
Abstract
We conducted a field investigation and controlled experiments in a glasshouse to determine how litter, seed position, and/or water supply affect seedling emergence and the establishment of two native grass species (Leymus chinensis (Trin.) Tzvel. and Bromus inermis Leyss) in northern China. Both water supply and seed position dramatically influenced seedling emergence and growth, as well as the accumulation of biomass for these two species. For L. chinensis, the quantity of litter had a significant impact on seedling survival, height increment, and biomass production when grown in either the glasshouse or the field while that component had little influence on B. inermis seedling emergence or survival. The effects of litter were physical but not biological or chemical, suggesting that this component is important only in the first year of seedling establishment. Therefore, longer-term experiments are probably necessary to examine the biological and chemical impacts that litter has on seedling performance.
Similar content being viewed by others


Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
Cessation of grassland management, a reduction in grazing pressure, or the installation of fencing results in litter accumulation, which leads to a decline in species diversity because of the decreased availability of open patches (Rotundo and Aguiar 2005; Bissels et al. 2006). This accumulation of plant litter can regulate seedling recruitment both positively (Eckstein and Donath 2005; Rotundo and Aguiar 2005; Loydi et al. 2013) and negatively (Xiong and Nilsson 1999; Jensen and Meyer 2001). The extent of these effects is related to the level of existing litter (Hovstad and Ohlson 2008; Ruprecht et al. 2008; Eckstein et al. 2012; Wellstein 2012; Loydi et al. 2013), with small or moderate amounts positively influencing recruitment on dry grasslands or under water-limiting conditions (Eckstein and Donath 2005; Rotundo and Aguiar 2005; Eckstein et al. 2012). However, high litter densities (>500 g m−2) can inhibit recruitment (Loydi et al. 2013). Thus, the most important constraint when trying to establish new seedlings at many study sites appears to be a lack of amenable locations that encourage germination due to an overly thick layer of litter (Jensen and Meyer 2001; Jensen and Gutekunst 2003; Ruprecht et al. 2010; Miglécz et al. 2013). This phenomenon varies considerably among species within a community as well as across microhabitats in a particular landscape (Hovstad and Ohlson 2008). Unfortunately, there is a bias of published papers in favor of North American and European grasslands and temperate biomes, while patterns of response in other regions, especially in Asia, have been poorly reported. Therefore, more efforts should be undertaken to study the effects of litter in those types of ecosystems (Loydi et al. 2013).
Here, we recorded how two typical, semiarid native grass species, Leymus chinensis (Trin.) Tzvel. and Bromus inermis Leyss in northern China respond to such litter accumulations. L. chinensis is a rhizomatous grass, and B. inermis is an erect rhizomatous grass, both grass species are C3 perennials with high productivity and forage value (Jia 1987). B. inermis is distributed in northern China and L. chinensis is distributed widely in East Asia, including China, Japan, Mongolia, and eastern Russia (Xiao et al. 1995; Wang and Ripley 1997). Grasslands dominated by L. chinensis are among the most important grazing and mowing pastures in China (Wang et al. 2004). They are critical to vegetation rehabilitation in that area because of their high forage and ecological values (Gao 2004; Wang et al. 2004; Zhao and Sun 2004). However, little is known about how litter density and the availability of water influence seedling establishment, which ultimately determines how successfully these grassland ecosystems can be restored. Our primary study objective was to evaluate the effects of litter amounts during the first year in which both species are being established. We specifically addressed the following hypotheses:
-
(1)
the influence of litter on seedling emergence is predominantly positive under dry conditions but negative or neutral under a favorable water supply;
-
(2)
physical factors are the main for short term and, possibly, facilitative determinants of how litter cover modulates seedling emergence, with the rate of establishment decreasing as the amount of litter increases;
-
(3)
litter affects seed positioning in a natural environment, with seed lying on the litter surface, between the litter and the soil, or else buried within the soil.
Materials and methods
Site description
Field experiments were conducted at the National Field Station of the Grassland Ecosystem (SaiBei) (NFSGE) (41º45′–41°57′N, 115°39′–115°48′E, elev: 1400 m), Saibei administrative region, Hebei Province, China. This site is located in the southern portion of the Xilingol steppe grassland where the climate is semiarid and the growing season is 100 days long. Mean annual precipitation is 398.8 mm and mainly occurs in July, August, and September. The mean annual temperature is 1.9 °C, and the accumulated temperature ≥10 °C is 1513.1 °C. Annual average wind speed is 4.3 m s−1 and the annual sunshine duration is 2930.9 h. The main soil type, Chestnut, is slightly alkaline (pH 7.7).
