
Abstract
In many fire-prone habitats fires may be relatively frequent but of low severity or small areal extent. However, these same habitats may occasionally be subject to large, severe fires when extreme conditions and ignitions coincide. After >50 years without significant fire, a mega-fire burnt >50,000 ha of Eucalyptus–Callitris forest in southeastern Australia. We assessed the impact of this fire on vegetation structure at a landscape scale by quantifying post-fire responses of 11 tree species over 97 sites with varying fire severity. At low severity over 60 % of Callitris trees survived by escaping crown scorch, but they were almost all killed at higher severity. Fewer eucalypts escaped crown scorch (33 % at low fire severity) but there was no evidence of mortality due to the fire. Most eucalypts were topkilled (55 %) but less frequently at low (39 %) compared to moderate (55 %) or high (74 %) fire severity. Larger trees were less likely to suffer topkill. Taken together these results indicate that this wildfire has caused major changes to vegetation structure within the area burnt. Death of Callitris trees reduced canopy tree density by 25 % and a high proportion of eucalypt topkill has resulted in a shorter, more open forest. Recovery of the tallest structural components through eucalypt regrowth and maturation of Callitris may require fire-free intervals of several decades. Any fires within this period may extend the recovery time and lead to declines in populations of the obligate-seeding Callitris species.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
In many fire-prone ecosystems, the occurrence of large and severe fires is constrained by the availability of fuel (e.g. temperate grassy woodlands—Bradstock 2010). In such systems, ignition may be common in the form of lightning but unless spring/early summer conditions are unusually wet, there is insufficient fuel in the ground layer to allow the development of large fires. Hence fires are predominantly small and patchy, with large and high severity wildfires occurring only under extreme weather conditions where discontinuities in ground fuels are overcome by propagation across the canopy of the overstory trees.
Disturbance by fire and post-fire successional processes place these ecosystems on a spectrum with relative proportions of plant species varying according to their response to the previous fire and the time since that disturbance (e.g. Batllori et al. 2015). Prolonged absence of fire may lead to the establishment of species that are susceptible to fire when small, but resilient when large. Once thresholds of relative abundance are exceeded, these ecosystems are resistant to change, with the likelihood of fire reduced by the presence of pyrophobic species that limit ignition, severity or spread of fire (e.g. Cocking et al. 2014; Batllori et al. 2015).
Where a regime of infrequent or of low severity fire has persisted over human generation times, a perception develops that large and severe fires can only be detrimental to ecosystems (DellaSala and Hanson 2015). Indeed, when coupled with habitat fragmentation, large fires may lead to local extinctions of species that rely on dispersal from unburnt patches for persistence. Nevertheless, a lack of knowledge of the effects of large, severe fires prevents broader understanding of their role in ecosystem processes (DellaSala and Hanson 2015). One critical way of alleviating this problem is by examining the effects of fire on the most visually obvious component—the overstory. Since these trees are major drivers of ecosystem processes, this also provides useful ecological information.
Fire severity maps derived from remotely sensed data are commonly used to capture the immediate impact of fires at a landscape scale. However, changes in immediate greenness cannot be used to predict longer-term impacts on ecosystems without knowledge of species or functional group responses (Cocking et al. 2014; Harvey and Holzman 2014). Combining spatially explicit fire severity data with such knowledge allows prediction of the effect of other fires in similar habitats or with similar species composition.
In areas west of the Great Dividing Range in NSW, southeastern Australia, forests are generally dominated by eucalypts that resprout after fire (Myrtaceae: genera Eucalyptus, Angophora, Corymbia), but conifers (Cupressaceae: Callitris endlicheri (J. Thomps. & L. A. S. Johnson) and C. glaucophylla (Parl.) F. M. Bailey) that are obligate seeders (killed by fire) are often codominant. When of sufficient size and density, Callitris reputedly reduce local fire severity via characteristics of their foliage and litter and by reducing the cover of the herbaceous understory (Cohn et al. 2011, 2013; Trauernicht et al. 2012). These Callitris species retain a canopy seed bank (although the temporal dynamics of this are poorly known—Ladd et al. 2013), can recruit in the absence of fire (Ross et al. 2008; Whipp et al. 2012) and are also capable of recruitment into burnt patches from unburnt areas through the dispersal of winged seeds (Cohn et al. 2011).
