
Abstract
Tropical forests play an important role in carbon cycle. However, the temporal and spatial variation in soil carbon dioxide (CO2) emission of tropical forest remains uncertain, especially near the Tropic of Cancer. In this research, we studied the annual soil CO2 fluxes from three tropical montane rainforests on the Hainan Island of China (pristine montane rainforest, PF; secondary montane rainforest, SF; and Podocarpus imbricatus plantation, PP). The results showed a lower annual average soil CO2 flux as 6.85 ± 0.52 Mg C-CO2 ha−1 (9.17 Mg C-CO2 ha−1 in the wet season and 4.50 Mg C-CO2 ha−1 in the dry season). The CO2 fluxes exhibited obviously seasonal variation during the study period. Among the three forest types, PF had the highest average CO2 flux rate of 317.77 ± 147.71 mg CO2 m−2 h−1 (433.08 mg CO2 m−2 h−1 in the wet season and 202.47 mg CO2 m−2 h−1 in the dry season), followed by PP of 286.84 ± 137.48 mg CO2 m−2 h−1 (367.12 mg CO2 m−2 h−1 in the wet season and 206.56 mg CO2 m−2 h−1 in the dry season) and SF of 255.09 ± 155.26 mg CO2 m−2 h−1 (351.48 mg CO2 m−2 h−1 in the wet season and 155.71 mg CO2 m−2 h−1 in the dry season). We found between CO2 fluxes and soil temperature a highly significant linear relation (P < 0.01) at 5 cm depth and a highly significant exponential correlation (P < 0.01) at 10 cm depth for all three forest types; a significant linear relation (P < 0.05) between CO2 fluxes and soil moisture content was found for SF and PF, but not for PP (P > 0.05). The CO2 flux was significantly correlated (P < 0.05) with water-filled pore space only for PF. In conclusion, our results suggested soil CO2 fluxes in the three forest types that exhibit obviously spatial and temporal variation, and the temperature is the major factor affecting soil CO2 fluxes from this region.
Similar content being viewed by others


Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
1 Introduction
The global terrestrial carbon (C) sink was estimated as 2.7 Pg C year−1 from 2000 to 2006, equivalent to about 30 % of total anthropogenic emissions during this period (Canadell et al. 2007). Soil organic carbon (SOC) releases into the atmosphere as carbon dioxide (CO2), mineralized, and decomposed by soil microorganisms and soil enzymes. Soil respiration accounts for much of the CO2 released from terrestrial ecosystems into the atmosphere (Bujalský et al. 2014). CO2 is the main greenhouse gas that accounts for 70 % of the total effect of these gases on current climate change (IPCC 2013). Soil CO2 flux is the largest component of forest CO2 flux, accounting for 40 to 70 % of total forest respiration (Itoh et al. 2012; Ohkubo et al. 2007). Under the background of increasing CO2 concentration in the atmosphere and the related potential change in climate, soil CO2 emission research is done to estimate future atmospheric CO2 concentration and global change (Liang 2002).
Soil CO2 emission, commonly referred to as soil respiration, is the sum of decomposition of organic matter, tiny animals, microbial respiration, and root respiration (Schlesinger and Andrews 2000). Hence, the temporal and spatial variability of soil respiration is affected by soil physical, chemical, and biological properties. Among them, soil moisture content and temperature directly affect roots and microbial activities and, at the same time, indirectly influences soil physical-chemical properties (Davidson et al. 1998). Besides, atmospheric carbon dioxide (CO2) concentration and nitrogen (N) deposition may interactively impact soil respiration in terrestrial ecosystems (Deng et al. 2013). Soil CO2 fluxes were also found to vary with vegetation species (Adachi et al. 2006; Akburak and Makineci 2013; Huang et al. 2014; Katayama et al. 2009; Nottingham et al. 2012; Soe and Buchmann 2005) and the degree of mineralization (Cardoso et al. 2013; Silvola et al. 1996; Wood et al. 2012).
Tropical forests, which are among the largest terrestrial reservoirs of C, store more than 40 % of the global C in vegetation. They contain most abundant terrestrial biome and the greatest global source of net primary production (NPP) and, therefore, are important contributors to global carbon cycling and play an important role in regulating global climate (Osuri et al. 2014). Though it is acknowledged that the release of C from tropical forests might exacerbate future climate change, the magnitude of the effect on climate remains uncertain (Cox et al. 2013). In particular, the spatial and temporal variations in soil respiration and its relationship with biophysical factors in forests near the Tropic of Cancer are not clearly understood (Tang et al. 2006a). Although a great deal of studies have investigated soil CO2 fluxes in tropical forests, most focused on those in Amazonia and Southeast Asia (Davidson et al. 2008; Itoh et al. 2012; Jauhiainen et al. 2005; Koehler et al. 2009; Kosugi et al. 2007; Wangluk et al. 2013; Zanchi et al. 2014), while very few explored East Asian forests. For Chinese tropical forests, several studies were about the soil CO2 fluxes in Xishuangbanna (Dong et al. 2012; Sha et al. 2005; Wang et al. 2004; Werner et al. 2006; Zhou et al. 2008) and in Hainan (Luo et al. 2001; Wu et al. 1997; Zhou et al. 2013). Therefore, this research aimed to: (1) understand the temporal and spatial variations of soil CO2 fluxes in the main tropical montane rainforest types of China and (2) to determine the influencing factors on soil CO2 fluxes from these tropical forests.
