
Abstract
We have examined populations of brown trout in low-conductivity mountain lakes (5.0–13.7 μS/cm and 0.14–0.41 mg/l Ca) in southwestern Norway during the period 2000–2010. Inlets to the lakes were occasionally even more dilute (2007; conductivity = 2.9–4.8 μS/cm and Ca = 0.06–0.17 mg/l). The combination of pH and conductivity was the best predictor to fish status (CPUE), indicating that availability of essential ions was the primary restricting factor to fish populations in these extremely diluted water qualities. We suggest that conductivity <5 μS/cm is detrimental to early life stages of brown trout, and subsequently that there are lakes in these mountains having too low conductivity to support self-reproducing trout populations. Limited significance of alkalinity, Ca, Al, and color suggests that effects of ion deficit apparently overruled the effects of other parameters.
Similar content being viewed by others

Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
1 Introduction
Several studies have established the impact conductivity (ionic strength) has on brown trout populations (Bua and Snekvik 1972; Leivestad et al. 1976; Wright and Snekvik 1978). A large study of fish status (based on interviews) and water chemistry (pH and conductivity) based on 1,034 lakes in southern Norway in 1974–1979 showed the significance of both conductivity and pH in explaining the status of trout populations in these mountain regions (Sevaldrud and Muniz 1980). At conductivities <10 μS/cm, pH values >5.8 were required to sustain healthy populations of trout, but at conductivities >30 μS/cm, a pH value of 5.0 was sufficient. After 1980, the interaction between low conductivity and fish status has received very little attention. Brown trout status is currently explained using acid-neutralizing capacity (ANC), calcium (Ca), and labile aluminum (LAl) as the main water quality variables (Bulger et al. 1993; Hesthagen et al. 2008; Lien et al. 1996).
Due to osmotic effects, fish living in freshwater experience loss of ions, primarily across the gills. Fish compensate this loss with actively uptake of ions from the water. Ion transport in the opposite direction of osmotic gradients is an energy-intensive process, where several enzymes are participating (Heath 1995). In low ionic strength water, the uptake–loss balance of ions is particularly delicate, and easily disturbed by chemical agents as H+ and LAl (Brown 1983; Gensemer and Playle 1999; Hutchinson et al. 1989). Ca, however, stabilizes the gill membrane and prevents loss of ions (McWilliams and Potts 1978).
A persistent lack of correlation between fish status in these mountain areas and “modern” chemical parameters has actualized a reconsideration of the impact of conductivity. Despite the fact that acidification has decreased the last decades, and the pH values have increased considerably, stocking of brown trout have failed in many mountain lakes in southwestern Norway. Even restocking in lakes that earlier supported self-reproducing populations has yielded highly variable results (Dalziel et al. 1995). The chemical parameters used in the referred study (ANC, Ca, and LAl) did not explain fish status and stocking success.
Mountain lakes in southwestern Norway are characterized by very low conductivity, due to elevation, distance to the sea, and geology. A study performed in Rogaland in 2002 showed that most of the lakes above 500 m had conductivity values <20 μS/cm, and the lowest reported value was 4 μS/cm, corrected for the H+ contribution (Enge and Lura 2003). Values down to 3 μS/cm were measured in mountain lakes in Aust-Agder (neighbor county to Rogaland) in 1975 (Sevaldrud and Muniz 1980). Our hypothesis is that trout populations in these mountain lakes are restricted by availability of substantial ions. This hypothesis may also explain why a number of mountain lakes in Rogaland never have supported trout populations (Hesthagen et al. 1997; Sevaldrud and Muniz 1980), despite the fact that the physical conditions in many of these lakes apparently seem to be satisfactory.
For a more precise and updated elucidation of conductivity effects on fish status in extremely diluted water qualities, this study has compiled data from local surveys conducted in mountain lakes in southwestern Norway during the years 2000–2010. The study also includes analysis of two time series of water chemical monitoring during the period 1985–2010.
2 Study Area
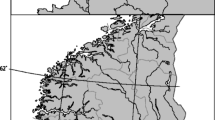
The lakes were situated at 427–1,021 m above sea level and located within the counties of Rogaland, Vest-, and Aust-Agder (Fig. 1 and Table 1). The bedrock is mainly of granitic origin, and most of the draining areas have barren rock and are located above the forest limited.