Although Eucalyptus–Callitris forests are resilient to fire, there is likely to be considerable variation in the response of these forests to the characteristics of particular fire events. Fires of predominantly low severity are likely to have minimal impact on the dominant trees, with many Callitris surviving by escaping crown scorch and eucalypts by epicormic resprouting in the upper branches. However, after high severity (crown) fires, most Callitris are expected to be killed by fire while eucalypts may be topkilled and resprout from buds protected from fire below the soil surface. These contrasting responses have several implications for the post-fire habitat. Firstly, after severe fire, recruitment of Callitris from the canopy seed bank or via dispersal from unburnt patches will be necessary to replace adults killed by the fire. Secondly, the loss of living tissue of eucalypts (upper branches and trunks) will affect the rate of recovery of the canopy and hence the timing and abundance of post-fire reproduction. Thirdly, for understory species, the reduction in abundance or biomass of trees will influence the availability of space for new recruits and the capacity of trees to suppress them. Lastly, these major changes to vegetation structure resulting from severe fire will influence the availability and quality of habitat for dependent fauna, including tree hollows and coarse woody debris (Chambers and Mast 2005; Parnaby et al. 2010; Lindenmayer et al. 2013; Bassett et al. 2015). Hence quantifying the overstory response to fire events of varied severity over a landscape scale will improve understanding of the likely community responses (Belote et al. 2015) and allow informed decisions for post-fire management for conservation.
We used a remotely sensed fire severity map to stratify sampling across local fire severity classes to characterise the response of tree species to a large mixed-severity fire in the Warrumbungle Ranges in January 2013. Eleven dominant tree species were classified into one of the two functional groups with eucalypts predicted to resprout and Callitris species predicted to be killed by crown scorch. For each individual tree we determined whether or not it had been exposed to crown scorch. For eucalypts exposed to crown scorch, we scored whether they were topkilled—resprouting only from basal shoots at the base of the trunk from tissues protected by soil or whether they had epicormic resprouts, where shoots arise from the trunk and upper branches of the tree. In addition, for all trees we measured diameter at breast height as an estimate of tree size, because we predicted that larger trees were more likely to escape crown scorch or if burnt, were more likely to resprout epicormically than smaller ones.
Integrating these data across broadscale vegetation and fire severity classes allowed us to estimate landscape-level change to woody vegetation structure in this ecosystem.
Methods
Study site and study species
The study was undertaken in the Warrumbungle National Park (23,000 ha) on the north-west slopes of New South Wales in southeastern Australia. The region has a climate of warm to hot summers (mean max. 23.7 °C) and mild winters (mean min. 7.4 °C). The mean annual rainfall is 750 mm and, while more rainfall occurs in summer, available soil moisture is higher during the winter months. The topography is mostly mountainous with dissected ranges originating from a shield volcano active some 13 million years ago. Soils are derived from pre-volcanic sandstones, basalts, trachyte and pyroclastic flows. The vegetation is mostly open Eucalyptus-Callitris forest (North-west Slopes Dry Sclerophyll Woodlands and Western Slopes Dry Sclerophyll Forests—Keith 2004) with a variable understory depending on soil fertility (Hunter 2008). Sclerophyllous shrubs dominate the understory where soil fertility is low, and grasses and forbs where soil fertility is higher. Our study species comprise the two dominant conifers C. endlicheri and C. glaucophylla, the dominant eucalypts Eucalyptus crebra F. Muell., E. albens Benth., E. rossii R. T. Baker & H. G. Sm., E. blakelyi Maiden, E. macrorhyncha F. Muell. ex Benth. and Corymbia trachyphloia (F. Muell.) K. D. Hill & L. A. S. Johnson, as well as Angophora floribunda (Sm.) Sweet, E. dealbata A. Cunn. ex Schauer and E. melliodora A. Cunn. ex Schauer which are common in the region (Hunter 2008). There is considerable overlap among canopy-dominant species across vegetation communities, with several species present in most communities. C. glaucophylla, E. albens and E. crebra are the predominant tree species in the area (Tables 1, 2; Hunter 2008).