2 Materials and Methods
2.1 The Study Site
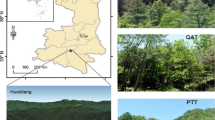
The study site is located in Jianfengling National Natural Reserve (18° 23′–18° 52′ N, 108° 36′–109° 05′ E) on the southwestern Hainan Island, China. The total area of the reserve is approximately 470 km2. This part of Hainan Island has a tropical monsoon climate with obvious wet season from May to October and dry season from November to April. The annual mean temperature is 19.8 °C ranging from 14.8 to 23.3 °C, the annual mean rainfall 2449 mm, and yearly mean relative humidity (RH) 88 %. Rainfalls during the wet season account for over 80 % of the annual total (Bai et al. 2014). The montane rainforests cover about 163 km2 (Chen et al. 2010), with several major forest types including pristine and secondary montane rainforest and plantations. In the present study, we chose on an altitude of 800–900 m three experiment sites representing the three typical local forests, namely the pristine montane rainforest (PF), secondary montane rainforest (SF) and Podocarpus imbricatus plantation (PP) (Fig. 1). Among the three, SF and PP were nearly cutover in 1960s to 1970s, and PP reforested with native trees including P. imbricatus. In PF, the typical trees are Gironniera subaequalis, Litsea baviensis, Cryptocarya chinensis, Altingia chinensis, Livistona saribus, Castanopsis hystrix, and Mallotus hookerianus. In SF, the predominant trees are Brassaia actinophylla, Castanopsis fissa, Endospermum chinense, Cyclobalanopsis glauca, Adinandra hainanensis, and Castanopsis Jianfenglingsis. In PP, the particular trees are Sapium discolor, P. imbricatus, and Dacrydium pierrei. There is no record of fertilization in any forest (Bai et al. 2014; Li et al. 2002).

Map of Jianfengling National Natural Reserve in Southwest Hainan Province, South China, showing the location and appearance of the primary mountain rainforest (PF), secondary mountain rainforest (SF), and Podocarpus imbricatus plantation (PP) forest types
2.2 Sampling Site Setting and Gas Flux Measurement
For each forest type, six static chambers randomly placed. A distance of at least 10 m was kept between any two chambers in each forest. The chambers were made of polyvinylchloride (PVC) pipe and made up by a fixed collar and a chamber with 20 cm internal diameter and 40 cm height. The collar of static chamber was permanently installed in the earth at a depth of about 7 cm, as described by Bai et al (2014).
Soil CO2 flux was measured fortnightly from July 2013 to June 2014. For each sampling, gas samples were collected with 10 ml one-off vacuum tubes (without additives) from 8:30 a.m. to 12:30 p.m., four tubes each time with 10-min intervals. Therefore, for each sampling, we collected all together 72 tubes of samples. The inner air temperature of the chamber was recorded at 0 and 30 min of each hour. The CO2 concentration of these samples was measured using a gas chromatograph (Agilent 7890A, Agilent Co., USA), equipped with an electron capture detector (ECD), and operated at 350 °C. The flux F of CO2 quantified with mg CO2 m−2 h−1 was calculated as:
Where P is the atmospheric pressure of the sampling site (Pa); M is the molar mass of CO2 (g mol−1); d c /d t is the rate of concentration change; T is the absolute temperature at sampling time (oC); V 0 , P 0 , and T 0 are the molar volume, atmospheric pressure, and absolute temperature, respectively (mL, Pa, and °C), under standard conditions; and H is the chamber height over the soil surface (cm). According to the flux Eq. (1), the annual mean CO2 flux in Mg CO2 ha−1 year−1 was determined based on 24 sampling times with 18 data of CO2 fluxes from the three forests each time.
2.3 Soil Characteristics
Once every 3 months from July 2013 to June 2014, 5 soil samples were randomly collected from 0 to 10 cm soil depth within 25 cm distance around each chamber, after the leaf litter was cleared off the ground. The 5 soil samples were then mixed into a composite sample for the chamber. The samples were transported to the laboratory of Chengdu Institute of Biology, Chinese Academy of Sciences with a portable ice box. Then each composite sample was passed through a 2-mm sieve to remove all visible sundries, such as litter fragments, stones, coarse roots, and small soil animals and stored at 4 °C to determine soil physicochemical properties including dissolved organic carbon (DOC) and pH. The DOC was measured with the continuous flow analytical system (SKALAR San++, SKALAR Co., Netherlands). The soil pH was measured using a pH meter (PB-10, Sartorius Instruments Inc., Germany; soil/water = 1:2.5). With each sampling, we used a handheld digital thermometer (plug-in thermometer, Cixiguanghua digital instruments, Ningbo) to measure soil temperature at depths of 5 cm (T5) and 10 cm (T10) within 5 cm of each chamber and a kerosene thermometer to test the temperature inside chamber (Tc). Meanwhile, soil moisture content (SM) was measured with a soil moisture meter (Delta-T HH2 Read 2.7-ML2X, UK) and was converted into volume percentage. SM could be described as water filled pore space % (WFPS) which was calculated by the equation:
Where Vol is volumetric water content (%), SBD is soil bulk density (g cm−3), and 2.65 is the density of quartz (g cm−3) (Bai et al. 2014). The test frequency of WFPS, pH, and DOC is identical with the soil sampling frequency, once every 3 months, but different from the twice-a-month frequency of gas sample collecting and measuring for SM, T10, T5, and Tc.
2.4 Statistical Analysis
One-way analysis of variance (ANOVA) was used to compare gas fluxes among three forest types and to examine the differences in soil moisture, temperature, pH, WFPS, and DOC. To correlate T10, T5, Tc, SM, and WFPS with CO2 flux, we used regression analysis. The difference among the three types was tested using the LSD method. All statistical analyses were done with SPSS 19.0, and significance was set at P < 0.05. All figures were plotted with Origin 8.5.
3 Results
3.1 Soil Temperature, Temperature Inside Chamber, Soil Moisture, and WFPS in the Three Forest Types
In this research, 10 cm soil temperature (T10), 5 cm soil temperature (T5), and temperature inside chamber (Tc) showed significant seasonal variation in all three forest types (Fig. 2). The mean Tc, T5, and T10 for three forest types were all higher during the wet season than during the dry season (Table 1), exhibiting a maximum in May and a minimum in December (Fig. 2). The mean T10 (mean ± SD) was 19.45 ± 3.32 °C for PF, 19.87 ± 2.98 °C for SF, and 19.83 ± 3.01 °C for PP respectively. The mean T5 was 19.43 ± 3.35 °C for PF, 19.84 ± 2.97 °C for SF, and 19.73 ± 2.95 °C for PP respectively. The mean Tc was 20.68 ± 3.77 °C for PF, 20.64 ± 3.04 °C for SF, and 20.80 ± 3.24 °C for PP respectively. Among the three forest types, the differences of Tc were not significant (P > 0.05). At both 5 and 10 cm soil depths, the soil temperature was significantly different between SF and PF and between PP and PF (P < 0.01), but not between SF and PP (P > 0.05).