South Norway. Test-fished lakes are shown by black triangles
Meteorological data were available from the station “Tjørhom”, located in the middle of the project area at altitude 500 m. Annual average precipitation and temperature are 1,760 mm and 3.2°C, respectively (Eklima.no 2010). Average annual snow accumulation at Tjørhom in April is 290 mm water equivalents (Sira-Kvina Power Company, unpublished data). In the northern part of the project area (Auråhorten), average snow accumulation is 1,420 mm.
All lakes in this study supported self-reproducing brown trout populations originally, but the populations declined to a minimum in the 1960s and 1970s due to acidification (Sevaldrud and Muniz 1980). In the last decades, however, the trout populations in many of these mountain lakes have recovered considerably due to reduced acidification (Enge 2009). The current study does not include lakes in areas where trout went extinct, due to lack of population response to enhanced water quality.
Regulated lakes were also included in the study, but the regulations do not exclude natural reproduction (will be discussed later). Some of the regulated lakes are considered as “satellites” in large reservoirs. These lakes were only included in the reservoirs in years with high water level.
With one exception (Lake N. Førevatn), the brown trout populations were not influenced by stockings. In Lake N. Førevatn, however, all the trout caught were traced back to stockings, and were excluded from the statistical analyzes. In some of the lakes, brook trout (Salvelinus fontinalis) were stocked, but in Norway, this species has shown up to be inferior to healthy brown trout populations (Qvenild 1986).
Except for a limited net fishing of brook trout in some of the regulation reservoirs, the exploitation of the fish resources in the current lakes was generally poor.
Limed lakes were not included in this study.
3 Methods and Materials
Test fishing was conducted with Jensen series (Jensen 1972). The “classical” Jensen series includes eight nets of mesh size 52, 45, 39, 35, 29, 26, and 2 × 21 mm. The nets were normally of 25 m length and 1.5 m depth. For catching smaller fish, Jensen series were often extended with one or two small meshed nets (Jensen 1972), often denoted “extended” Jensen. In this study, Jensen series were extended with two nets, 16 and 13 mm, of 2 m depth.
The lakes were test-fished with 1–4 net series, usually in July (Table 2). Catch-per-unit-effort (CPUE) was calculated as fish per 100 m2 net per night of fishing.
Water samples were collected on the day of test fishing, with a standard Ruttner sampler, and transferred to acid-washed PE bottles. Samples were collected from depths of 0, 5, 10, 20, 30, and 40 m, depending on the maximum lake depth. Median values for all depths were used in the statistical tests.
To illustrate water chemical changes over time, analysis of two time series of water chemical monitoring was also included in the current study. These locations, “Gjuvatn” and “Degevatn”, drain large (100 and 70 km2) high altitude (800–1,500 m) mountain areas.
pH, conductivity, color, calcium, alkalinity, and aluminum were analyzed on all samples. Chloride was analyzed on about one third of the samples. pH, conductivity, and color were measured on the day of sampling, using a mobile field lab. As all major ions were not measured, ANC cannot be calculated. All conductivity values used or presented in this study were corrected for the H+ contribution (1 μEq/l, H+ = 0.35 μS/cm). The conductivity then represents the sum of dissolved salts in the water.
pH was determined with an Orion 221 digital pH meter with Radiometer pHC2401 electrode, calibrated with standard buffers (pH = 4.01 and 7.00). Conductivity was determined by conductivity meter HACH CO150, calibrated with standard KCl solutions. Color was measured visually with HACH comparator CO-1. Average of three independent color measurements on each sample was used. Calcium was determined potentiometrically with electrode Radiometer ISE25Ca and calomel reference. Alkalinity was titrated to fixed end point pH = 4.5 with sulfuric acid and calculated to equivalence alkalinity (ALKe) according to Henriksen (1982). Total monomeric Al was determined photometrically with ECR (Eaton et al. 1995). This method is an alternative to the more commonly used pyrocatechol method (Eaton et al. 1995). Chloride was determined by conductometric titration with silver nitrate.