Observations were also made at the NFSGE glasshouse, where the mean temperature was 23.8 °C, mean humidity was 60.0 %, and the annual sunshine duration was the same as at the field site. All data were collected with an automatic HOBO temperature and humidity apparatus (H12-002; Onset Computer Corporation, USA). Humidity in the glasshouse was controlled via wet shading, and the cooling system maintained the interior temperature between 7 °C (= minimum outdoor temperature) and 25 °C.
Seed sources
Seeds of L. chinensis and B. inermis were collected from the surrounding region in September of 2004 and 2005. They were then dry-stored in the dark at room temperature until June 2006, when they were sorted with a blowing machine prior to sowing. The mean seed masses were 2.41 mg for L. chinensis and 3.91 mg for B. inermis. Average germination rates in laboratory Petri dishes (at 20–30 °C) were 79.5 and 94.5 % for L. chinensis and B. inermis, respectively.
Field experiments
For the field tests, the effects of litter and seed position on seedling emergence and early establishment were studied in 60 plots (c. 30 × 30 cm) per species (total of 120 plots). Dead plant material lying loose on the ground (Facelli and Pickett 1991) was collected and then re-distributed at densities of 0 g m−2 (no litter), 50 g m−2 (low), 100 g m−2 (average), or 200 g m−2 (high), which corresponded to 0.0, 4.5, 9.0, or 18.0 g of litter per plot (the density of the litter that was actually applied was calculated based on a typical, mean field weight of 100 g m−2) (cf., Rotundo and Aguiar 2005; Eckstein and Donath 2005). Some seeds were placed directly on the soil and covered by litter (“between” treatment) or else vertically inserted into the soil at a 2-cm depth and then covered by litter (“buried”). For the “surface” treatment, the seeds were placed on top of the litter layer (i.e., no soil–seed contact) that corresponded to the low, average, or high litter densities. The treatments were assigned to plots in a randomized block design, with 50 seeds sown per plot spaced 50 cm apart. Prior to sowing, all of the existing plants and litter were removed from the plots to eliminate any competition. All seeds were planted in a single day.
Glasshouse experiments
The glasshouse tests comprised 144 pots (20-cm diam./16 cm tall, surface area of 0.0314 m2) per species, for a total of 288 pots. All pots were filled with field soil that had been passed through a 0.5-mm sieve to eliminate seeds, stones, litter, and other extraneous materials. The soil (pH 7.7) was supplemented with 0.26 % ammonia nitrogen, 4.2 mg kg−1 P2O5, and 179.6 mg kg−1 K2O. Each pot contained 50 seeds, all of which were sown in a single day. A four-way factorial design was used to assess the role of litter type as a component along with litter quantity, water supply, and vertical seed position. Either naturally sourced litter (collected as described above for the field tests) or plastic fibers were used that simulated the size and shape of natural litter. Because plastic neither releases nutrients nor produces allelopathic compounds (cf., Facelli and Pickett 1991), it was considered a contributor to the physical effects on seedling establishment, similar to the influence of an increased water supply. In contrast, chemical effects, e.g., nutrient release, contributed to the positive effects associated only with natural litter while biological effects included pathogen infections from spores carried by the litter. Both natural and plastic litters were applied at densities of 0 g m−2 (no litter), 50 g m−2 (low), 100 g m−2 (average), or 200 g m−2 (high), which corresponded to 0.0, 1.6, 3.2, or 6.4 g of litter per pot. Two levels of water supply were also tested: 50 mm per month (low = 157 mL per pot) or 100 mm per month (high = 314 mL per pot). Those levels represented the lowest and highest mean monthly amounts of precipitation based on data collected at the local weather bureau since 2003. All pots were watered at their assigned irrigation level at 3-day intervals. Treatments for seed positions were again “surface,” “between,” and “buried.” A split block experimental design was used for arranging the pots.
Data collection
In 2006, seeds were sown on 18 June in the field and on 5 July in the glasshouse. The rates of seedling emergence per treatment were recorded on 7, 14, and 21 July, and 1 and 8 August in the field and on 16, 19, 22, and 29 July, and 5 and 12 August in the glasshouse. The seedlings were marked with non-toxic, colored toothpicks to differentiate newly emerged seedlings from those already present. Because only seedlings that went through the litter layer could become established, we considered only those seeds that had successfully germinated when calculating the percentage emergence for each date (Eckstein and Donath 2005).
