisons of biomass dynamics in Scirpus americanus and Scirpus maritimus on the Fraser River Delta. J Ecol 79:459–476. doi:10.2307/2260726
Keddy PA, Ellis TH (1985) Seedling recruitment of 11 wetland plant species along a water level gradient: shared or distinct responses? Can J Bot 63:1876–1879
Kirby CJ, Gosselink JG (1976) Primary production in a Louisiana gulf coast Spartina alterniflora marsh. Ecology 57:1052–1059. doi:10.2307/1941070
Klopatek JM, Stearns FW (1978) Primary productivity of emergent marsh macrophytes in a Wisconsin freshwater marsh ecosystem. Am Midl Nat 100(2):320–332. doi:10.2307/2424831
Lee SY (1990) Net aerial primary productivity, litter production and decomposition of the reed Phragmites communis in a nature reserve in Hong Kong: management implications. Mar Ecol Prog Ser 66:161–173. doi:10.3354/meps066161
Linthurst RA, Reimold RJ (1978a) An evaluation of methods for estimating net aerial primary productivity of estuarine angiosperms. J Appl Ecol 15:919–931. doi:10.2307/2402787
Linthurst RA, Reimold RJ (1978b) Estimated net aerial primary productivity for selected estuarine angiosperms in Maine, Delaware, and Georgia. Ecology 59(5):945–955. doi:10.2307/1938546
McNaughton SJ (1975) r- and K-selection in Typha. Am Nat 109:251–262. doi:10.1086/282995
Miao S, Sindhoj E, Edelstein C (2008) Allometric relationships of field populations of two clonal species with contrasting life histories, Cladium jamaicense and Typha domingensis. Aquat Bot 88:1–9. doi:10.1016/j.aquabot.2007.08.001
Middleton BA, McKee KL (2001) Degradation of mangrove tissues and implications for peat formation in Belizean island forests. J Ecol 89:818–828. doi:10.1046/j.0022-0477.2001.00602.x
Miller RL, Hastings L, Fujii R (2000) Hydrologic treatments affect gaseous carbon soil loss from organic soils, Twitchell Island, California, October 1995–December 1997. US Geological Survey, water resources investigations report, Sacramento, CA, 00-4042
Miller RL, Fram M, Fujii R, Wheeler G (2008) Subsidence reversal in a re-established wetland in the Sacramento-San Joaquin Delta, CA, USA. San Francisco Estuary and Water Science 6(3) article 1. Available from: http://repositories.cdlib.org/jmie/sfews/vol6/iss3/art1
Mitsch WJ, Gosselink JG (1993) Wetlands, 2nd edn. Van Nostrand Reinold, New York
Morris JT, Haskin B (1990) A 5-yr record of aerial primary production and stand characteristics of Spartina alterniflora. Ecology 71(6):2209–2217. doi:10.2307/1938633
Mount J, Twiss R (2005) Subsidence, sea level rise, and seismicity in the Sacramento-San Joaquin Delta. San Fran Estuary Watershed Sci 3(1). Article 5
Murayama S, Bakar ZA (1996) Decomposition of tropical peat soils 1. Decomposition kinetics of organic matter of peat soils. Jpn Agric Res Q 30:145–151
Neill C (1990) Nutrient limitation of hardstem bulrush (Scirpus acutus Muhl.) in a Manitoba interlake region marsh. Wetlands 10(1):69–76
Pearsall WH, Gorham E (1956) Production ecology I. Standing crops of natural vegetation. Oikos 7(11):193–201
Penfound WT (1956) Primary production of vascular aquatic plants. Limnol Oceanogr 1(2):92–101
Pratolongo P, Vicari R, Kandus P, Malvarez I (2005) A new method for evaluating net aboveground primary production (NAPP) of Scirpus gigantus (Kunth). Wetlands 25(1):228–232. doi:10.1672/0277-5212(2005)025[0228:ANMFEN]2.0.CO;2
Reddy KR (1981) Diel variations of certain physico-chemical parameters of water in selected aquatic systems. Hydrobiologia 85:201–207. doi:10.1007/BF00017610
Reed DJ (2002) Understanding tidal marsh sedimentation in the Sacramento-San Joaquin Delta, California. J Coast Res 36(special issue):605–611
Rocha AV, Potts DL, Goulden ML (2008) Standing litter as a driver of interannual CO2 exchange variability in a freshwater marsh. J Geophys Res 113:G04020. doi:10.1029/2008JG00713.2008
Rodgers JH Jr, Mckevitt ME, Hammerlund DO, Dickson KL, Cairns J Jr (1983) Primary production and decomposition of submerged and emergent aquatic plants of two Appalacian rivers. In: Fontaine TDIII, Bartell SM (eds) Dynamics of lotic ecosystems. Ann Arbor Science, Ann Arbor