The statistical analyzes were performed as backward elimination (BWE) and forward selection (FWS) multiple regressions, succeeded by standard t and F tests.
4 Results
In the 25 lakes, pH values ranged from 4.84 to 6.02, conductivity 5.0–13.7 μS/cm, and calcium 0.14–0.41 mg/l (Table 3). In 2007, the inlet rivers and lakes were even more dilute. pH values were in the range 5.39–5.73, conductivity 2.9–4.5 μS/cm, and Ca 0.06–0.17 mg/l (Appendix).
There were considerable internal correlations between several of the chemical parameters used in the test. Most of the parameters correlated significantly (p < 0.01) to 2–4 other parameters (Table 4)
Altitude correlated to all chemical parameters (r 2 = 0.29–0.62, p < 0.01, n = 25) but Ca.
The lake samples including chloride data showed that conductivity mainly was caused by sea salts, even in these high-altitude mountain lakes. Correlation between conductivity and chloride (Fig. 2) gave r 2 = 0.93 (p < 0.001, n = 67), and conductivity and calcium r 2 = 0.42 (p < 0.001, n = 67).

Lake chemistry: Conductivity, adjusted for the H+ contribution, versus chloride and calcium
The lakes test-fished in 2007 were excluded from the statistical tests because the water chemistry was deemed not representative (see Discussion). Trout was caught in 21 of the 25 remaining lakes. The four lakes without catch had neighbor lakes or tributaries with trout populations and possibilities of immigration. They are therefore included in the statistical tests. Lake L. Bossvatn was test-fished with “classical” Jensen. However, this lake had no fish. Electrofishing supports these findings. In this case, CPUE = 0 regardless of net series.
Alkalinity was the only single parameter contributing significantly to CPUE (r 2 = 0.28, p < 0.01, n = 25). Alkalinity, however, was highly correlated to other important parameters to fish (Table 4). Alkalinity yielded limited significance in combinations with pH, probably due to good correlation between these two parameters (Table 4). On somewhat lower level of significance, pH, color, and Ca contributed too (p < 0.05, n = 25).
In the multiple statistic analysis, backward elimination regression (BWE) suggested pH and conductivity as the most appropriate predictors to CPUE (Table 5). These parameters alone appear to be satisfactory in explaining CPUE (Fig. 3). Forward selection regression (FWS) on the other hand suggested alkalinity and color (Table 5).

Fish status and water chemistry (pH and conductivity, corrected for the H+ contribution), stratified by three ranges of CPUE
The FWS regression yielded slightly lower correlation (r 2) and somewhat higher residual standard deviation than the BWE regression.
Residual analysis suggested 2–3 outliers. r 2 values of about 0.7–0.8 were obtained by excluding these apparent outliers. Because of the limited data material, we have, however, decided to not exclude any observations.
There were no time dependence in the residuals (p > 0.01), suggesting limited biological delay effects due to possible changes in water chemistry during the period 2000–2010.
pH values in “Gjuvatn” and “Degevatn” showed a distinct increase during the period 1985–2009 (Degevatn: r 2 = 0.65, p < 0.001, n = 98; Gjuvatn: r 2 = 0.73, p < 0.001, n = 90). Conductivity on the other hand has decreased considerably (Degevatn: r 2 = 0.34, p < 0.001, n = 72; Gjuvatn: r 2 = 0.71, p < 0.001, n = 67) the last two decades (Table 6).
5 Discussion
Trout normally reproduce in inlet rivers and brooks. Ideally, the water chemistry should have been sampled from the inlets. However, there are severe practical problems in sampling a large number of brooks over some time in these remote areas. A large number of samples from each brook were also required to get a representative selection, due to variations in water chemistry.
The extremely low calcium and conductivity values in the water reflect extremely low rates of chemical weathering in the catchment soils. Lake chemistry then represents an integrated sample of inlet chemistries during a period of time, depending on retention time of the lake.
In these mountain areas, the trout eggs hatch during early summer (June), so lake chemistry in the summer (July/August) represents an “average” inlet water quality experienced by critical stages for fry and eggs. Because of stratification, there are some differences in chemistry throughout the water column. We have used median chemistry of samples from several depths in this study.