Final seedling survival was counted in the field on 8 September and in the glasshouse on 12 September. Shoot heights were measured on 23 August in the field and on 27 August in the glasshouse. This allowed us to determine growth rates, based on 5–10 seedlings randomly selected from each treatment type. On 15 September, the total aboveground biomass was harvested in the glasshouse, then dried to a constant mass at 65 °C, and weighed. The root masses were also removed from each pot, then washed, dried to a constant mass at 65 °C, and weighed.
Analysis
The percentages of seedling emergence and survival were log-transformed to achieve normality and homogeneity of variances. Means were back-transformed to report the data. Two-way ANOVA was used for evaluating the effects of litter quantity and seed position in the field. A GLM Univariate ANOVA was conducted to examine the influence of seed position, litter quantity, water supply, and litter type on dependent variables in the glasshouse tests. Those variables included final percentage emergence, survival, growth rate (i.e., change in seedling height over time), and the biomass of both above- and belowground tissues as well as total production per pot. All factors were considered fixed. Repeated Measures in GLM ANOVA (SPSS, Chicago, IL, USA) were performed to test the effect of sampling date and litter treatment on soil moisture. Type III Sums of Squares were used for calculating F statistics, and Tukey’s tests were conducted for post hoc comparisons. All statistical analyses utilized SPSS 17.0.
Results
Field experiments
Seedling emergence
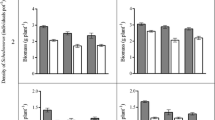
The percentage of L. chinensis seedlings that emerged was significantly affected by seed position, litter quantity, and their interaction (Table 1). Values calculated from the buried in soil treatment were 4.8-fold higher than for the litter-surface treatment and 1.8-fold higher than for the between (soil covered with litter) treatment (Fig. 1a). Finally, a significantly higher percentage of seedlings emerged from soil without litter cover than from any treatment in which the seeds had been covered (Fig. 1b).

Effects of seed position (a) and litter quantity (b) on emergence percentages for field-grown seedlings of Leymus chinensis. For each panel, bars not labeled with the same letter indicate significantly different values at (p < 0.05, based on) (Tukey’s tests)
The emergence percentages for B. inermis were significantly affected only by seed position (Table 1). Values for seeds initially buried in the soil were 12.0 % higher than for those placed on the litter surface and 12.7 % higher than for those within the litter layer (Fig. 2a). The amount of litter had no impact on emergence percentages for this species (Fig. 2b).

Effects of seed position (a) and litter quantity (b) on emergence percentages for field-grown seedlings of Bromus inermis. For each panel, bars not labeled with the same letter indicate significantly different values at p < 0.05, based on Tukey’s tests
Seedling survival
Survival of L. chinensis seedlings was significantly affected only by litter quantity, with rates being nearly two-fold higher in average and high litter treatments than in no-litter and low-litter treatments (Tables 1, 2). By contrast, rates for B. inermis were not significantly influenced by seed position, litter quantity, or their interaction.
Glasshouse experiments
Soil moisture and light quality
Figure 3 shows how soil moisture levels changed over time after pots containing different densities of litter layers were watered. Regardless of species or irrigation treatment, i.e., 100 or 50 mm per month, moisture contents tended to decrease during the 4 days after water was applied. However, when compared with the control treatment (no litter), the rate at which the soil surface dried was negatively correlated with the amount of litter that was present. This indicated that litter had an obviously positive effect on the retention of soil moisture (p < 0.01).

Effects on soil moisture content during first 4 days after water was supplied to seedlings at rates of 100 mm/month (a) or 50 mm/month (b)
Seedling emergence
Three main factors—seed position (S), water supply (W), and litter type (T)—significantly affected the emergence percentage for glasshouse-grown L. chinensis, while litter quantity (L) had no significant influence. The interactions of S × L, S × W, L × W, L × T, S × T, S × L × W, S × W × T, L × W × T, and S × L × W × T also produced significant effects (Table 3).
For this species, the percentage of emergence from the buried treatment was 37.9 % higher than from the surface treatment and 51.9 % higher than from the “between” treatment. Values were also significantly higher (15.5 %) for treatments receiving more water (100 mm/month) than for the low-water supply treatment (50 mm/month). Moreover, emergence percentages were significantly greater from plastic litter than from the natural grass litter. Litter quantity had a negative influence on seedling emergence, with percentages being higher when no litter was present than when the soil surface was covered (Table 4). However, among treatments involving some amount of litter, those percentages did not differ significantly.
