
Abstract
The biodiversity value of human-modified landscapes has become a central question in the tropical forest conservation biology, yet the degree to which plant populations and communities are restructured in response to environmental change remains unclear. Here, we address tree species density in a fragmented Atlantic forest landscape to test the hypothesis that tree assemblages inhabiting edge-dominated forest habitats approach typical conditions of early successional systems. Seedlings and adults from 141 tree species were sampled across 39 0.1-ha plots: 19 in small fragments (<100 ha) and 20 in mature forest stands within the interior of the largest forest remnant of the study landscape (3500 ha). A total of 5448 seedlings and adults were recorded, with >55 % of all tree species exhibiting higher densities in small fragments than in mature forest, particularly pioneers (>60 % of all species). Seedlings and adults of these proliferating species differed from species exhibiting population declines in terms of wood density and seed size, respectively. Additionally, pioneers were more abundant than shade-tolerant species, as were hardwood species in the case of seedlings. Tree species showing highest population increases consisted largely of long-lived, light-demanding canopy species bearing soft or hardwood and small-to-medium-sized seeds. Tree assemblage structure also differed in terms of forest habitats with small forest fragments supporting few rare species, whereas the most rapidly proliferating species were much more widespread and abundant in fragments. However, 60 % of all adult pioneer species recorded in small fragments were not recorded as seedlings in this habitat type, although both seedling and adult assemblages were dominated by pioneer species. Edge-dominated tree assemblages are likely to experience long-term shifts toward greater dominance of long-lived, pioneer canopy species.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Tropical forests represent the world’s most diverse biome, accounting for the provision of key ecosystem services such as carbon storage and climate regulation (Avissar and Werth 2005; FAO 2010). However, mounting human population pressure driven by greater demand for forest commodities and agricultural land is expected to threaten the largest remaining tracts of old-growth forests globally (FAO 2011; Laurance et al. 2014). In particular, habitat loss and fragmentation are major threat processes to tropical biodiversity, driving species extinction at multiple scales (Sala et al. 2000). Accordingly, it has been argued that large tropical forest reserves represent an irreplaceable biodiversity conservation strategy and a core element of any effective conservation initiative aiming to safeguard representative portions of regional biodiversity (Laurance 2005; Peres 2005).
However, large forest remnants are either dwindling or missing in many regions, including biodiversity hotspots and centers of species endemism (Ribeiro et al. 2009). Therefore, human-modified landscapes have attracted increasing interest from conservation biologists, particularly as complementary landscape elements such as biodiversity corridors or biodiversity-friendly matrices (Melo et al. 2013). In this context, recent research efforts have identified (1) several ecological or functional groups sensitive to human disturbances, most of which referred to as forest-dependent species (Girão et al. 2007; Lopes et al. 2009; Melo et al. 2010; Santos et al. 2008; Tabarelli et al. 2010a) and (2) the presence of altered but rather impoverished communities in taxonomic, ecological, and phylogenetic terms along edge-affected habitats (Santo-Silva et al. 2013; Santos et al. 2008, 2010). Population declines, species extinctions, shifts in community structure, and collapse in forest structure proceed rapidly as soon as forest edges are created. Although species losses are expected to reach a relaxation point over time, the ultimate nature/composition of tree assemblages in human-modified landscapes remains an open question (see Arroyo-Rodriguez et al. 2009, 2012; Santos et al. 2008, 2010).
The Brazilian Atlantic forest biome has a long history of human disturbances and over 80 % of all forest remnants are smaller than 50 ha (Ribeiro et al. 2009). Tree assemblages inhabiting forest edges and small forest fragments (i.e., edge-affected habitats) have provided key evidence on the potential effects imposed by habitat loss and fragmentation, particularly the establishment of forest edges immersed into open-habitat matrices (see syntheses in Joly et al. 2014; Tabarelli et al. 2010b). Impoverished tree assemblages are dominated by some pioneer species, which tend to proliferate from local to regional scales (Lôbo et al. 2011; Tabarelli et al. 2010a). Proliferation of pioneer and light-demanding species (successional species sensu Laurance et al. 2006b) parallels population collapses and extirpations of species in several ecological groups belonging to the old-growth flora, such as large-seeded (Santos et al. 2008) and emergent species (Oliveira et al. 2008), and those with specialized floral biology (Girão et al. 2007; Lopes et al. 2009). Collectively, these shifts have been interpreted as a successional trajectory triggered by habitat fragmentation, transforming edge-affected habitats into early successional systems (Tabarelli et al. 2008). However, it remains unclear whether shifts in plant assemblage composition have already reached a saturation point even after centuries of forest conversion.