CPUE provides a measure of population density in lakes. However, there is no strict linearity between CPUE and trout populations, because low population density increases the activity of the fish and thus the CPUE (Borgstrøm 1992). In these mountains, CPUE <2.5 n/100 m2 is often associated with sparse populations, while >7.5 n/100 m2 is normally registered in overstocked populations (Fig. 3). These limits were probably somewhat lower than generally used in Norway (Ugedal et al. 2005), but were justified due to the fact that the current lakes were ultraoligothropic mountain lakes.
The BWE multiple regressions suggested pH and conductivity as the best predictors to CPUE, but FWS suggested alkalinity and color. Both BWE and FWS regressions, however, suffer from major drawbacks (Sokal and Rohlf 1994). Once a variable has been added to FWS, it remains in the predictor set despite of eventually loss of significance when adding other variables. In BWE regressions, an eliminated variable stays out of the predictor set even if it may become an excellent predictor at a later stage. Such effects were primarily caused by intercorrelated predictor variables (Sokal and Rohlf 1994). Bulger et al. (1993) suggested that insufficient interpretation of these phenomena in earlier studies has lead to misleading conclusions regarding chemical parameters and effects on fish (calcium). Similar problems occur when variables have limited significance alone, but appear significant only in combinations with certain other variables (Bulger et al. 1993). The last type of variables will never enter a standard FWS regression. Thus, results of multiple regressions should always be validated with biological theory (Sokal and Rohlf 1994).
FWS regression suggested two parameters (alkalinity and color), both of limited direct physiological significance (Gensemer and Playle 1999; Heath 1995). The apparent effects of these parameters were probably caused by correlation to a number of other important parameters (Table 4), and/or to a modifying effect on parameters of direct significance (e.g., color and LAl). The parameters suggested by the BWE regression however either have direct physiological significance (H+) or represent essential ions (conductivity).
However, left-out variables were not necessarily unimportant. They may simply be correlated with other variables already included in the model (Sokal and Rohlf 1994). In the current data material, alkalinity, as single parameter, contributed slightly more to CPUE than pH. When alkalinity has been added to FWS, this excluded subsequent addition of pH, due to the good correlation between these two parameters (r 2 = 0.83, p < 0.001, n = 25). In combinations simultaneously including pH, alkalinity, and other parameters, pH always contributed more significantly than alkalinity (Table 5).
In a “forced” FWS regression (pH as default), alkalinity never entered the model. Already, the first step suggested pH and conductivity as the best predictor set (same as BWE).
The FWS regressions also yielded somewhat lower correlation (r 2), and slightly higher residual standard deviation.
These findings, both of statistical and physiological nature, suggest that BWE was the most relevant regression method, and that the FWS regression, used on the current data material, yielded questionable results, due to various effects of intercorrelated predictor variables.
In a study of density of brown trout in rivers and brooks in southwestern Norway, Hesthagen et al. (1999) found limited effect of 8–44 μg/l LAl (mean values), and concluded that LAl in this range did not have any toxic effect on brown trout. In the current study, total monomeric Al were 51 ± 19 μg/l (n = 25). Due to the fact that this Al fraction includes not only LAl but also nontoxic fractions of Al, we may conclude that the present levels of Al probably were too low to have any significant effects on fish populations. This also establishes that the apparent color effects, suggested by the FWS regression, indeed were caused by correlation to other parameters, due to the fact that Al levels probably were too low to cause any harm, independent of speciation.
Several studies have established the positive effect of calcium on fish (Brown and Lynam 1981; Brown 1981a, b; 1983; Bulger et al. 1993; Hesthagen et al. 1999). Calcium affects the gill permeability and prevents loss of ions (McWilliams and Potts 1978). At calcium 2.0 mg/l, Brown (1983) found nearly complete survival of brown trout fry, even at Al (total monomeric Al) as high as 250 μg/l. Hesthagen et al. (1999) found that calcium had a positive effect on density of young brown trout in brooks even at quite low levels (mean values 0.35–0.84 mg Ca per liter). Wathne and Rosseland (2000) found that a calcium level at 0.38 mg/l was required to sustain life of brown trout in mountain lakes. However, there were apparently no effects of calcium on CPUE in the present study (Table 5). The statistical tests indicated that conductivity (ionic strength) was a better predictor to fish status than calcium in these extremely diluted water qualities, and that Ca levels probably were below a threshold where decreasing Ca had limited effect.