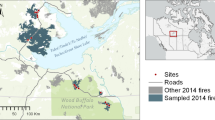
Wildfires have occurred in the area at irregular intervals and fire history records show that most of the study area had not burned for at least 50 years (OEH unpublished data, see OEH 2015). On the 12th–13th January 2013, during a period of hot, dry weather, a fire was initiated and burned over 39,000 ha under extreme fire conditions (Fig. 1). Over the following 2 weeks the Wambelong fire burned a further 17,000 ha under moderate to low fire-weather conditions (BAAT 2013). Approximately 84 % of Warrumbungle National Park was burnt in the fire, including more than half of the reserve at high fire severity (BAAT 2013). The fire burnt across all mapped vegetation communities with a range of fire severities present in each community (Table 1).

Map of the study area showing the fire perimeter, distribution of fire severity classes, National Park reserves and study sites. Inset shows the location of the study area in New South Wales, Australia
Data collection
Sampling was undertaken from April–October 2014, 15–20 months after the fire. Within the burned area, 97 sites were selected where there were 10 or more individuals of one of the 11 target species. We selected 11 tree species that were common in the major vegetation communities of the area, including nine eucalypts and the two obligate-seeding Callitris species (Table 2). Site selection was stratified across fire severity to result in approximately the same number of sites in each of high (23), moderate (23) and low (26) fire severity as well as unburnt (25). Fire severity was determined using a map (Fig. 1, OEH unpublished data) derived from remotely sensed (RapidEye™) data using the relative difference normalised burn ratio (RdNBR) which accounts for different vegetation types (Miller and Thode 2007). Corrections were also made for cloud cover and rockiness. Fire severity classes comprised: Unburnt, RdNBR <4; Low, RdNBR = 4–12 (ground fire, minimal canopy damage); Moderate, RdNBR = 12–21 (understory burnt and canopy mostly scorched); and High, RdNBR >21 (understory burnt and canopy mostly consumed by fire). Sites were also stratified across major lithological groups (sandstone, porphyritic, alluvium and basalt) and to cover variation in aspect and slope.
Since we had no pre-fire data on the relative abundance of trees or the proportion of standing dead stems, unburnt sites were sampled to estimate the likelihood that plants found dead post-fire were dead before the fire. In addition to the minimum 10 individuals of a target species at each site, there were several sites where more than one species were targeted. We also collected data for individuals of any of the study species that occurred in a site, resulting in a total of 5502 individuals. Geographic coordinates and elevation data were recorded from the centre of each site. To account for small-scale variation in mapped fire severity, severity was also visually assessed on-site by the same observer.
At each site, a plot based on a 20 × 20 m quadrat (400 m2) was set up. We measured the diameter at breast height (DBH) and scored the response of each individual of the study species to the fire. These responses could be classified across several binary categories; whether a plant had complete canopy scorch or canopy consumption or not (simplified to burnt or escaped), if burnt whether the plant was alive and if so, whether new growth emerged only from the lignotuber or at base of the trunk (topkilled) or if epicormic growth emerged from the trunk or on branches.
Analysis
To relate the escape of trees from crown scorch to fire severity, tree size and functional group we intended to use Bayesian logistic regression, with random effects for species within functional group. However, from inspecting the raw data it was clear that the disparity in response between the two Callitris species at low fire severity plus the relatively narrower variation between eucalypt species did not support such a model. Another obstacle was the low number of observations for small trees of C. endlicheri within low fire severity plots. We therefore opted for a descriptive analysis, fitting Bayesian credible intervals to the probability of fire escape for combinations of species, tree size and fire severity. We treated the number of escapes as a binomial variable and set a flat Beta distribution as the prior for escape probability. Tree size was expressed as DBH categories: up to 5, 5–10, >10 cm. Intervals were fitted with Markov Chain Monte Carlo using the rjags package for R (Plummer 2015).