Here, we address the population density of 141 tree species as both seedlings and adults in a fragmented Atlantic forest landscape with a long history of sugarcane production. We tested the hypothesis that tree assemblages inhabiting edge-dominated forest habitats with no record of clear-cutting approach typical conditions of early successional systems; this change is more permanent rather than transitory (Tabarelli et al. 2008). Briefly, we document population density and the structure of tree assemblages in adult and seedling stages to project the magnitude of fragmentation-related effects and the future of these assemblages in small forest fragments. We identify which species seems to be favored and disadvantaged in small forest fragments based on their density ratio (density in small forest fragments over their density in mature areas of a large forest remnant), considering both age classes. Density ratio is examined across functional groups relative to regeneration strategy, seed size, and wood density. Finally, we discuss the implications of our findings in the light of current knowledge on tree assemblage responses to habitat fragmentation and the conservation value of human-modified landscapes.
Materials and methods
Study area
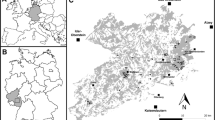
This study was carried out in a severely fragmented landscape of the Brazilian northeastern Atlantic forest, located within the 66,700-ha Usina Serra Grande landholding (8°58′50″S, 36°04′30″W; Fig. 1; Table S1). The area is comprised largely of sugarcane monoculture surrounding 109 forest fragments ranging in size between 1.6 and 3500 ha. The region presents a tropical climate, with a mean annual rainfall of ~2000 mm, and temperatures between 22 and 24 °C, with a dry season lasting 3 months (November–January). At a regional scale, the evergreen or semideciduous forest remnants (Veloso et al. 1991) are largely dominated by Lauraceae, Sapotaceae, Moraceae, and Fabaceae tree species (Grillo et al. 2005).

Map of the Serra Grande landscape and the distribution of forest fragments investigated
Serra Grande represents a long-standing fragmented forest landscape, given that sugarcane plantations date back to the nineteenth century (Santos et al. 2008). In addition, the size and shape of forest fragments have remained stable to ensure water supply for sugarcane irrigation. The Serra Grande landscape also holds the 3500-ha Coimbra Forest, the largest and best preserved forest patch in the region, which still retains large areas of mature forest with typical elements of undisturbed primary forest, such as large-seeded tree species and large frugivorous birds (Oliveira et al. 2004; Santos et al. 2008).
Seedling and adult tree surveys
Surveys of adult and seedling trees were carried out only in the dry seasons. Adult trees were surveyed from 2003 to 2005, while seedling inventory was carried out from 2007 to 2009. Tree assemblages were sampled in 39 plots of 0.1-ha (10 × 100 m) within two habitat types: (1) mature forest—20 plots in old-growth forest stands located inside the Coimbra Forest and at least 200 m from the nearest edge and (2) small forest fragments—19 plots in 19 forest fragments (one plot placed at the geometric center of each fragment) ranging in size from 3.4 to 91.2 ha (mean = 40.9 ha; median = 31.7 ha). This set of forest fragments was selected because (1) the remaining Atlantic forest is vastly dominated by patches smaller than 100 ha (Ribeiro et al. 2009); (2) they have not been exposed to clear-cutting; and (3) they are isolated from continuous forest for more than a century (Santos et al. 2008). As previously stated (Santos et al. 2008), we acknowledge that the Coimbra Forest consists of a single and unreplicated tract of forest and thereby it does not truly represent a ‘continuous forest.’ However, Coimbra is the best available contemporary scenario and its undisturbed areas of forest interior have been successfully adopted as ‘control’ sites to assess the long-term effects of habitat fragmentation on the structure of the plant assemblages (see for example Girão et al. 2007; Lopes et al. 2009; Santo-Silva et al. 2013; Santos et al. 2008; Tabarelli et al. 2010a).
For adult tree assemblages, we surveyed all stems ≥10 cm DBH (diameter at breast height). During seedling surveys, we randomly selected 100 seedlings (10–50 cm in total height) in 0.1-ha plots. In particular, we set 200 random points within each plot and sampled the nearest seedling to each point. We subsequently discarded all seedlings that did not belong to the tree species, and then randomly selected a subset of 100 seedlings. We decided to use this methodology to avoid sampling monospecific seedling aggregations and to ensure an equal sample size across all sites. These overlapping adult and seedling tree assemblages have yet to be examined together, mainly in functional terms, but previous studies have considered them separately [Santos et al. (2008) and Santo-Silva et al. (2013), respectively]. The spatial distribution of plots is necessarily limited because all mature forest areas are restricted to the same large fragment (Coimbra Forest). However, the overall distribution of the tree flora in the Serra Grande landscape is not affected by large-scale spatial effects (Santos et al. 2008).