The apparent positive effect of conductivity is caused by effects of essential ions. Brown (1981a) suggested that an apparent conductivity effect in fact is a calcium effect. Other studies have reported positive effects of sodium (Hesthagen et al. 1999; Hutchinson et al. 1989), but Brown and Lynam (1981) did only find positive effect of sodium in the presence of calcium. Because of the marine origin of conductivity in lakes in this study (Fig. 2), sodium effects cannot be rejected. Wright and Snekvik (1978), however, did not find any effects of either sodium or chloride on population status in 700 lakes in southern Norway. Hesthagen et al. (1999) analyzed the effect of various ions and found positive effect of cation sum on fish densities in brooks. This study did not reject a possible positive effect of ionic strength itself in very soft water qualities. Hutchinson et al. (1989) suggested that measurements of conductivity or cation sum might be as useful as specific ion analysis. Bulger et al. (1993) found that pH and seven major ions correlated to fish status equally well as pH and LAl. This suggests that certain combinations of ions are essential, and that these combinations, in dilute water qualities, are adequately described by conductivity. The good correlations between “lumped parameters” (various ion sums and conductivity) and fish status also suggest possible effects of ionic strength itself.
In the regulated lakes, the breeding habitat located in tributaries crossing the regulation zone was considered as lost. This damage increases with increasing regulation height. Most of the regulated lakes, however, receive water from a number of rivers and brooks, and only a fraction of the original suitable breeding area in these tributaries was located in the regulation zone. There were found no effects of regulation height on CPUE (p > 0.01), suggesting that the breeding areas above the regulation zone were sufficient in the current lakes.
Due to large snow accumulation in these mountains in 2007, the water quality of the lakes became extremely diluted throughout the summer. In the lakes of Storsteindalen, the conductivity and calcium values were 2.9–3.8 μS/cm and 0.06–0.16 mg/l, respectively (Appendix). Kringlevatn had a dense population of trout at pH = 5.69 and conductivity = 4.8 μS/cm in 2007 (Appendix). At this conductivity, pH >6 is required to sustain life of brown trout (extrapolated from Fig. 3). If the 2007 water quality represents a permanent change, the trout population in Kringlevatn may disappear over some time. Thus, data from 2007 were considered as not representative, and excluded from the statistical tests.
No trout fry were registered during electrofishing of brooks in the Kringlevatn area in 2007, suggesting that extremely diluted water qualities are lethal to early life stages. Dietrich et al. (1989) reported acute toxicity of lake water in Lake Laiozza to 1- and 2-year-old brown trout at pH = 5.37 ± 0.22, conductivity = 7 μS/cm, LAl = 41–52 μg/l, and Ca = 0.5 mg/l. The 2007 water quality in the Kringlevatn area was considerably more dilute than Lake Laiozza.
The ionic balance of fish is a result of uptake and loss of ions. Many of the referred studies have focused on loss of ions. It is well known that both H+ and Al cause leakage of ions over the gills, and that Ca prevents this loss (Gensemer and Playle 1999). Conductivity, and not “leakage parameters”, was correlated to fish status, suggesting that availability of essential ions (such as Na and Cl) might be critical in these extremely diluted water qualities.
Of the lakes in Rogaland County, 13.4% have never supported trout populations (Hesthagen et al. 1997), including the four most dilute waters in the County Governors study (Enge and Lura 2003). These four lakes had conductivity 4.0–5.6 μS/cm and Ca 0.13–0.19 mg/l. pH requirements at these low conductivities are estimated to approximately 6.5 (extrapolated from Fig. 3). Based on general water chemical considerations, such pH values are obviously not consistent with Ca levels <0.2 mg/l, leading to the conclusion that water chemistry excludes brown trout from these lakes.
Restocking has failed in many lakes, despite the fact that pH has increased considerably the last decades (Table 6). The coincidental decrease in conductivity has however caused a considerable increase in pH requirements (Fig. 3), and to some degree “neutralized” the positive effect of increased pH.