At the plot level, we graphed basal area and stem density of Callitris against the proportion of trees that escaped crown scorch to see if there was support for the previously hypothesised severity modifying effects of Callitris trees. This hypothesis (Cohn et al. 2011; Trauernicht et al. 2012) considers that Callitris reduce understory litter flammability and vegetation with the effect size related to the density and or local biomass of Callitris, both of which are expected to be correlated with basal area and stem density.
To assess survival of trees through the fire, we planned to condition our estimate of the proportion of trees that were killed using data on the abundance of dead trees in unburnt plots. However, samples from unburnt sites were found to be strongly biased across species and size distributions and had many more dead individuals for some eucalypt species than any burnt site. Hence we were unable to attribute any dead eucalypts in burnt plots to fire-related mortality. For Callitris, exposure to crown scorch always led to death of the tree. Thus escape from crown scorch (above) can be used to estimate survival for this group.
For eucalypts that were exposed to crown scorch, we used Bayesian logistic regression, fitted by MCMC using the rjags package for R (Plummer 2015), to relate the proportion of trees that were topkilled (i.e. only had basal resprouts in contrast to epicormic resprouts) to fire severity and tree size, with random effects for species. Tree size was expressed in DBH categories as above. We set vague Normal prior distributions for all regression coefficients. Species random effects were drawn from Normal distributions with zero mean and a standard deviation specific to each fire severity category.
Results
Escape from crown scorch
We found a strong effect of fire severity on the likelihood that trees escaped crown scorch with very few trees escaping at moderate and high severity (Fig. 2). At low fire severity, the likelihood of escape was highly variable among species and size classes, while at higher severity the range and variability was lower. There was no clear influence of functional group apparent in the fitted intervals for probability of escape, but C. glaucophylla had notably greater likelihood of escape at low severity, especially in smaller size classes. In contrast to all other species, large trees of E. rossii maintained high likelihood of escape (greater than 50 %) at moderate fire severity. The relative composition of plots did not appear to influence likelihood of escape from crown scorch, i.e. we found no indication at the plot level of Callitris density or stem basal area reducing flammability (Fig. 3).

Proportion of trees that escaped crown scorch arranged by species, diameter at breast height (DBH) size class and fire severity. Data are fitted probabilities and credible intervals

Proportion of all trees that escaped crown scorch at the plot level in relation to fire severity and a Callitris basal area and b Callitris stem density
Survival through fire
For the two Callitris species, exposure to crown scorch leads to mortality, hence trees that did not escape fire were killed (Fig. 4, left-most species). In contrast, for the nine eucalypt species there were few dead individuals observed in burnt plots while considerably more were observed in unburnt plots (Fig. 4, top left). This disparity left no option but to conclude that there was negligible or no fire-caused mortality in eucalypts.

Occurrence of dead trees across species, diameter at breast height (DBH) and fire severity
Resprouting responses of eucalypts after fire
For eucalypts that were exposed to crown scorch, fire severity, tree size and their interaction all significantly influenced the likelihood of topkill (Fig. 5; Table 3). There was a much greater probability of topkill at high fire severity compared to moderate or low fire severity and larger trees were less likely to be topkilled (Fig. 6). Hence larger trees and those burnt at lower severity were more likely to resprout from epicormic buds on the trunk or upper branches. The influence of tree size on the likelihood of topkill was stronger at moderate and high severity. Among species, A. floribunda was less likely to be topkilled across all fire severities, while E. albens and E. crebra were more likely to be topkilled at high severity and E. dealbata was more likely to be topkilled at low severity (Fig. 5).