Regeneration niche, seed size, and wood density
To identify likely ecological attributes driving changes in population density, we assigned each species into functional groups according to their regeneration niche, seed size, and wood density, which is a good correlate of individual growth rate (Enquist et al. 1999). In terms of regeneration niche, all stems were assigned into either (1) pioneer or (2) shade-tolerant species (Hartshorn 1978). For seed size, species were assigned into four groups: (1) small (<6 mm in length), (2) medium-sized (6–15 mm), (3) large (>15–30 mm), and (4) very large (>30 mm). For wood density, we assigned each species into two major groups: (1) softwood (wood density <0.7 g/cm3) or (2) hardwood (>0.7 g/cm3). These criteria for assigning species into functional groupings had been adopted previously (Michalski et al. 2007; Oliveira et al. 2004; Santo-Silva et al. 2013; Santos et al. 2008). Information on regeneration niche and seed size were obtained from a comprehensive literature review, including floras and checklists (e.g., Lorenzi 1992; Oliveira 2007; van Roosmalen 1985), and complemented by our own personal knowledge following over two decades of research on the Atlantic forest tree species (e.g., Melo et al. 2006; Oliveira et al. 2004; Santo-Silva et al. 2013; Silva and Tabarelli 2000; Tabarelli et al. 1999; Tabarelli and Peres 2002). Data of wood density were obtained from Zanne et al. (2009), and we used average values for genera or families if species-specific data were unavailable.
Data analysis
For each ontogenetic stage (seedlings and adults) and habitat type (small forest fragments and mature forest), we calculated the average population density of each species (i.e., total abundance divided by the number of 0.1-ha plots). To infer on either positive or negative changes in population densities in small fragments, we calculated separately for adults and seedlings the ratio between the density in small fragments and the density in mature forest for each species (hereafter, SF/MF ratio). In the lack of long-term demography data (which are logistically and financially unfeasible in most circumstances), this ratio is a very simple metric indicating potential cross-habitat changes in species abundance of even presence (i.e., SF/MF ratios >1 and <1 indicate species that were either more abundant or less abundant in small fragments than in mature forest areas, respectively). However, this metric must be interpreted with caution. Since some species were only found in either small fragments or mature forest, we arbitrarily added one individual for all species in each habitat type in order to enable estimates of the SF/MF ratio.
Using only those species that had been assigned into a functional group in terms of regeneration niche, seed size, and wood density, we first performed χ 2 tests to evaluate if species with SF/MF ratio >1 and SF/MF ratio <1 (hereafter proliferating and declining species, respectively) differed in terms of functional composition. Second, we performed GLMs (family = Gaussian; link = identity) adopting the regeneration niche, seed size, and wood density as factors and the ln-transformed SF/MF ratio as a continuous-response variable. χ 2 tests and Linear Models were performed separately for adults and seedlings. Finally, we provide species rank curves on the basis of both species abundance and distribution (number of plots) for each habitat (Magurran 1988), considering both adult and seedling assemblages. To obtain species rank curves, all species recorded as adult and seedling were considered, even those not assigned into functional groups. All analyses were performed in Statistica 7 and in R (R Core Team 2015).
Results
We recorded a total of 2778 adult trees representing 103 species (Table S2), 58 (56.3 %) of which were assigned to proliferating species. Consistent with the pattern recorded for seedlings, this group consisted predominantly of pioneers (62.1 %) and softwood species (70.7 %) bearing medium-sized (60.3 %) and small seeds (24.1 %). Considering the whole adult flora recorded at both habitats, there were no differences between proliferating and declining species in relation to regeneration niche (χ 2 = 0.19, df = 1, P = 0.659; Fig. 2a) and wood density (χ 2 = 1.29, df = 1, P = 0.256; Fig. 2c). However, large- and very large-seeded adult trees were more frequent among declining species than those that were proliferating (χ 2 = 8.23, df = 3, P = 0.041; Fig. 2b).

Percentage of species assigned into functional groups in terms of regeneration niche, seed size, and wood density at the Serra Grande-fragmented landscape. a–c indicate the percentages of species in adult assemblages, whereas d–f indicate the percentages of species in seedling assemblages. Open bars represent the percentage of species with SF/MF ratios <1, whereas black bars represent those with a SF/MF >1. Pio pioneer, ST shade-tolerant, n.s. nonsignificant, *P < 0.05
Considering the seedling flora, a total of 2640 individuals representing 76 tree species were recorded across both mature forest and small fragments (Table S2), with 48 species (63.2 %) exhibiting higher abundances in small fragments (SF/MF ratios >1). Most of these proliferating species had been classed as pioneers (72.9 %), hard-wooded (54.2 %), and bearing either medium-sized (50 %) or small seeds (27.1 %). Furthermore, proliferating and declining species in seedling assemblages did not differ in terms of regeneration niche (χ 2 = 1.21, df = 1, P = 0.27; Fig. 2d) and seed size (χ 2 = 5.76, df = 3, P = 0.124; Fig. 2e), but the percentage of softwood species was one-third lower in proliferating species (χ 2 = 6.12, df = 1, P = 0.013; Fig. 2f).