Soil (at depths of 5 and 10 cm) and chamber air temperature from July 2013 to June 2014 from the three forest types
The SM also exhibited seasonal differences during this study period. For three forest types, the mean SM during the wet season was relatively higher than that during dry season (Table 1). The SM value (%, mean ± SD) was 21.99 ± 4.84 for PF, 25.98 ± 4.66 for SF, and 29.48 ± 5.83 for PP and the SM range was 11.07–28.89 %, 11.65–31.62 %, and 17.08–40.52 % (for PF, SF, and PP, respectively). The mean WFPS also presented highly seasonal variation (Table 1), being (%, mean ± SD) 57.95 ± 13.71 for PF, 67.90 ± 11.31 for SF, and 72.60 ± 11.88 for PP, and the WFPS range was 31.95–83.35 %, 44.68–85.50 %, and 49.09–96.11 % (for PF, SF, and PP, respectively). Among the three forest types, both SM and WFPS showed significant differences, with the mean SM and WFPS both in the order of PP > SF > PF. Other soil properties were indicated in Table 2.
3.2 CO2 Fluxes from the Three Forest Types
The average ± SD of CO2 flux was 317.77 ± 147.71 mg CO2 m−2 h−1 for PF, 255.09 ± 155.26 mg CO2 m−2 h−1 for SF, and 286.84 ± 137.48 mg CO2 m−2 h−1 for PP respectively. The mean CO2 flux of PF was markedly higher than that of SF (P < 0.05), but there was no significant difference between those of PF and PP or between those of SF and PP (P > 0.05, Fig. 3a). In each forest type, the soil CO2 fluxes were significantly higher during the wet season than during the dry season (P < 0.01, Fig. 3b–d), with a coefficient of variation (CV) as 51.76 %. The highest CO2 flux rate (727.15 mg CO2 m−2 h−1) was from SF and lowest rate (64.72 mg CO2 m−2 h−1) from PF; for all three forest types, the maximum fluxes all occurred in July 2013 (585.98 mg CO2 m−2 h−1 for PF, 727.15 mg CO2 m−2 h−1 for SF, and 514.23 mg CO2 m−2 h−1 for PP); and the minimum fluxes occurred during the dry season of December 2013 or January 2014 (64.72 mg CO2 m−2 h−1 for PF, 65.08 mg CO2 m−2 h−1 for SF, and 67.56 mg CO2 m−2 h−1 for PP). The CV was 46.48 % for PF, 47.93 % for SF, and 60.86 % for PP. The extent of CO2 fluxes variation was markedly greater in SF than of the other two forest types (P < 0.05). For all three forest types, the CO2 fluxes exhibited a significant seasonal variation, with the fluctuation of CO2 fluxes greater during wet season than during dry season (Fig. 4).

Annual mean soil CO2 emissions (a) and mean soil CO2 emissions in the wet and dry seasons (b, c, d) from the three forest types. LSD test, error bars mean ± 1 SD (n = 3). WS and DS indicate wet season and dry season respectively. Different letters indicate significant differences among the forest types for each period, P < 0.05 for lowercases and P < 0.01 for uppercases

Seasonal pattern of soil CO2 emission in primary mountain rainforest (PF), secondary mountain rainforest (SF), and Podocarpus imbricatus plantation (PP). Error bars indicate ±1 SD (n = 3). WS and DS indicate wet season and dry season, respectively
3.3 Relation Between the Environmental Factors and CO2 Fluxes
For all three forest types, the average CO2 flux was positively related to T10 (R 2 = 0.661, P < 0.01 for PF; R 2 = 0.404, P < 0.01 for SF; R 2 = 0.315, P < 0.01 for PP) and T5 (R 2 = 0.545, P < 0.01 for PF; R 2 = 0.242, P < 0.01 for SF; R 2 = 0.346, P < 0.01 for PP) (Fig. 5). The optimal temperature for CO2 fluxes was 22–23 °C at both 5 and 10 cm depths for all three forest types (Fig. 5). Similarly, CO2 fluxes were also highly significantly related to Tc for PF (R 2 = 0.395, P < 0.01) and for PP (R 2 = 0.311, P < 0.01) and positively related for SF (R 2 = 0.181, P < 0.05) (Table 3).

Linear relationship between soil temperature (at depths of 5 and 10 cm) and soil CO2 emission in PF, SF, and PP. (P < 0.05 for SF and P < 0.01 for PF and PP)
CO2 fluxes were not significantly related to SM for PP (P > 0.05), but positively related for SF (R 2 = 0.194, P < 0.05) and highly significantly related for PF (R 2 = 0.298, P < 0.01) (Table 3). The optimum SM range for CO2 fluxes was 20–30 %.
The WFPS was highly correlated with CO2 fluxes only for PF (R 2 = 0.127, P < 0.05), but not for SF or PP (P > 0.05 for both) (Table 3). The optimum WFPS range for CO2 fluxes was 55–80 %.
A negative correlation was observed between mean soil density and CO2 fluxes, and it was no clear difference among all three forest types (Table 2). Soil pH value in PF was higher than that in SF (P < 0.05) and PP (P > 0.05). Furthermore, significant differences in the DOC content were found among the three forest types soils as PF > SF > PP (P < 0.05) (Table 2).
4 Discussion
4.1 CO2 Fluxes from the Tropic Montane Forests
In this study, we recorded a mean CO2 emission rate of 286.57 ± 146.82 mg CO2 m−2 h−1, equivalent to a yearly mean CO2 emission rate of 6.85 ± 3.35 Mg C-CO2 ha−1 year−1, within the range of soil respiration (6.53–23.72 Mg C ha−1 year−1) (Bond-Lamberty and Thomson 2010) in tropical montane forests, but relatively lower than the 15.92 Mg C ha−1 year−1 reported by Zhou et al (2013) for the same region. In comparison with results for other forest types in the world (Table 4), our result was relatively higher than those from tropical rainforests in Xishuangbanna (4.11 and 3.71 Mg C ha−1 year−1, respectively) (Dong et al. 2012; Werner et al. 2006) and in Thailand (3.20 Mg C ha−1 year−1) (Wangluk et al. 2013), but lower than that for tropical forests in Indonesia (11.49 Mg C ha−1year−1) (Jauhiainen et al. 2005), Malaysia (15.23 Mg C ha−1year−1) (Kosugi et al. 2007), Brazil (12.80 Mg C ha−1year−1) (Davidson et al. 2008), and America (9.37 Mg C ha−1year−1) (Koehler et al. 2009). Since several studies had suggested that nitrogen deposition would reduce the soil C emission and promote ecosystem C accumulation (Deng et al. 2010; Fan et al. 2014; Mo et al. 2005; Xu et al. 2009), we considered the lower nitrogen deposition rate in Jianfengling (6.1 Kg N ha−1year−1) (Bai et al. 2014) than that in Xishuangbanna (9.0 Kg N ha−1year−1) (Sha et al. 2005) as the main reason of the relatively higher CO2 emission rate in this study.