Proportion of topkilled trees (eucalypts only) arranged by species, diameter at breast height (DBH) size class and fire severity

Predicted probability of topkill in relation to diameter at breast height (DBH) size class and fire severity for eucalypt species pooled
Discussion
Escape from crown scorch
Crown scorch (dead foliage and branches) results from two components of the fire: (i) patchiness which determines whether or not a plant is burnt or heated; and (ii) when a plant is burnt, the local intensity of the fire which influences how much heat stress it experiences (Balfour and Midgley 2006; Ooi et al. 2006). Fire severity is expected to be correlated with both these measures such that heat stress during a fire is lowest and patchiness is greatest at low fire severity (Penman et al. 2007; Leonard et al. 2014). We found a strong influence of fire severity on the likelihood that individual plants escaped crown scorch and at low fire severity, an increased likelihood of escape for larger trees across all species. Very few plants escaped from crown scorch at moderate or high fire severity. There was not a clear functional group separation in escape probability due to the apparent differences between Callitris species at low fire severity exceeding the range of variation of eucalypt species. At low fire severity, both observed proportions and estimated probability of escape in C. endlicheri were low for small- and medium-sized plants. In contrast, C. glaucophylla showed a strong capacity to escape from low severity fire across the range of size classes, consistent with other studies of this and other coniferous species (e.g. Callitris—Cohn et al. 2011; Trauernicht et al. 2012; Larix occidentalis—Belote et al. 2015; Pseudotsuga menziesii—Cocking et al. 2012). These studies indicate that these conifers alter the fuel characteristics in their immediate vicinity by suppressing understory vegetation and by having litter that is less flammable than angiosperm litter. At the individual level we found support for this model but, it was not supported at the plot level. Despite covering a range of basal area encompassing that considered by Cohn et al. (2011) to characterise Callitris patches (approximately 0.13 m2 per 100 m2), there was no clear relationship between Callitris basal area and the overall proportion of trees that escaped from crown scorch even at low fire severity (Fig. 3a). Escape from crown scorch was the only means for individuals of Callitris species to survive fire.
For eucalypt trees capable of resprouting, exposure to crown scorch has minimal effect on survival. However, escaping crown scorch reduces the impact of the fire on living tissue and hence size structure by preventing loss of canopy (Dodonov et al. 2014). For all eucalypts combined, 13.3 % escaped crown scorch comprising low severity (11.6 %), moderate severity (1.6 %) and high severity (0.1 %). In contrast to all other species, large trees of E. rossii had a greater than 50 % likelihood of escaping crown scorch at moderate severity. Whether this reflects characteristics of the species or differences in its microhabitat preferences warrants further investigation. This difference was not apparent at high severity, where all species had very low probability of escape.
Survival through fire
For trees that did not escape crown scorch, survival was dependent on functional group—the obligate-seeding Callitris were all killed by fire. Given the relative abundance of Callitris across all plots, death of Callitris trees reduced canopy tree density by 25 %, mostly concentrated in areas of moderate and high fire severity. In these areas, re-establishment of Callitris will rely on seedling recruitment either from the canopy seed bank of killed plants or via seed dispersal from surviving plants in low severity or unburnt areas.
Despite observations of dead eucalypts in burnt plots, based on our samples of unburnt plots, we concluded that there was negligible or no eucalypt mortality due to fire. The relatively high proportion of standing dead trees in unburnt plots suggests that the dead trees observed after the fire may simply be pre-existing dead trees that were not consumed by the fire. Near zero losses through fire for adult eucalypt species are commonly assumed and have been observed elsewhere (e.g. Bennett et al. 2013) although others have reported significant mortality in resprouting eucalypts through similar fires (e.g. 7.5 % Strasser et al. 1996; Benyon and Lane 2013).