The current study concludes that low conductivity must be addressed as a possible cause for reduced trout population health in dilute water qualities, due to lack of either essential ions, or combinations of ions. The relationship between CPUE and the variables pH and conductivity appears to be satisfactory in explaining most of the variability in fish density. These findings are probably not unique to southern Norway, and should be evaluated as a fish-restricting element in other high mountain areas too.
References
Borgstrøm, R. (1992). Effect of population density on gillnet catch ability in four allopatric populations of brown trout (Salmo trutta). Canadian Journal of Fisheries and Aquatic Sciences, 49, 1539–1545.
Brown, D. J. A. (1981a). The effect of pH and calcium on fish and fisheries. Water, Air, and Soil Pollution, 18, 343–351.
Brown, D. J. A. (1981b). The effects of various cations on the survival of brown trout, Salmo trutta at low pHs. Journal of Fish Biology, 18, 31–40.
Brown, D. J. A. (1983). Effect of calcium and aluminium concentrations on the survival of brown trout (Salmo trutta) at low pH. Bulletin of Environmental Contamination and Toxicology, 30, 582–587.
Brown, D.J.A., & Lynam, S. (1981) The effect of sodium and calcium concentrations on the hatching of eggs and the survival of the yolk sac fry of brown trout, Salmo trutta L. at low pH. The Fisheries Society of the British Isles, 205–211
Bua, B. & Snekvik, E. (1972). Klekkeforsøk med rogn av laksefisk 1966–1971. Virkninger av surhet og saltinnhold i klekkevannet. VANN 1–72: 86–93 (in Norwegian. Hatching experiments with roe of salmonids 1966–1971. Effects of pH and ionic strength of the hatchery water)
Bulger, A. J., Lien, L., Cosby, B. J., & Henriksen, A. (1993). Brown trout (Salmo trutta) status and chemistry from the Norwegian Thousand Lake Survey: statistical analysis. Canadian Journal of Fisheries and Aquatic Sciences, 50, 575–585.
Dalziel, T. R. K., Kroglund, F., Lien, L., & Rosseland, B. O. (1995). The REFISH (Restoring Endangered Fish In Stressed Habitats) Project, 1988–1994. Water, Air, and Soil Pollution, 85(2), 321–326.
Dietrich, D., Schlatter, C. H., Blau, N., & Fisher, M. (1989). Aluminium and acid rain: mitigating effects of NaCl on aluminium toxicity to brown trout (Salmo trutta fario) in acid water. Toxicological and Environmental Chemistry, 19, 17–23.
Eaton, A. D., Clesceri, L. S., Greenberg, A. E. (Eds) (1995). Standard methods for the examination of water and wastewater (19.edt.). American public health association, American water works association & water environment federation, Washington DC
Eklima.no (Norwegian Meteorological Institutes public access database) (2010)
Enge, E. (2009). Sira-Kvina utbyggingen - Effekter på vannkjemi, forsuringssituasjon og fiskebestander i Sira. MSc-thesis, University of Stavanger (In Norwegian. Sira-Kvina—effects on water chemistry, acidification and fish populations in river Sira)
Enge, E. & Lura, H. (2003). Forsuringsstatus i Rogaland 2002. Ambio Environmental Consultants & County Governor of Rogaland. (in Norwegian. Acidification status in Rogaland 2002)
Gensemer, R. W., & Playle, R. C. (1999). The bioavailability and toxicity of aluminium in aquatic environments. Critical reviews in Environmental Science and Technology, 29(4), 315–450.
Heath, A. G. (1995). Water pollution and fish physiology (2nd ed.). Boca Raton: CRC. 2000.
Henriksen, A. (1982). Alkalinity and acid precipitation research. Vatten, 38, 83–85.
Hesthagen, T., Sevaldrud, I.H., Berger, H.M. (1997). Assessment of damage to fish populations in Norwegian lakes due to acidification. Manus in: Hesthagen, T. 1997: Population responses of Arctic char (Salvelinus alpinus L.) and brown trout (Salmo trutta L.) to acidification in Norwegian inland waters. PhD thesis, Norwegian University of Science and Technology
Hesthagen, T., Heggenes, J., Larsen, B. M., Berger, H. M., & Forseth, T. (1999). Effects of water chemistry and habitat on the density of young brown trout (Salmo trutta) in acidic streams. Water, Air, and Soil Pollution, 112, 85–106.