4.2 Variations of CO2 fluxes Among the Three Forest Types
There were significantly spatial and temporal disparities in CO2 fluxes from the three forest types (Fig. 4), with the average CO2 emission rates significantly higher in PF than in SF (P < 0.05) (Fig. 3a), but only minor differences between SF and PP, PP, and PF. The result was consistent with previous reports (Dong et al. 2012; Werner et al. 2006). The major causes of spatial and temporal variation in soil CO2 emission from tropical forests have been widely studied (Adachi et al. 2006; Cardoso et al. 2013; Katayama et al. 2009; Kosugi et al. 2007; Rowlings et al. 2012; Soe and Buchmann 2005). On the one hand, tree species may substantially alter the soil source or sink strength for greenhouse gases through root related processes (Fender et al. 2013), and microbial processes play a central role in the global fluxes of the key biogenic CO2 and are likely to respond rapidly to climate change (Singh et al. 2010). On the other hand, other abiotic factors like temperature and moisture also regulate CO2 production and emission by influencing decomposition of these incorporated organic inputs (Hassan et al. 2014). In addition, for some rainforest ecosystems, soil texture and drainage capacity constraints are more important flux controlling factors than vegetation and seasonal variability (Rowlings et al. 2012). Soil texture also plays an important role in determining the tipping point of the positive effect of soil moisture on soil CO2 efflux in tropical soils (Wood et al. 2013). Nevertheless, CO2 emission is such an extremely complicated process that we have to incorporate the actual situation in consideration (Schlesinger and Andrews 2000). For instance, according to Nottingham et al (2012), there was no significant difference in soil CO2 fluxes between the two tropical montane cloud forest systems in the lowland tropical forest and on the garden lawn, but in our study, CO2 fluxes were significant higher in PF than in SF (P < 0.05). Shifts in tree species composition would likely provoke the difference of soil CO2 flux (Díaz-Pinés et al. 2014; Katayama et al. 2009; Osuri et al. 2014). From the point of view of ecological system, we considered such difference of soil CO2 flux was mainly due to the significant differences in environmental conditions including different species composition and biomasses (Chen et al. 2010; Li et al. 2002), pH, soil temperature, soil moisture, WFPS, and DOC only between SF and PF (Table 2). Although species composition and biomasses were considerably higher in PF and SF than in PP, there were similar environmental factors (Table 2), and no significant difference between PF and PP, SF, and PP (P > 0.05) (Fig. 3), which suggested species composition and biomasses are not limiting factors for soil CO2 emissions in this region.
Besides, CO2 emissions also showed significant seasonal differences in all forest types (P < 0.01), with relatively higher CO2 emission rates during the wet season than during the dry season (Fig. 3b–d), similar to many previous results (Akburak and Makineci 2013). This is probably because of the seasonal variation of environmental factors (Table 1). In other words, the seasonality of soil respiration coincides with the seasonal climate pattern, with high respiration rates in the hot humid season and low rates in the cool dry season (Itoh et al. 2012; Tang et al. 2006b). Moreover, we observed an interesting phenomenon that the CO2 flux showed significant fluctuations during the wet season, from July 2013 to December 2013 and from May 2014 to June 2014 (P < 0.01), but only slight fluctuations during the dry season, from November 2013 to April 2014 (P > 0.05) (Fig. 4). This is probably because higher temperature and sufficient soil water content during the wet season could supply appropriate circumstance for soil microorganisms and all biochemical processes (Yan et al. 2014; Zhou et al. 2014), which led to greater changes of CO2 fluxes during the wet season (Fig. 4). For all three forest types, the peak of CO2 flux occurred in July during the wet season (Fig. 4), probably because of the optimal temperature through the month (Figs. 2 and 5).
4.3 Effects of Soil Properties on CO2 Flux
Soil temperature showed significant seasonal variation (Table 1, Fig. 2), and it was indicated as the major influencing factor of CO2 fluxes by ANOVA (Fig. 5). Studies reported that soil temperature among numerous environmental factors plays an important role to affect soil CO2 emissions (Han et al. 2007; Tang et al. 2006b; Wood et al. 2013; Zanchi et al. 2014). A report found that soil CO2 emissions significantly correlated with soil temperature (R 2 = 0.87, P < 0.01) in the pine forest (Cheng et al. 2013). There was a high exponential relationship between soil temperature and soil respiration whose seasonal variation was mainly determined by soil temperature and substrate availability (Deng et al. 2013; Zhou et al. 2013). Our study found a highly significant linear relationship between CO2 fluxes and soil temperature (T5) for PF, SF, and PP (P < 0.01). More interestingly, we also found a significant exponential correlation between CO2 fluxes and soil temperature (T10) among the three forest types (P < 0.01). It might imply that the CO2 production process from 10-cm depth soil and 5-cm depth soil are a little different. For all three forest types, the optimal temperature for soil CO2 emissions at both 5 and 10-cm depths was 20–23 °C (Fig. 5). Moreover, the result of our research suggested that the temperature inside chamber is an important factor to influence soil CO2 fluxes (Table 3). Although our study site is located in the tropic region, its relatively higher altitude may lead to larger temperature variation than other tropic rainforests. As a result, the marked seasonal variation in soil and atmosphere temperature may interactively influence soil CO2 fluxes.
Different from soil temperature, soil moisture content showed no significant effects on soil CO2 fluxes for PP (P > 0.05), but a significant linear relation with CO2 fluxes from PF (P < 0.01) and SF (P < 0.05) (Table 3), which might be related with the wider range of SM for PP than that for PF and SF (Table 2). Several studies reported SM as a major factor influencing seasonal variation of soil respiration (Hashimoto et al. 2004; Itoh et al. 2012; Zanchi et al. 2014). In our study, SM was an important but not limiting factor for soil CO2 emissions because most emitted CO2 is produced in top 10–20 cm soil where there is low water content (Subke et al. 2003; Yan et al. 2014).