The SF/MF ratios were highly variable ranging from 0.004 to 81.1 in seedling assemblages and from 0.003 to 77.9 in adult assemblages. Among all functional groups, pioneers had higher SF/MF ratios than shade-tolerant species (Adults, F = 4.82, P = 0.031; Seedlings, F = 5.21, P = 0.025; Fig. 3a, b), whereas hardwood species had higher SF/MF ratios than softwood species in seedlings (F = 10.44, P = 0.002). Although pioneer species on average exhibited higher SF/MF ratios than shade-tolerant species in both ontogenetic stages, this elevated density was more pronounced in seedlings than in adults. When we compared population densities between adult and seedling assemblages, almost all shade-tolerant species exhibited a reduced density at the seedling stage in both habitat types (Fig. 4a), whereas pioneer species were much more evenly split between those exhibiting either lower or greater densities at the seedling stage (Fig. 4b). Considering both adult and seedling assemblages, hyperdominant species in small fragments exhibiting the highest SF/MF ratios were predominantly long-lived, canopy gap-demanding species bearing small-to-medium-sized seeds (Table 1).

Relationship between SF/MF ratios and the regeneration niche of a adults and b seedlings. Boxes indicate the 25th and 75th percentiles, error bars represent the 10th and 90th percentiles, and the points are the outliers. Solid and dashed lines indicate the median and mean, respectively. *P < 0.05

Relationship between population density in adult and seedling tree assemblages for a shade-tolerant and b pioneer species, considering only those species that were found in both ontogenetic stages. Each arrow represents one species and the end of the arrow (black dots) indicates density in adult assemblages, while the arrow heads indicate the density in seedling assemblages in mature forest areas and small forest fragments
Finally, adult and seedling assemblages differed markedly in terms of community structure across habitat types since only a small number of rare seedling species were recorded in small fragments (Fig. 5), whereby the number of species represented by only one or two individuals in small fragment plots was 81 % lower compared to those in mature forest areas. Among the ten top ranking proliferating species (highest SF/MF ratios), only three were recorded as both adults and seedlings in mature forest, where they were typically rare. In contrast, most of these top ten species were very abundant in small fragments as both seedling and adults, including the pioneers B. sericea, B. virgilioides, C. oblongifolia, and C. racemosa (see Table 1). A similar pattern was observed considering species occupancy across plots (Fig. 6). Compared to mature forest, seedling assemblages in small forest fragments exhibited a reduction of two-thirds in the number of species recorded in only one or two plots. Additionally, the three top ten species in mature forest patches were only recorded in 1–3 plots. In small fragments, however, the top ten species were relatively ubiquitous, occurring as adults in 2–17 plots and as seedlings in 2–18 plots. B. virgilioides was the most frequent species in adult assemblages (17 plots), while C. oblongifolia was the most frequent in seedling assemblages (18 plots). Despite the high relative abundance and occupancy exhibited by some pioneer species and a marked reduction in rare species, over 60 % of all pioneer species recorded as adults in small fragments failed to be recorded as seedlings in this habitat type.

Rank-abundance curves for different tree species based on the relative abundance of adults (a, b) and seedlings (c, d) at the Serra Grande landscape. a and c represent stands within the Coimbra mature forest, whereas b and d represent small forest fragments. Key to the top ten proliferating species (i.e., highest SF/MF ratios) for each ontogenetic stage as follows: alfl, Alseis floribunda; arhe, A. heterophyllus; bovi, B. virgilioides; brgu, B. guianense; byse, B. sericea; cuob, C. oblongifolia; cura, C. racemosa; cuim, C. impressinervia; cusp, Cupania sp.; cusp2, Cupania sp. 2; essp2, Eschweileria sp. 2; eusp3, Eugenia sp. 3; eusp4, Eugenia sp. 4; guop, G. opposita; guul, G. ulmifolia; mipr, M. prasina; swma, S. macrostachya

Rank-occupancy curves for different tree species based on the number plots where adults (a, b) and seedlings (c, d) were recorded at the Serra Grande landscape. a and c represent stands within the Coimbra mature forest, whereas b and d represent small forest fragments. Arrows indicate the ten highest SF/MF ratios. Key to the top ten proliferating species (i.e., highest SF/MF ratios) for each ontogenetic stage as follows: alfl, A. floribunda; arhe, A. heterophyllus; bovi, B. virgilioides; brgu, B. guianense; byse, B. sericea; cuob, C. oblongifolia; cura, C. racemosa; cuim, C. impressinervia; cusp, Cupania sp.; cusp2, Cupania sp. 2; essp2, Eschweileria sp. 2; eusp3, Eugenia sp. 3; eusp4, Eugenia sp. 4; guop, G. opposita; guul, G. ulmifolia; mipr, M. prasina; swma, S. macrostachya
Discussion