Resprouting responses of eucalypts after fire
For eucalypts that were exposed to fire, the resprouting response (epicormic or basal) is critical in determining the level of structural change at a landscape scale. Despite the prominent role of eucalypts in fire-prone ecosystems in Australia (e.g. Clarke et al. 2015), quantification of degree of topkill is surprisingly rare for eucalypt forests outside the tropical savanna. We observed 55 % topkill for nine eucalypt species pooled across the range of fire severities, with more than 70 % topkilled at high fire severity. There was a strong size class effect and considerable variation in topkill probability among species, with some species (e.g. A. floribunda) having more epicormic resprouting individuals across all fire severity classes. Strasser et al. (1996) observed a similar influence of tree size on likelihood of topkill for several Eucalyptus species in high altitude forests in southern Australia, with less than 60 % topkilled overall. In tropical Brazil, Hoffmann et al. (2009) found that forest (52 %) and savanna (41 %) species were topkilled after fire, despite lower fire severity in the forested area. Similarly, Hoffmann and Solbrig (2003) found relationships between fire severity, DBH and probability of topkill for several Brazilian savanna trees, with probabilities in the range 40–80 % for the largest trees at high severity. The proportion of trees that are topkilled affects the relative dominance of individual trees or species within a stand and their influence on other plants and dependent fauna. A topkilled tree may take longer to reproduce and the loss of upper branches will likely reduce their post-fire fecundity and re-establishment of a canopy seed bank. The mean height of trees was 10.3 m (n = 1370, range 1–26 m); hence there has been a substantial reduction in taller canopy plants after the fire, particularly in high severity locations.
Overall impact of the Wambelong fire
Combining data from each of our analyses with data on local abundance (Table 2) and considering the area burnt by each fire severity class (Table 1) allows a broad overall assessment of the impact of the fire. For example, for the White box—Ironbark—White pine community, the predominant community across the region, 34 % or almost 11,000 ha was burnt at high fire severity. At that severity the likely impact on the three most common species (E. albens, E. crebra and C. glaucophylla) was topkill for over 75 % of the eucalypts and mortality for all the Callitris trees, leading to a major alteration of structure and a change in dominance among the canopy species of this landscape. Death of adult and juvenile Callitris trees removes any severity ameliorating influence they may have across the landscape for several decades until post-fire seedlings reach a size or canopy density where they have some effect (Cohn et al. 2011). A fire-free period of at least 10 years will be necessary to allow Callitris seedlings to reach maturity (Lunt et al. 2011). Fires of any severity within that period may lead to local extinctions of Callitris and possibly allow increased establishment of the more flammable eucalypt species. Hence, while this fire has caused significant alteration of forest structure and relative abundance of tree species, it is the future fire regime that will greatly influence the trajectory of the assemblage of overstory species. This scenario is analogous to oak-conifer systems in western United States, where conifers (e.g. Abies concolor, Pseudotsuga menziesii) dominate long unburnt forests, while moderate to high frequency fire allows persistence of resprouting oaks such as Quercus kelloggii (Cocking et al. 2012, 2014). However, unlike the American situation, information on the rate of population increase of Callitris in the absence of fire is largely qualitative (e.g. Austin and Williams 1988; Clayton-Greene and Ashton 1990; Cohn et al. 2011; Whipp et al. 2012) and we found no evidence that C. endlicheri, which dominates the slopes and poorer soils of the region, had similar fire suppression or avoidance strategies characteristic of C. glaucophylla or A. concolor and P. menziesii.
Conclusions
This study has characterised the impact of a large, mixed-severity fire on several resprouting eucalypt species and two fire-sensitive Callitris species. A substantial proportion of Callitris trees survived by escaping crown scorch at low fire severity, but most were killed at moderate and high severity. In contrast, although fewer eucalypt trees escaped crown scorch, most resprouted after fire with the manner of resprouting strongly influenced by fire severity. In addition, there were consistent influences of tree size on fire impacts, with larger trees more likely to escape crown scorch or resprout epicormically than smaller ones. These findings, together with knowledge of tree density and size distributions provide a basis on which to predict fire-vegetation dynamics at the landscape level. In particular, it is clear that fires within the next decade would likely cause declines in Callitris as well as further loss of canopy structure and potentially, loss of biodiversity.