A linear relation existed between WFPS and CO2 fluxes only for PF (P < 0.01), but not for SF or PP (P > 0.05) (Table 3). Such relation pattern could be explained by differences in soil texture (Wood et al. 2013). In previous studies, CO2 emission rates were reported to be positively correlated to changes in WFPS at dry to moderate soil water contents during the dry season, but negatively correlated to changes in WFPS during the wet season (Kiese and Butterbach-Bahl 2002). Moreover, the reduced WFPS of the saturated soil might decrease the diffusion of CO2 out of the soil, leading to lower CO2 emissions (Liptzin et al. 2011; Wood et al. 2013). In addition, altered precipitation strongly influences soil respiration through the well-known direct effects of soil moisture on root and microbial activities, and by modification of both moisture and temperature sensitivity of soil respiration (Jiang et al. 2013), because oxygen availability is a main factor controlling the reaction rates and temperature sensitivity of CO2 production (Blagodatskaya et al. 2014); on the other hand, saturated soils may also limit the diffusion of O2 into the soil, which could create anaerobic conditions that limit the production of CO2 (Silver et al. 1999; Wood et al. 2013). In other words, excessive moisture would affect soil permeability and then affect the activity of soil microorganisms and enzymes, ultimately resulting in reduced CO2 emissions (Davidson et al. 1998; Hanson et al. 2000). During the wet season from July 2013 to June 2014 at Jianfengling, the rainfall was close to 2000 mm and the soil moisture for most time ranged from 20 to 30 %, especially relatively higher for PP (40.52 %) (Table 2). Therefore, in our study, soil moisture did not play a major role in affecting soil CO2 fluxes in tropical montane rainforest, which was consistent with Zhou et al (Zhou et al. 2013). However, it was also reported that the positive effect of soil temperature on CO2 emissions is constrained by soil moisture availability (Wood et al. 2013). Soil moisture may have a complicated interaction with soil temperature and together influence soil respiration. For instance, with the same temperature rise, wetter soils may emit more CO2 into the atmosphere via heterotrophic respiration (Zhou et al. 2014). The complicated interaction between soil temperature and soil moisture in our study (higher temperature and higher SM during the wet seasons than that during the dry seasons) needs further research.
Other soil properties (such as pH, DOC, N–NH4 +, N–NO3 −, and mean soil density) all directly or indirectly affect soil CO2 emission rate (Mosier 1998; Schlesinger and Andrews 2000). For instance, DOC can be used as an indicator of carbon availability to soil microorganisms (Boyer and Groffman 1996). The spatial variation of CO2 emission rate may be related to the complex forest structures and abundant species diversity (Cook et al. 2014; Katayama et al. 2009; Matvienko et al. 2014), as well as complicated CO2 production processes (Hassan et al. 2014; van Groenigen et al. 2011). In our research, the study site was disturbed by constant tropical storms and typhoon during the wet season, resulting in more fresh litter input to the forest floor, which caused a mass of C substrates in soil organic matter and thus regulate soil respiration (Zhou et al. 2013). Among the three forest types in our study, PF was found to have the lowest soil temperature (at both depths of 5 and 10 cm) and soil moisture, but the highest DOC content and the highest CO2 emission rate. It might be because DOC content is one of the important factors influencing soil CO2 flux and can explain about 23.8–33.6 % of soil CO2 flux variability determined by parent materials (Lou et al. 2004). Of course, the relationship between CO2 emission and DOC needs further studying.
5 Conclusions
The results of the present study showed a high temporal variation of CO2 fluxes in the three tropical montane rainforest types, with distinctly higher CO2 fluxes during wet season for all three forest types than during dry season. The seasonal variation is especially obvious for SF where CO2 flux in wet season is twice that in dry season, which is probably due to soil temperature and substrate availability. The annual mean CO2 emission rate also shows spatial variation, being higher in PF than the other two forest types, which indicates pristine forest soil have much higher CO2 emission potential than other forest types in this region. The spatial differences might be caused by differences in vegetation structure and soil environment among the three forests. In all forest types, soil CO2 emission rate is strongly correlated with soil temperature, but not with SM and WFPS. In addition, other soil properties (pH, DOC, mean soil density, and soil total porosity) show no significant effect on soil CO2 fluxes. Soil temperature is considered the major factor affecting soil CO2 fluxes from the tropical montane rainforests on Hainan Island, China.
References
Adachi, M., Bekku, Y. S., Rashidah, W., Okuda, T., & Koizumi, H. (2006). Differences in soil respiration between different tropical ecosystems. Applied Soil Ecology, 34, 258–265.
Akburak, S., & Makineci, E. (2013). Temporal changes of soil respiration under different tree species. Environmental Monitoring and Assessment, 185, 3349–3358.
Bai, Z. Z., Yang, G., Chen, H., Zhu, Q. V., Chen, D. X., Li, Y. D., Wang, X., Wu, Z. M., Zhou, G. Y., & Peng, C. H. (2014). Nitrous oxide fluxes from three forest types of the tropical mountain rainforests on Hainan Island, China. Atmospheric Environment, 92, 469–477.
Blagodatskaya, E., Zheng, X., Blagodatsky, S., Wiegl, R., Dannenmann, M., & Butterbach-Bahl, K. (2014). Oxygen and substrate availability interactively control the temperature sensitivity of CO2 and N2O emission from soil. Biology and Fertility of Soils, 50, 775–783.
Bond-Lamberty, B., & Thomson, A. (2010). A global database of soil respiration data. Biogeosciences, 7, 1915–1926.
Boyer, J. N., & Groffman, P. M. (1996). Bioavailability of water extractable organic carbon fractions in forest and agricultural soil profiles. Soil Biology and Biochemistry, 28, 783–790.
Bujalský, L., Kaneda, S., Dvorščík, P., & Frouz, J. (2014). In situ soil respiration at reclaimed and unreclaimed post-mining sites: responses to temperature and reclamation treatment. Ecological Engineering, 68, 53–59.