Tropical forest dynamics cannot be properly addressed without long-term demographic surveys, but for obvious reasons this approach is rarely feasible. We adopted simple measures of population density in seedling and adult tree assemblages to infer on the nature of future floristic transitions and the magnitude of fragmentation-related effects. Our results suggest that a large proportion of tree species are still experiencing altered abundances as seedlings and adults in small forest fragments, even two centuries after primary forest fragmentation and patch isolation. Several tree species are apparently declining while others are proliferating in small forest fragments. Both negative and positive shifts in abundance, rather than occurring randomly, were observed in particular tree functional groups related to regeneration strategy, seed size, and wood density. In general, shade-tolerant representatives of the old-growth flora have declined, while pioneers could respond in both directions. However, most top-ranking species that proliferated as seedlings included both hard and softwood pioneers, and long-lived species inhabiting the forest canopy. Several proliferating taxa that are becoming hyperabundant in small forest fragments present low abundances in mature forest areas of the Coimbra forest, suggesting a release in abundance in edge-dominated forest habitats. These differential responses across several trait-based functional groups largely result in severely altered plant assemblages in small fragments, which are becoming impoverished in both taxonomic and functional terms. Finally, if we accept taxonomic and ecological differences between seedling and adult assemblages in small fragments as a prognosis of future floristic transitions, the future adult tree flora of the ancient Serra Grande forest landscape will likely be radically different from the one today.
Our findings reinforce key notions about fragmentation-related effects on tropical tree species assemblages, several of which are supported by the empirical evidence from Amazonian and other studies in the Atlantic forests. In particular, (1) edge-affected habitats tend to support distorted tree species assemblages in both taxonomic and functional terms (Laurance et al. 2006a, b; Tabarelli et al. 2008), which can be observed also in seedling assemblages (Santo-Silva et al. 2013); (2) small forest fragments are becoming increasingly dominated by pioneer species (Pütz et al. 2011; Tabarelli et al. 2008); and (3) some native pioneer species are proliferating at multiple spatial scales (Tabarelli et al. 2012), particularly B. sericea, B. virgilioides, C. oblongifolia and C. racemosa. Although it has been shown that tree assemblages in small forest fragments are predominantly composed of small-seeded species (Melo et al. 2010; Santo-Silva et al. 2013; Santos et al. 2008), our results suggest that production of large numbers of small seeds in itself does not necessarily confer the ability to proliferate in small fragments.
Finally, we offer new evidence for the establishment of adult tree assemblages dominated by a mix of short- and long-lived successional canopy species that are light-demanding pioneers, which represent only a small fraction of species in undisturbed tropical forest (Laurance et al. 2006b). This more detailed description of the floristic drift experienced by edge-affected habitats support other key working hypotheses: (1) assemblage-wide compositional shifts tend to persist for however long forest environments remain exposed to the open-habitat matrix, suggesting that these shifts are more permanent rather than transitory and (2) edge-dominated forest habitats approach typical conditions of early successional systems, even without clear-cut (Tabarelli et al. 2008).
Although we did not explicitly examine causal mechanisms for the patterns we uncovered in this study, they have been generally considered to be caused by edge effects and seed-dispersal failure. In particular, mechanistic explanations of nonrandom compositional drift are likely related to reduced dispersal movement of large seeds at multiple spatial scales (Cramer et al. 2007; Melo et al. 2006, 2010; Peres and Palacios 2007; Silva and Tabarelli 2000), persistently low seedling recruitment of shade-tolerant old-growth tree species (Santo-Silva et al. 2013), increased mortality of large trees due to uprooting and physiological stress (Laurance et al. 2000), and hyperproliferation of light-demanding species facilitated by high propagule pressure and increased light availability (Tabarelli et al. 2008). These mechanisms are likely to account for the prevalence of canopy, rather than emergent, hard-wooded pioneer tree species among the ten top-ranking SF/MF ratios, such as those from the genus Cupania.
Tree assemblage response to habitat loss and fragmentation, and particularly forest edge creation, in Amazonian forests is consistent with the patterns described here and in other studies for the Atlantic forest. For example, the collapse of the old-growth tree flora and the proliferation of pioneers, particularly short-lived species, have been well documented in Amazonian forest landscapes dominated by both cattle pastures and young secondary forests (Laurance et al. 2006b) and open water (Benchimol and Peres 2015). Based on the patterns found here, we also expect that recently fragmented Amazonian forest landscapes will eventually experience a proliferation of long-lived, canopy pioneers, including hard-wooded species. In other words, patterns documented for ‘old’ and ‘newly’ fragmented landscapes represent different stages of a long continuum of floristic drift experienced by edge-dominated habitats.