References
Austin MP, Williams OB (1988) Influence of climate and community composition on the population demography of pasture species in semi-arid Australia. Vegetatio 77:43–49
Balfour DA, Midgley JJ (2006) Fire induced stem death in an African acacia is not caused by canopy scorching. Austral Ecol 31:892–896
Bassett M, Chia EK, Leonard SWJ, Nimmo DG, Holland GJ, Ritchie EG, Clarke MF, Bennett AF (2015) The effects of topographic variation and the fire regime on coarse woody debris: insights from a large wildfire. For Ecol Manag 340:126–134
Batllori E, Ackerly DD, Moritz MA (2015) A minimal model of fire-vegetation feedbacks and disturbance stochasticity generates alternative stable states in grassland-shrubland-woodland systems. Environ Res Lett 10:034018
Belote RT, Larson AJ, Dietz MS (2015) Tree survival scales to community-level effects following mixed-severity fire in a mixed-conifer forest. For Ecol Manag 353:221–231
Bennett LT, Aponte C, Tolhurst KG, Loew M, Baker TG (2013) Decreases in standing tree-based carbon stocks associated with repeated prescribed fires in a temperate mixed-species eucalypt forest. For Ecol Manag 306:243–255
Benyon RG, Lane PNJ (2013) Ground and satellite-based assessments of wet eucalypt forest survival and regeneration for predicting long-term hydrological responses to a large wildfire. For Ecol Manag 294:197–207
Bradstock RA (2010) A biogeographic model of fire regimes in Australia: current and future implications. Glob Ecol Biogeogr 19:145–158
Burned Area Assessment Team (BAAT) (2013) Post-fire rapid risk assessment and mitigation: Wambelong fire, Warrumbungle National Park. Unpublished report to NSW National Parks and Wildlife Service, Hurstville
Chambers C, Mast J (2005) Ponderosa pine snag dynamics and cavity excavation following wildfire in northern Arizona. For Ecol Manag 216:227–240
Clarke PJ, Lawes MJ, Murphy BP, Russell-Smith J, Nano CEM, Bradstock R, Enright NJ, Fontaine JB, Gosper CR, Radford I, Midgley JJ, Gunton RM (2015) A synthesis of postfire recovery traits of woody plants in Australian ecosystems. Sci Total Environ 531:31–42
Clayton-Greene K, Ashton D (1990) The dynamics of Callitris columellaris/Eucalyptus albens communities along the Snowy River and its tributaries in south-eastern Australia. Aust J Bot 38:403–432
Cocking MI, Varner JM, Sherriff RL (2012) California black oak responses to fire severity and native conifer encroachment in the Klamath Mountains. For Ecol Manag 270:25–34
Cocking MI, Varner JM, Knapp EE (2014) Long-term effects of fire severity on oak-conifer dynamics in the southern Cascades. Ecol Appl 24:94–107
Cohn JS, Lunt ID, Ross KA, Bradstock RA (2011) How do slow-growing, fire-sensitive conifers survive in flammable eucalypt woodlands? J Veg Sci 22:425–435
Cohn JS, Lunt ID, Bradstock RA, Hua Q, McDonald S (2013) Demographic patterns of a widespread long-lived tree are associated with rainfall and disturbances along rainfall gradients in SE Australia. Ecol Evolut 3:2169–2182
DellaSala DA, Hanson CT (2015) The ecological importance of mixed-severity fires. Elsevier, Netherlands
Dodonov P, de Oliveira Xavier R, dos Santos Tiberio FC, de Lucena IC, Brandão Zanelli C, da Silva Matos DM (2014) Driving factors of small-scale variability in a savanna plant population after a fire. Acta Oecol 56:47–55
Harvey BJ, Holzman BA (2014) Divergent successional pathways of stand development following fire in a California closed-cone pine forest. J Veg Sci 25:88–99
Hoffmann W, Solbrig O (2003) The role of topkill in the differential response of savanna woody species to fire. For Ecol Manag 180:273–286