For glasshouse-grown B. inermis, the emergence percentages were significantly affected by seed position and litter type, but not by litter quantity or level of water supplied (Table 3). The interactions of S × L, S × W, L × W, S × T, and L × T were also significant. Values were significantly higher for initially buried seeds than for seeds that had been placed either on top of the litter surface or between the litter and the soil. Percentages were also higher for treatments with plastic litter than for those testing natural grass litter. Water supply and litter quantity had only a small influence on seedling emergence (Table 5).
Seedling growth
The heights of L. chinensis seedlings in the glasshouse were significantly affected by seed position, litter quantity, water supply, and litter type; the interactions of S × L, L × W, L × T, and L × W × T affected this growth parameter significantly (Table 3). Seedlings that developed from buried seeds were significantly larger than those from other treatments, i.e., 7.0 cm or 6.5 cm taller than seedlings emerging from the surface or “between” treatments, respectively. Seedlings that received more water were also significantly taller than those in the low-water supply treatments. Moreover, covering the soil with plastic litter resulted in seedlings that were 5 cm taller than those in pots where grass litter was used.
For glasshouse-grown B. inermis, heights were significantly affected by seed position, water supply, and litter type, but not by litter quantity or any of those interactions (Table 3). In particular, seedlings grown from buried seeds were 3 cm taller than those from either the surface or “between” treatments (Table 4). Furthermore, values for height were greater for seedlings in the high-water supply treatment as well as those where the soil surface was covered with plastic litter rather than grass litter.
Seedling survival
Survival of L. chinensis in the glasshouse was significantly affected by litter quantity and type (Table 3). Rates were significantly lower for seedlings that had no litter cover. Survival was increased by 16.8, 18.4, or 16.9 % when the soil surface was covered with 50, 100, or 200 g m−2 litter. The type of litter also influenced survival, with rates being higher in pots topped with plastic material. By contrast, seed position and water supply did not significantly affect survival by that species. However, the interactions of S × W, L × T, and S × L × W were significant.
For B. inermis, survival was not significantly affected by these main factors, and only the interaction of seed position × litter quantity had a significant influence on survival (Table 3).
Seedling biomass
The total biomass, aboveground biomass, and belowground biomass of L. chinensis were significantly affected by seed position and litter quantity (Table 3). Values for total biomass of seedlings obtained from buried seed were 2.1 and 1.8 times higher when compared with the surface and “between” treatments. The aboveground biomass produced by seedlings from buried seeds was approximately twice as great as for plants in the other two positional treatment. A similar trend was noted for belowground biomass. Total biomass was also significantly and positively correlated with litter quantity and the amount of water supplied. Whereas the aboveground biomass was significantly affected by water supply and litter quantity, the level of irrigation had no significant impact on belowground biomass (Table 4). The interactions of S × L, L × W, L × T, and S × W × T had significant effects on aboveground biomass, belowground biomass, and total biomass for L. chinensis, while the W × T interaction influenced both aboveground and total biomass. Those latter two parameters were also significantly affected by the interactions of S × L × W and S × T (Table 4).
For B. inermis, total, aboveground, and belowground biomasses were significantly affected by seed position while the aboveground biomass was significantly influenced by water supply and litter type. Moreover, the interactions of L × W and W × T had significant effects on all three categories of biomass while the interaction S × W showed significant effects on aboveground biomass and total biomass (Table 3). Values for aboveground biomass, belowground biomass, and total biomass were significantly higher for the buried treatment than for the other two but biomass results did not differ significantly between the surface and “between” treatments (Table 5). Significantly more aboveground biomass was produced when seedlings were exposed to the higher water supply but those values were significantly lower in pots where plastic litter was used instead of grass litter (Table 5).