Canadell, J. G., Le Quéré, C., Raupach, M. R., Field, C. B., Buitenhuis, E. T., Ciais, P., Conway, T. J., Gillett, N. P., Houghton, R., & Marland, G. (2007). Contributions to accelerating atmospheric CO2 growth from economic activity, carbon intensity, and efficiency of natural sinks. Proceedings of the National Academy of Sciences, 104, 18866–18870.
Cardoso, S. J., Vidal, L. O., Mendonca, R. F., Tranvik, L. J., Sobek, S., & Roland, F. (2013). Spatial variation of sediment mineralization supports differential CO2 emissions from a tropical hydroelectric reservoir. Frontiers in Microbiology, 4, 8.
Chen, D. X., Li, Y. D., Liu, H. P., Xu, H., Xiao, W. F., Luo, T. S., Zhou, Z., & Lin, M. X. (2010). Biomass and carbon dynamics of a tropical mountain rain forest in China. Science China. Life Sciences, 53, 798–810.
Cheng, J. Z., Lee, X. Q., Zhou, Z. H., Wang, B., Xing, Y., Cheng, H. G., & Tang, Y. (2013). The effects of litter layer and soil properties on the soil-atmosphere fluxes of greenhouse gases in Karst Forest, Southwest China. Polish Journal of Ecology, 61, 79–92.
Cook, R. L., Binkley, D., Mendes, J. C. T., & Stape, J. L. (2014). Soil carbon stocks and forest biomass following conversion of pasture to broadleaf and conifer plantations in southeastern Brazil. Forest Ecology and Management, 324, 37–45.
Cox, P. M., Pearson, D., Booth, B. B., Friedlingstein, P., Huntingford, C., Jones, C. D., & Luke, C. M. (2013). Sensitivity of tropical carbon to climate change constrained by carbon dioxide variability. Nature, 494, 341–344.
Davidson, E., Belk, E., & Boone, R. D. (1998). Soil water content and temperature as independent or confounded factors controlling soil respiration in a temperate mixed hardwood forest. Global Change Biology, 4, 217–227.
Davidson, E. A., Nepstad, D. C., Ishida, F. Y., & Brando, P. M. (2008). Effects of an experimental drought and recovery on soil emissions of carbon dioxide, methane, nitrous oxide, and nitric oxide in a moist tropical forest. Global Change Biology, 14, 2582–2590.
Deng, Q., Zhou, G., Liu, J., Liu, S., Duan, H., & Zhang, D. (2010). Responses of soil respiration to elevated carbon dioxide and nitrogen addition in young subtropical forest ecosystems in China. Biogeosciences, 7, 315–328.
Deng, Q., Cheng, X., Zhou, G., Liu, J., Liu, S., Zhang, Q., & Zhang, D. (2013). Seasonal responses of soil respiration to elevated CO2 and N addition in young subtropical forest ecosystems in southern China. Ecological Engineering, 61, 65–73.
Díaz-Pinés, E., Schindlbacher, A., Godino, M., Kitzler, B., Jandl, R., Zechmeister-Boltenstern, S., & Rubio, A. (2014). Effects of tree species composition on the CO2 and N2O efflux of a Mediterranean mountain forest soil. Plant and Soil, 384, 243–257.
Dong, L. Y., Wu, C. S., Gao, J. M., & Sha, L. Q. (2012). Effects of simulated rainfall on the soil respiration in tropical secondary forest and rubber plantation in Xishuangbanna of Yunnan, Southwest China. Chinese Journal of Ecology, 31, 1887–1892.
Fan, H. B., Wu, J. P., Liu, W. F., Yuan, Y. H., Huang, R. Z., Liao, Y. C., & Li, Y. Y. (2014). Nitrogen deposition promotes ecosystem carbon accumulation by reducing soil carbon emission in a subtropical forest. Plant and Soil, 379, 361–371.
Fender, A.-C., Gansert, D., Jungkunst, H. F., Fiedler, S., Beyer, F., Schützenmeister, K., Thiele, B., Valtanen, K., Polle, A., & Leuschner, C. (2013). Root-induced tree species effects on the source/sink strength for greenhouse gases (CH4, N2O and CO2) of a temperate deciduous forest soil. Soil Biology and Biochemistry, 57, 587–597.
Han, G., Zhou, G., Xu, Z., Yang, Y., Liu, J., & Shi, K. (2007). Soil temperature and biotic factors drive the seasonal variation of soil respiration in a maize (Zea mays L.) agricultural ecosystem. Plant and Soil, 291, 15–26.
Hanson, P., Edwards, N., Garten, C., & Andrews, J. (2000). Separating root and soil microbial contributions to soil respiration: a review of methods and observations. Biogeochemistry, 48, 115–146.
Hashimoto, S., Tanaka, N., Suzuki, M., Inoue, A., Takizawa, H., Kosaka, I., Tanaka, K., Tantasirin, C., & Tangtham, N. (2004). Soil respiration and soil CO2 concentration in a tropical forest, Thailand. Journal of Forest Research, 9, 75–79.
Hassan, W., David, J., & Abbas, F. (2014). Effect of type and quality of two contrasting plant residues on CO2 emission potential of Ultisol soil: Implications for indirect influence of temperature and moisture. Catena, 114, 90–96.
Huang, Z., Yu, Z. & Wang, M. (2014) Environmental controls and the influence of tree species on temporal variation in soil respiration in subtropical China. Plant and Soil, 1–13
IPCC (2013) Summary for policymakers. In Stocker, T.F., D. Qin, G.-K. Plattner, M. Tignor, S.K. Allen, J. Boschung, A. Nauels, Y. Xia, V. Bex and P.M. Midgley (Eds.), Climate Change 2013: The Physical Science Basis. Contribution of Working Group I to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change. Cambridge, United Kingdom and New York, NY, USA, Cambridge University Press.
Itoh, M., Kosugi, Y., Takanashi, S., Kanemitsu, S., Osaka, K. i., Hayashi, Y., Tani, M., & Rahim Nik, A. (2012). Effects of soil water status on the spatial variation of carbon dioxide, methane and nitrous oxide fluxes in tropical rain-forest soils in Peninsular Malaysia. Journal of Tropical Ecology, 28, 557–570.
Jauhiainen, J., Takahashi, H., Heikkinen, J. E., Martikainen, P. J., & Vasander, H. (2005). Carbon fluxes from a tropical peat swamp forest floor. Global Change Biology, 11, 1788–1797.