Hoffmann WA, Adasme R, Haridasan M, de Carvalho MT, Geiger EL, Pereira MAB, Gotsch SG, Franco AC (2009) Tree topkill, not mortality, governs the dynamics of savanna-forest boundaries under frequent fire in central Brazil. Ecology 90:1326–1337
Hunter JT (2008) Vegetation and floristics of Warrumbungle National Park. Report to NSW National Parks and Wildlife Service, Coonabarabran
Keith DA (2004) Ocean shores to desert dunes: the native vegetation of New South Wales. Department of Environment and Conservation (NSW), Hurstville
Ladd PG, Midgley JJ, Nield AP (2013) Serotiny in southern hemisphere conifers. Aust J Bot 61:486–496
Leonard SWJ, Bennett AF, Clarke MF (2014) Determinants of the occurrence of unburnt forest patches: potential biotic refuges within a large, intense wildfire in south-eastern Australia. For Ecol Manag 314:85–93
Lindenmayer DB, Blanchard W, McBurney L, Blair D, Banks SC, Driscoll D, Smith AL, Gill AM (2013) Fire severity and landscape context effects on arboreal marsupials. Biol Conserv 167:137–148
Lunt ID, Zimmer HC, Cheal DC (2011) The tortoise and the hare? Post-fire regeneration in mixed Eucalyptus-Callitris forest. Aust J Bot 59:575–581
Miller JD, Thode AE (2007) Quantifying burn severity in a heterogeneous landscape with a relative version of the delta normalized burn ratio (dNBR). Remote Sens Environ 109:66–80
OEH (2015) Fire management manual 2015-2016: policy and procedures for fire management. NSW Office of Environment and Heritage, Sydney. http://www.environment.nsw.gov.au/resources/firemanagement/final/140397FireManManual.pdf. Accessed 8 Feb 2016
Ooi MKJ, Whelan RJ, Auld TD (2006) Persistence of obligate-seeding species at the population scale: effects of fire intensity, fire patchiness and long fire-free intervals. Int J Wildland Fire 15:261–269
Parnaby H, Lunney D, Shannon I, Fleming M (2010) Collapse rates of hollow-bearing trees following low intensity prescription burns in the Pilliga forests, New South Wales. Pac Conserv Biol 16:209–220
Penman TD, Kavanagh RP, Binns DL, Melick DR (2007) Patchiness of prescribed burns in dry sclerophyll eucalypt forests in South-eastern Australia. For Ecol Manag 252:24–32
Plummer M (2015) rjags: Bayesian graphical models using MCMC. R package version 3-15. http://CRAN.R-project.org/package=rjags
Ross KA, Bedward M, Ellis MV, Deane A, Simpson CC, Bradstock RA (2008) Modelling the dynamics of white cypress pine Callitris glaucophylla woodlands in inland south-eastern Australia. Ecol Model 211:11–24
Strasser M, Pausas J, Noble I (1996) Modelling the response of eucalypts to fire, Brindabella ranges, ACT. Aust J Ecol 21:341–344
Trauernicht C, Murphy BP, Portner TE, Bowman DMJS (2012) Tree cover-fire interactions promote the persistence of a fire-sensitive conifer in a highly flammable savanna. J Ecol 100:958–968
Whipp RK, Lunt ID, Spooner PG, Bradstock RA (2012) Changes in forest structure over 60 years: tree densities continue to increase in the Pilliga forests, New South Wales, Australia. Aust J Bot 60:1–8
Acknowledgments
We thank Michael Bedward for providing formidable statistical analysis and advice of the highest quality and at times well beyond the normal call of duty. Michael Bedward and an anonymous reviewer also made helpful comments on earlier drafts of the manuscript.
Author information
Authors and Affiliations
Corresponding author
Additional information
Communicated by Michael Lawes, Ross Bradstock and David Keith.
Rights and permissions
About this article

Cite this article
Denham, A.J., Vincent, B.E., Clarke, P.J. et al. Responses of tree species to a severe fire indicate major structural change to Eucalyptus–Callitris forests. Plant Ecol 217, 617–629 (2016). https://doi.org/10.1007/s11258-016-0572-2
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11258-016-0572-2