Schlesinger WH (1997) Biogeochemistry: an analysis of global change, 2nd edn. Academic Press, San Diego
Sharitz RR, Phillips SC (2006) Development of wetland plant communities. In: Batzer PB, Sharitz RR (eds) Ecology of freshwater and estuarine wetlands. University of California Press, Berkeley
Sharma P, Asaeda T, Fujino T (2008) Effect of water depth on the rhizome dynamics of T. angustifolia. Wetlands Ecol Manag 16:43–49. doi:10.1007/s11273-007-9055-5
Simpson RL, Whigham DF, Walker R (1978) Seasonal patterns of nutrient movement in a freshwater tidal marsh. In: Good RE, Whigham DF, Simpson RL (eds) Freshwater wetlands: ecological processes and management potential. Academic Press, New York, pp 3–20
Smith SG (1987) Typha: its taxonomy and the ecological significance of hybrids. Arch Hydrobiol 27:29–138
Smith LM, Kadlec JA (1985) Fire and herbivory in a Great Salt Lake marsh. Ecology 66(1):259–265. doi:10.2307/1941326
Squires MM, Lesack LFW (2003) The relation between sediment nutrient content and macrophyte biomass and community structure along a water transparency gradient among lakes of the Mackenkie Delta. Can J Fish Aquat Sci 60:333–343. doi:10.1139/f03-027
Squires L, van der Valk AG (1992) Water-depth tolerances of the dominant emergent macrophytes of the Delta Marsh, Manitoba. Can J Bot 70:1860–1867. doi:10.1139/b92-230
Szumigalski AR, Bayley SE (1996) Decomposition along a bog to rich fen gradient in central Alberta, Canada. Can J Bot 74:573–581
Tanner CC (1994) Growth and nutrition of Schoenoplectus validus in agricultural wastewaters. Aquat Bot 47:131–153. doi:10.1016/0304-3770(94)90010-8
Taylor BR, Parkinson D, Parsons WFJ (1989) Nitrogen and lignin content as predictors of litter decay rates: a microcosm test. Ecology 70(1):97–104. doi:10.2307/1938416
Thursby GB, Chintala MM, Stetson D, Wigland C, Champlin DM (2002) A rapid, non-destructive method for estimating aboveground biomass of salt marsh grasses. Wetlands 22(3):626–630. doi:10.1672/0277-5212(2002)022[0626:ARNDMF]2.0.CO;2
van der Valk AG, Attiwill PM (1984) Decomposition of leaf and root litter of Avicennia marina at Westernport Bay, Victoria, Australia. Aquat Bot 18:205–221. doi:10.1016/0304-3770(84)90062-7
Waters I, Shay JM (1992) Effect of water depth on population parameters of a Typha glauca stand. Can J Bot 70:349–351. doi:10.1139/b92-046
Westlake DF (1963) Comparisons of plant productivity. Biol Rev Camb Philos Soc 38:385–425. doi:10.1111/j.1469-185X.1963.tb00788.x
Whigham DF, Simpson RL (1978) The relationship between aboveground and belowground biomass of freshwater tidal macrophytes. Aquat Bot 5:355–364. doi:10.1016/0304-3770(78)90076-1
Whigham DF, McCormick J, Good RE, Simpson RL (1978) Biomass and primary production in freshwater tidal wetlands of the middle Atlantic coast. In: Good RE, Whigham DF, Simpson RL (eds) Freshwater wetlands: ecological processes and management potential. Academic Press, New York, pp 3–20
Zedler JB (2003) Wetlands at your service: reducing impacts of agriculture at the watershed scale. Front Ecol Environ 1(2):65–72
Acknowledgments
We owe great thanks to the California Department of Water Resources for long-term funding of this long-term research project. And, we would like to specially thank Lauren Hastings for her hard work getting the project started. Also, thanks to the many people who helped with data collection and site maintenance and repair on this study over all the years. Finally, thanks to Allison Brown, Lisa Marie Windham Myers, and the reviewers for Wetlands, Ecology, and Management for their helpful comments and suggestions for this manuscript. It is all greatly appreciated.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Miller, R.L., Fujii, R. Plant community, primary productivity, and environmental conditions following wetland re-establishment in the Sacramento-San Joaquin Delta, California. Wetlands Ecol Manage 18, 1–16 (2010). https://doi.org/10.1007/s11273-009-9143-9
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11273-009-9143-9