Hesthagen, T., Fiske, P., & Skjelkvåle, B. L. (2008). Critical limits for acid neutralizing capacity of brown trout (Salmo trutta) in Norwegian lakes differing in organic carbon concentrations. Aquatic Ecology, 42, 307–316. doi:10.1007/s10452-008-9191-x.
Hutchinson, N. J., Holtze, K. E., Munro, J. R., & Pawson, T. W. (1989). Modifying effects of life stage, ionic strength and post-exposure morality on lethality of H+ and Al to lake trout and brook trout. Aquatic Toxicology, 15, 1–26.
Jensen, K. W. (1972). Drift av fiskevann. Fisk og fiskestell, 5, 1–61. Direktoratet for vilt og ferskvannsfisk. (in Norwegian. Managing fishing lakes. Directorate for Wildlife and Freshwater Fish).
Leivestad, H., Hendrey, G, Muniz, I.P. Snekvik, E. (1976). Effects of acid precipitation on freshwater organisms. In: Brække, F.H. (Ed.) Impact of acid precipitation on forest and freshwater ecosystems in Norway (86–111). SNSF, Research Report 6/76
Lien, L., Raddum, G. G., Fjellheim, A., & Henriksen, A. (1996). A critical limit for acid neutralizing capacity in Norwegian surface waters, based on new analyses of fish and Invertebrate responses. The Science of the Total Environment, 177, 173–193.
McWilliams, P. G., & Potts, W. T. W. (1978). The effects of pH and calcium concentrations on gill potentials in the brown trout, Salmo trutta. Comp. Physiol., 126, 277–286.
Qvenild, T. (1986). Utsettinger av bekkerøye i Norge. Fisk og fiskestell, 9, 1–41. Direktoratet for vilt og ferskvannsfisk. (in Norwegian. Stockings of brook trout in Norway. Directorate for Wildlife and Freshwater Fish).
Sevaldrud, I. & Muniz, I.P. (1980). Sure vatn og innlandsfiske i Norge. Resultater fra intervjuundersøkelsene 1974–1979. SNSF-project, IR77/80 (in Norwegian, summary in English: Acid lakes and inland fisheries in Norway. Results from interviews from 1974–1979
Sokal, R. R., & Rohlf, F. J. (1994). Biometry (3rd ed., p. 887). New York: Freeman.
Ugedal, O., Forseth, T., Hesthagen, T. (2005). Garnfangster og størrelse på gytefisk som hjelpemiddel i karakterisering av aurebestander. NINA-report 75–2005. (in Norwegian: Characterizing trout populations by net catch and size of spawners)
Wathne, B. M., & Rosseland, B. O. (Eds) (2000). MOLAR final report 4/1999. Measuring and modelling the dynamic response of remote mountain lake ecosystems to environmental change: a program of Mountain Lake Research—MOLAR. NIVA, Report nr. 96061-1
Wright, R. F., & Snekvik, E. (1978). Acid precipitation: chemistry and fish population in 700 lakes in southernmost Norway. Verh. Int. Ver. Limnol., 20, 765–775.
Acknowledgments
The authors thank Dr Richard F. Wright for valuable comments to this paper, Associate Professor Bjørn Auestad for performing the regression analysis, and Tom Arild Homme for testfishing 3 of the lakes. The registrations of fish and water chemistry were financed by Sira-Kvina Power Company (n = 24) and the County Governor of Aust-Agder (n = 1).
Author information
Authors and Affiliations
Corresponding author
Appendix
Appendix
Rights and permissions
About this article
Cite this article
Enge, E., Kroglund, F. Population Density of Brown Trout (Salmo trutta) in Extremely Dilute Water Qualities in Mountain Lakes in Southwestern Norway. Water Air Soil Pollut 219, 489–499 (2011). https://doi.org/10.1007/s11270-010-0722-4
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11270-010-0722-4