On the other hand, these apparently general and widespread patterns are in marked contrast to those documented in other neotropical biotas, such as Lacandona in Mexico, where forest fragments smaller than 100 ha exhibit tree assemblages with species diversity, structure, and relative abundance of pioneer similar to those in continuous forest (Hernandez-Ruedas et al. 2014). Given our rationale, this puzzling contrast may result from a different disturbance history and configuration of these Mesoamerican landscapes. Nevertheless, Amazonian and the Atlantic forests present analogous responses regardless of landscape configuration, time elapsed since forest fragmentation and isolation, and abundance of large-bodied vertebrate seed dispersers (Laurance et al. 2006a, b; Michalski et al. 2007; Santos et al. 2008). We call attention to the evolutionary biogeography of different regions as underlying forces shaping the intensity with which, rather than the direction, different tropical forest settings respond to habitat loss fragmentation. With an evolutionary time exposure to catastrophic natural disturbance events such as hurricanes, plus a long history of human disturbance from pre-Colombian populations, Mexican tropical forest floras may “naturally support” a higher proportion of disturbance-adapted species, thereby exhibiting less pronounced responses to contemporary conversion to nonforest land uses.
Although marked responses of tree assemblages to habitat fragmentation have already been documented, the floristic and functional trajectories experienced by these assemblages over longer isolation time lags remain poorly understood. Tree assemblages inhabiting edge-affected habitats probably experience long-term shifts toward greater dominance of long-lived, pioneer canopy species (Tabarelli et al. 2008). In terms of conservation implications, pioneers and light-demanding species represent only a small fraction of tropical forest floras (Laurance et al. 2006b), although their relative contribution is likely to vary across biotas. This has clear implications to contemporary forest diversity even if only a modest fraction of this ecological group can become hyperabundant in edge-dominated habitats as advocated here. Pioneer-dominated tree assemblages, including those at Serra Grande, have reduced floristic and functional diversity (Santos et al. 2008), reduced size structure (Oliveira et al. 2008), reduced low levels of carbon retention (Paula et al. 2011), reduced diversity of floral attributes (Girão et al. 2007; Lopes et al. 2009), and reduced phylogenetic diversity (Santos et al. 2010). Moreover, pioneer-dominated assemblages have an altered pattern of fruit phenology and much lower relative abundance of trees bearing large fruits (Tabarelli et al. 2010a), which can reduce habitat quality for many frugivorous vertebrates (sensu lato) that play important roles in forest regeneration as seed dispersers and seed predators. Finally, tropical forest ecosystems experiencing high levels of recurrent disturbance tend to provide reduced levels of ecosystems services, including carbon retention and water cycling (Melo et al. 2013). This reinforces the notion that landscapes consisting of small fragments far from large tracts of old-growth forest and surrounded by a cropland matrix represent low conservation value (Melo et al. 2013; Tabarelli et al. 2010b). However, exactly what functional traits ensures species persistence in fragmented landscapes such as Serra Grande are still required, as this question is far from completely elucidated.
References
Arroyo-Rodriguez V, Pineda E, Escobar F, Benitez-Malvido J (2009) Value of small patches in the conservation of plant-species diversity in highly fragmented rainforest. Conserv Biol 23:729–739
Arroyo-Rodriguez V, Cavender-Bares J, Escobar F, Melo FPL, Tabarelli M, Santos BA (2012) Maintenance of tree phylogenetic diversity in a highly fragmented rain forest. J Ecol 100:702–711
Avissar R, Werth D (2005) Global hydroclimatological teleconnections resulting from tropical deforestation. J Hydrometeorol 6:134–145
Benchimol M, Peres CA (2015) Edge-mediated compositional and functional decay of tree assemblages in Amazonian forest islands after 26 years of isolation. J Ecol 103:408–420
Cramer JM, Mesquita RCG, Williamson GB (2007) Forest fragmentation differentially affects seed dispersal of large and small-seeded tropical trees. Biol Conserv 137:415–423