Jia, H. T., Zhu, X. P., Sheng, Y., Zhao, C. Y., Xu, Y. J., & Kadipov, K. G. (2013). Characteristics of soil CO2 emission of 3 kinds of woodland ecosystems in arid areas. Science of Soil and Water Conservation, 11, 95–98.
Jiang, H., Deng, Q., Zhou, G., Hui, D., Zhang, D., Liu, S., Chu, G., & Li, J. (2013). Responses of soil respiration and its temperature/moisture sensitivity to precipitation in three subtropical forests in southern China. Biogeosciences, 10, 3963–3982.
Katayama, A., Kume, T., Komatsu, H., Ohashi, M., Nakagawa, M., Yamashita, M., Otsuki, K., Suzuki, M., & Kumagai, T. (2009). Effect of forest structure on the spatial variation in soil respiration in a Bornean tropical rainforest. Agricultural and Forest Meteorology, 149, 1666–1673.
Kiese, R., & Butterbach-Bahl, K. (2002). N2O and CO2 emissions from three different tropical forest sites in the wet tropics of Queensland, Australia. Soil Biology and Biochemistry, 34, 975–987.
Koehler, B., Corre, M. D., Veldkamp, E., & Sueta, J. P. (2009). Chronic nitrogen addition causes a reduction in soil carbon dioxide efflux during the high stem-growth period in a tropical montane forest but no response from a tropical lowland forest on a decadal time scale. Biogeosciences, 6, 2973–2983.
Kosugi, Y., Mitani, T., Ltoh, M., Noguchi, S., Tani, M., Matsuo, N., Takanashi, S., Ohkubo, S., & Nik, A. R. (2007). Spatial and temporal variation in soil respiration in a Southeast Asian tropical rainforest. Agricultural and Forest Meteorology, 147, 35–47.
Li, H. F. (2011). Study on soil CO2 and CH4 fluxes in four typical plantations in Southern China. Journal of Zhejiang for Science & Technology, 31, 6–12.
Li, Y. D., Chen, B. F., & Zhou, G. Y. (2002). The research of tropical forests and biodiversity protection in Hainan, China. Beijing: China Forestry Publishing House.
Liang, J.B., (2002). Greenhouse gases and animal agriculture in Asia. Greenhouse Gases and Animal Agriculture: Proceedings, 15–20.
Lin, L. S., Han, S. J., Wang, Y. S., & Gu, Z. J. (2004). The study of soil CO2 efflux from four forest types in Changbaishan, China. Chinese Journal of Ecology, 23, 42–45.
Liptzin, D., Silver, W. L., & Detto, M. (2011). Temporal dynamics in soil oxygen and greenhouse gases in two humid tropical forests. Ecosystems, 14, 171–182.
Lou, Y., Li, Z., Zhang, T., & Liang, Y. (2004). CO2 emissions from subtropical arable soils of China. Soil Biology and Biochemistry, 36, 1835–1842.
Luo, T., Chen, B., Li, Y., Lin, M., Zhou, G., Chen, D., & Qiu, J. (2001). Litter and soil respiration in a tropical mountain rain forest in Jianfengling, Hainan Island. Acta Ecologica Sinica, 21, 2013–2017.
Matvienko, A. I., Makarov, M. I., & Menyailo, O. V. (2014). Biological sources of soil CO2 under Larix sibirica and Pinus sylvestris. Russian Journal of Ecology, 45, 174–180.
Mo, J. M., Fang, Y. T., Xu, G. L., Li, D. J., & Xue, J. H. (2005). The short-term responses of soil CO2 emission and CH4 uptake to simulated N deposition in nursery and forests of Dinghushan in subtropical China. Acta Ecologica Sinica, 25, 682–690.
Mosier, A. R. (1998). Soil processes and global change. Biology and Fertility of Soils, 27, 221–229.
Nottingham, A. T., Ccahuana, A. J. Q., & Meir, P. (2012). Soil properties in tropical montane cloud forests influence estimates of soil CO2 efflux. Agricultural and Forest Meteorology, 166, 215–220.
Ohkubo, S., Kosugi, Y., Takanashi, S., Mitani, T., & Tani, M. (2007). Comparison of the eddy covariance and automated closed chamber methods for evaluating nocturnal CO2 exchange in a Japanese cypress forest. Agricultural and Forest Meteorology, 142, 50–65.
Osuri, A. M., Kumar, V. S., & Sankaran, M. (2014). Altered stand structure and tree allometry reduce carbon storage in evergreen forest fragments in India’s Western Ghats. Forest Ecology and Management, 329, 375–383.
Pypker, T. G., & Fredeen, A. L. (2003). Below ground CO2 efflux from cut blocks of varying ages in sub-boreal British Columbia. Forest Ecology and Management, 172, 249–259.
Rowlings, D., Grace, P., Kiese, R., & Weier, K. (2012). Environmental factors controlling temporal and spatial variability in the soil-atmosphere exchange of CO2, CH4 and N2O from an Australian subtropical rainforest. Global Change Biology, 18, 726–738.
Schindlbacher, A., Zechmeister-Boltenstern, S., Glatzel, G., & Jandl, R. (2007). Winter soil respiration from an Austrian mountain forest. Agricultural and Forest Meteorology, 146, 205–215.
Schlesinger, W. H., & Andrews, J. A. (2000). Soil respiration and the global carbon cycle. Biogeochemistry, 48, 7–20.
Sha, L. Q., Zheng, Z., Tang, J. W., Wang, Y. H., Zhang, Y. P., Cao, M., Wang, R., Liu, G. G., Wang, Y. S., & Sun, Y. (2005). Soil respiration in tropical seasonal rain forest in Xishuangbanna, SW China. Science in China Series D: Earth Sciences, 48, 189–197.
Silver, W. L., Lugo, A., & Keller, M. (1999). Soil oxygen availability and biogeochemistry along rainfall and topographic gradients in upland wet tropical forest soils. Biogeochemistry, 44, 301–328.
Silvola, J., Alm, J., Ahlholm, U., Nykanen, H., & Martikainen, P. J. (1996). The contribution of plant roots to CO2 fluxes from organic soils. Biology and Fertility of Soils, 23, 126–131.
Singh, B. K., Bardgett, R. D., Smith, P., & Reay, D. S. (2010). Microorganisms and climate change: terrestrial feedbacks and mitigation options. Nature Reviews Microbiology, 8, 779–790.
Soe, A. R. B., & Buchmann, N. (2005). Spatial and temporal variations in soil respiration in relation to stand structure and soil parameters in an unmanaged beech forest. Tree Physiology, 25, 1427–1436.
Subke, J. A., Reichstein, M., & Tenhunen, J. D. (2003). Explaining temporal variation in soil CO2 efflux in a mature spruce forest in Southern Germany. Soil Biology and Biochemistry, 35, 1467–1483.
Tang, X. L., Liu, S. G., Zhou, G. Y., Zhang, D. Q., & Zhou, C. Y. (2006a). Soil-atmospheric exchange of CO2, CH4, and N2O in three subtropical forest ecosystems in southern China. Global Change Biology, 12, 546–560.
Tang, X. L., Zhou, G. Y., Liu, S. G., Zhang, D. Q., Liu, S. Z., Li, J., & Zhou, C. Y. (2006b). Dependence of soil respiration on soil temperature and soil moisture in successional forests in southern China. Journal of Integrative Plant Biology, 48, 654–663.
Thomas, M. V., Malhi, Y., Fenn, K. M., Fisher, J. B., Morecroft, M. D., Lloyd, C. R., Taylor, M. E., & McNeil, D. D. (2011). Carbon dioxide fluxes over an ancient broadleaved deciduous woodland in southern England. Biogeosciences, 8, 1595–1613.
van Groenigen, K. J., Osenberg, C. W., & Hungate, B. A. (2011). Increased soil emissions of potent greenhouse gases under increased atmospheric CO2. Nature, 475, 214–216.
von Arnold, K., Nilsson, M., Hanell, B., Weslien, P., & Klemedtsson, L. (2005). Fluxes of CO2, CH4 and N2O from drained organic soils in deciduous forests. Soil Biology and Biochemistry, 37, 1059–1071.
Wang, M., Han, S., & Wang, Y. (2004). Important factors controlling CO2 emission rates from forest soil. Chinese Journal of Ecology, 23, 242229.
Wangluk, S., Boonyawat, S., Diloksumpun, S., & Tongdeenok, P. (2013). Role of soil temperature and moisture on soil respiration in a teak plantation and mixed deciduous forest in Thailand. Journal of Tropical Forest Science, 25, 339–349.
Werner, C., Zheng, X. H., Tang, J. W., Xie, B. H., Liu, C. Y., Kiese, R., & Butterbach-Bahl, K. (2006). N2O, CH4 and CO2 emissions from seasonal tropical rainforests and a rubber plantation in Southwest China. Plant and Soil, 289, 335–353.
Wood, T. E., Cavaleri, M. A., & Reed, S. C. (2012). Tropical forest carbon balance in a warmer world: a critical review spanning microbial-to ecosystem-scale processes. Biological Reviews, 87, 912–927.
Wood, T. E., Detto, M., & Silver, W. L. (2013). Sensitivity of soil respiration to variability in soil moisture and temperature in a humid tropical forest. Plos One, 8, 7.
Wu, Z. M., Zeng, Q. B., Li, Y. D., Zhou, G. Y., Chen, B. F., Du, Z. H., & Lin, M. X. (1997). A preliminary research on the carbon storage and CO2 release of the tropical forest soils in Jianfengling, Hainan an Island, China. Acta Phytoecologica Sinica, 21, 8.
Xu, X. K., Han, L., Luo, X. B., Liu, Z. R., & Han, S. J. (2009). Effects of nitrogen addition on dissolved N2O and CO2, dissolved organic matter, and inorganic nitrogen in soil solution under a temperate old-growth forest. Geoderma, 151, 370–377.
Yan, J. H., Zhang, W., Wang, K. Y., Qin, F., Wang, W. T., Dai, H. T., & Li, P. X. (2014). Responses of CO2, N2O and CH4 fluxes between atmosphere and forest soil to changes in multiple environmental conditions. Global Change Biology, 20, 300–312.
Zanchi, F. B., Meesters, A., Waterloo, M. J., Kruijt, B., Kesselmeier, J., Luizao, F. J., & Dolman, A. J. (2014). Soil CO2 exchange in seven pristine Amazonian rain forest sites in relation to soil temperature. Agricultural and Forest Meteorology, 192, 96–107.
Zhang, J. H., Han, S. J., & Yu, G. R. (2006). Seasonal variation in carbon dioxide exchange over a 200-year-old Chinese broad-leaved Korean pine mixed forest. Agricultural and Forest Meteorology, 137, 150–165.
Zhou, W., Sha, L., Shen, S., & Zheng, Z. (2008). Seasonal change of soil respiration and its influence factors in rubber (Hevea brasiliensis) plantation in Xishuangbanna, SW China. Journal of Mountain Science, 26, 317–325.
Zhou, Z., Jiang, L., Du, E. Z., Hu, H. F., Li, Y. D., Chen, D. X., & Fang, J. Y. (2013). Temperature and substrate availability regulate soil respiration in the tropical mountain rainforests, Hainan Island, China. Journal of Plant Ecology, 6, 325–334.
Zhou, W. P., Hui, D. F., & Shen, W. J. (2014). Effects of soil moisture on the temperature sensitivity of soil heterotrophic respiration: a laboratory incubation study. Plos One, 9, 10.
Acknowledgments
This study was supported by 100 Talents Program of The Chinese Academy of Sciences, Program for New Century Excellent Talents in University (NCET-12-0477), the National Natural Science Foundation of China (nos. 31100348 and 41201205), International S&T Cooperation Program of China (S2013GI0408), and China Qianren Project. The authors give special thanks to Ms. Wan Xiong for her editing and valuable comments on the manuscript.
Author information
Authors and Affiliations
Corresponding authors
Rights and permissions
About this article

Cite this article
Jiang, X., Chen, H., Peng, C. et al. Soil Carbon Dioxide Fluxes from Three Forest Types of the Tropical Montane Rainforest on Hainan Island, China. Water Air Soil Pollut 227, 213 (2016). https://doi.org/10.1007/s11270-016-2904-1
Received:
Accepted:
Published:
DOI: https://doi.org/10.1007/s11270-016-2904-1