Enquist BJ, West GB, Charnov EL, Brown JH (1999) Allometric scaling of production and life-history variation in vascular plants. Nature 401:907–911
FAO (2010) Global forest resources assessment 2010. Food and Agriculture Organization of the United Nations, Rome
FAO (2011) State of the world’s forests 2011. Food and Agriculture Organization of the United Nations, Rome
Girão LC, Lopes AV, Tabarelli M, Bruna EM (2007) Changes in tree reproductive traits reduce functional diversity in a fragmented Atlantic forest landscape. PLoS One 2:e908
Grillo AS, Oliveira MA, Tabarelli M (2005) Árvores. In: Pôrto K, Almeida-Cortez JS, Tabarelli M (eds) Diversidade biológica e conservação da floresta Atlântica ao norte do rio São Francisco. Ministério do Meio Ambiente, Brasília, pp 191–216
Hartshorn GS (1978) Treefalls and tropical forest dynamics. In: Tomlinson PB, Zimmermann MH (eds) Tropical trees as living systems. Cambridge University, New York, pp 617–638
Hernandez-Ruedas MA, Arroyo-Rodriguez V, Meave JA, Martinez-Ramos M, Ibarra-Manriquez G, Martinez E, Jamangape G, Melo FPL, Santos BA (2014) Conserving tropical tree diversity and forest structure: the value of small rainforest patches in moderately-managed landscapes. PLoS One 9(6):e98931
Joly CA, Metzger JP, Tabarelli M (2014) Experiences from the Brazilian Atlantic forest: ecological findings and conservation initiatives. New Phytol 204:459–473
Laurance WF (2005) When bigger is better: the need for Amazonian mega-reserves. Trends Ecol Evol 20:645–648
Laurance WF, Delamonica P, Laurance SG, Vasconcelos HL, Lovejoy TE (2000) Conservation: rainforest fragmentation kills big trees. Nature 404:836
Laurance WF, Nascimento HEM, Laurance SG, Andrade A, Ribeiro JEL, Giraldo JP, Lovejoy TE, Condit R, Chave J, Harms KE, D’Angelo SA (2006a) Rapid decay of tree-community composition in Amazonian forest fragments. Proc Natl Acad Sci-USA 103:19010–19014
Laurance WF, Nascimento HEM, Laurance SG, Andrade AC, Fearnside PM, Ribeiro JEL, Capretz RL (2006b) Rain forest fragmentation and the proliferation of successional trees. Ecology 87:469–482
Laurance WF, Sayer J, Cassman KG (2014) Agricultural expansion and its impacts on tropical nature. Trends Ecol Evol 29:107–116
Lôbo D, Leão T, Melo FPL, Santos AMM, Tabarelli M (2011) Forest fragmentation drives Atlantic forest of northeastern Brazil to biotic homogenization. Divers Distrib 17:287–296
Lopes AV, Girao LC, Santos BA, Peres CA, Tabarelli M (2009) Long-term erosion of tree reproductive trait diversity in edge-dominated Atlantic forest fragments. Biol Conserv 142:1154–1165
Lorenzi H (1992) Árvores brasileiras. Plantarum, Nova Odessa
Magurran AE (1988) Ecological diversity and its measurement. Cambridge University, Cambridge
Melo FPL, Dirzo R, Tabarelli M (2006) Biased seed rain in forest edges: evidence from the Brazilian Atlantic forest. Biol Conserv 132:50–60
Melo FPL, Martinez-Salas E, Benitez-Malvido J, Ceballos G (2010) Forest fragmentation reduces recruitment of large-seeded tree species in a semi-deciduous tropical forest of southern Mexico. J Trop Ecol 26:35–43
Melo FPL, Arroyo-Rodriguez V, Fahrig L, Martinez-Ramos M, Tabarelli M (2013) On the hope for biodiversity-friendly tropical landscapes. Trends Ecol Evol 28:462–468
Michalski F, Nishi I, Peres CA (2007) Disturbance-mediated drift in tree functional groups in Amazonian Forest fragments. Biotropica 39:691–701
Oliveira MA (2007) Fragmentação e riqueza de árvores em escala local e regional na floresta Atlântica nordestina: implicações para a conservação. Universidade Federal de Pernambuco, Recife
Oliveira MA, Grillo AS, Tabarelli M (2004) Forest edge in the Brazilian Atlantic forest: drastic changes in tree species assemblages. Oryx 38:389–394
Oliveira MA, Santos AMM, Tabarelli M (2008) Profound impoverishment of the large-tree stand in a hyper-fragmented landscape of the Atlantic forest. For Ecol Manage 256:1910–1917
Paula MD, Alves-Costa CP, Tabarelli M (2011) Carbon storage in a fragmented landscape of Atlantic forest: the role played by edge-affected habitats and emergent trees. Trop Conserv Sci 4:349–358
Peres CA (2005) Why we need megareserves in Amazonia. Conserv Biol 19:728–733
Peres CA, Palacios E (2007) Basin-wide effects of game harvest on vertebrate population densities in Amazonian forests: implications for animal-mediated seed dispersal. Biotropica 39:304–315
Pütz S, Groeneveld J, Alves LF, Metzger JP, Huth A (2011) Fragmentation drives tropical forest fragments to early successional states: a modelling study for Brazilian Atlantic forests. Ecol Model 222:1986–1997
R Core Team (2015) R: a language and environment for statistical computing. R Foundation for Statistical Computing, Vienna. http://www.R-project.org/
Ribeiro MC, Metzger JP, Martensen AC, Ponzoni FJ, Hirota MM (2009) The Brazilian Atlantic forest: how much is left, and how is the remaining forest distributed? implications for conservation. Biol Conserv 142:1141–1153
Sala OE, Chapin FS, Armesto JJ, Berlow E, Bloomfield J, Dirzo R, Huber-Sanwald E, Huenneke LF, Jackson RB, Kinzig A, Leemans R, Lodge DM, Mooney HA, Oesterheld M, Poff NL, Sykes MT, Walker BH, Walker M, Wall DH (2000) Biodiversity—global biodiversity scenarios for the year 2100. Science 287:1770–1774
Santos BA, Peres CA, Oliveira MA, Grillo A, Alves-Costa CP, Tabarelli M (2008) Drastic erosion in functional attributes of tree assemblages in Atlantic forest fragments of northeastern Brazil. Biol Conserv 141:249–260
Santos BA, Arroyo-Rodriguez V, Moreno CE, Tabarelli M (2010) Edge-related loss of tree phylogenetic diversity in the severely fragmented Brazilian Atlantic forest. PLos One 5:e12625
Santo-Silva EE, Almeida WR, Melo FPL, Zickel CS, Tabarelli M (2013) The nature of seedling assemblages in a fragmented tropical landscape: implications for forest regeneration. Biotropica 45:386–394
Silva JMC, Tabarelli M (2000) Tree species impoverishment and the future flora of the Atlantic forest of northeast Brazil. Nature 404:72–74
Tabarelli M, Peres CA (2002) Abiotic and vertebrate seed dispersal in the Brazilian Atlantic forest: implications for forest regeneration. Biol Conserv 106:165–176
Tabarelli M, Mantovani W, Peres CA (1999) Effects of habitat fragmentation on plant guild structure in the montane Atlantic forest of southeastern Brazil. Biol Conserv 91:119–127
Tabarelli M, Lopes AV, Peres CA (2008) Edge-effects drive tropical forest fragments towards an early-successional system. Biotropica 40:657–661
Tabarelli M, Aguiar AV, Girao LC, Peres CA, Lopes AV (2010a) Effects of pioneer tree species hyperabundance on forest fragments in northeastern Brazil. Conserv Biol 24:1654–1663
Tabarelli M, Aguiar AV, Ribeiro MC, Metzger JP, Peres CA (2010b) Prospects for biodiversity conservation in the Atlantic forest: lessons from aging human-modified landscapes. Biol Conserv 143:2328–2340
Tabarelli M, Peres CA, Melo FPL (2012) The ‘few winners and many losers’ paradigm revisited: emerging prospects for tropical forest biodiversity. Biol Conserv 155:136–140
van Roosmalen MGM (1985) Fruits of the Guianan flora. Institute of systematic botany, Utrecht
Veloso HP, Rangel-Filho ALR, Lima JCA (1991) Classificação da vegetação brasileira adaptada a um sistema universal. IBGE, Rio de Janeiro
Zanne AE, Lopez-Gonzalez G, Coomes DA, Jansen S, Lewis SL, Miller RB, Swenson NG, Wiemann MC, Chave J (2009) Global wood density database. http://hdl.handle.net/10255/dryad.235
Acknowledgments
We thank Alexandre Grillo (in memoriam), Marcondes Oliveira and Eleno José de Araújo (in memoriam) for plant identification and for making their adult tree database available to us. The Centro de Pesquisas Ambientais do Nordeste (CEPAN) and Usina Serra Grande provided infrastructure and logistic support. We also thank Conservation International—Brazil, Conselho Nacional de Ciência e Tecnologia (CNPq) and Coordenação de Aperfeiçoamento de Pessoal de Nível Superior (CAPES) for research grant to MT, and Fundação de Amparo a Ciência e Tecnologia de Pernambuco (FACEPE) for doctorate fellowship to EESS (process 0455-2.05/09). Finally, we thank CAPES for a doctorate “sandwich” scholarship to EESS (process 17547/12-1).
Author information
Authors and Affiliations
Corresponding author
Additional information
Communicated by Philip Ladd.
Electronic supplementary material
Below is the link to the electronic supplementary material.
Rights and permissions
About this article

Cite this article
Santo-Silva, E.E., Almeida, W.R., Tabarelli, M. et al. Habitat fragmentation and the future structure of tree assemblages in a fragmented Atlantic forest landscape. Plant Ecol 217, 1129–1140 (2016). https://doi.org/10.1007/s11258-016-0638-1
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11258-016-0638-1