Discussion
In our field and glasshouse experiments, litter had both positive and negative effects on seedling establishment for L. chinensis. This was in accordance with observations from previous studies (Xiong and Nilsson 1999; Jensen and Meyer 2001; Eckstein and Donath 2005; Rotundo and Aguiar 2005). The presence of litter in the field may have negatively influenced seedling emergence because it reduced the level of ambient photon flux density that was available to germinating seeds. Bosy and Reader (1995) have shown that litter also acts as a physical barrier to seedling establishment. Moreover, litter can have an indirect impact because of changes in abiotic conditions, such as light quantity and quality, or, as demonstrated by Fowler (1986) and Chambers (2000), because it can directly reduce the probability for good soil–seed contact. Our findings suggested that litter also had a positive effect on the survival of L. chinensis seedlings in the field because it helped maintain a sufficient level of soil moisture in that semiarid environment. This proposition agreed with that of Evans and Young (1970) and Carson and Peterson (1990), who showed that the presence of litter increases the likelihood of seedling establishment by enhancing the amount of water that is available. In fact, soil moisture improves seed germination for most grassland species (e.g., Baskin and Baskin 2001).
By contrast, the thickness of the litter layer had an insignificant influence on the establishment of B. inermis seedlings in the field and in the glasshouse. This indicated that, even when growing in the same environment, responses to litter quantity can differ among species. Furthermore, many research-related factors can have a role when investigating the impact of litter on plant development, including the type of existing vegetation; study method; length of the experimental period; latitude of the plots; habitat; type, quantity, and mixture of litter; and what target species is being evaluated (Xiong and Nilsson 1999; Quested and Eriksson 2006; Ruprecht et al. 2010; Loydi et al. 2013).
Results from our glasshouse experiments indicated that litter had only a positive effect on the survival of L. chinensis seedlings but was neutral with regard to seedling emergence, biomass production, and heights for both species. These findings concurred with those of Loydi et al. (2013) who reviewed 49 research studies, primarily in Europe and America, and reported that the presence of litter has an overall neutral effect on seedling emergence and survival.
Seed position had a significant effect on seedling emergence and growth performance (height and biomass), but did not influence survival rates. Those significant effects might have arisen because of how the existence of litter can modulate seed position. For example, litter that accumulates on top of seeds and seedlings will exert diametrically opposite effects from those seen in situations such as abandoned grasslands, where seeds will fall on top of the existing litter layer. There, litter can act as a mechanical barrier that prevents seeds from coming into contact with the moist soil, thereby reducing their potential for germination and causing desiccation for any seedlings that do develop within that litter (cf., Facelli and Pickett 1991; Donath and Eckstein 2010). We noted that seeds buried in the soil were associated with dramatically increased percentages of seedling emergence and growth performance both in the field and the glasshouse when compared with the other placements, especially the surface treatment. In our other field reseeding studies, it also showed that harrowing had significantly increased emergence of these two species in contrast to broadcasting only (Liu et al. 2008, 2015). Thus, one might suggest employing practices such as harrowing or an increase in the density of sheep-grazing during late Autumn or early Spring as a means for enhancing the likelihood of seed making adequate contact with the soil. We also found that seeds positioned within the litter layer (but not touching the soil) were linked with reduced seedling emergence and growth. This was in accord with previous reports that positioning of the seeds relative to the soil at the time of germination can ultimately affect seedling emergence (Fowler 1986; Facelli and Pickett 1991; Chambers 2000; Rotundo and Aguiar 2005).
Our glasshouse experiments presented no clear evidence for biological/chemical effects on seedling development by our experimental components. In fact, cumulative rates of emergence differed by only 7 % for L. chinensis and by 13 % for B. inermis in our comparison between plastic and natural grass litter. This has also been demonstrated by Xiong and Nilsson (1999), who stated that the short-term effects of litter tend to be physical rather than biological or chemical. In our examination, the use of plastic litter was associated with higher emergence percentages, growth rates, and survival because, when the same volumes were applied, the plastic layer was thicker than the grass litter, thus improving the amount of water available to both seeds and seedlings. However, there was no significant difference between natural grass litter and plastic litter when measuring total biomass, both for L. chinensis and B. inermis. This further indicated that the short-term effects of litter tend to be physical rather than biological or chemical.
In the glasshouse, a more abundant supply of water had a positive effect on seedling heights and biomass production for both species, but that influence was positive for seedling emergence only with B. inermis. This suggested that water supply is a critical component when determining the success of establishment by seedlings of both species during their first year of development.
In summary, our results provide evidence that litter can have both positive and negative effects on seedling emergence and establishment for L. chinensis and B. inermis. In the short term, the influence of the litter layer is primarily physical for these two semiarid species. Seed position appears to be the most important predictor of seedling emergence and establishment due to the indirect role that litter has as a physical barrier that prevents adequate seed contact with the surrounding soil. Finally, the establishment of seedlings of both species depends upon having a reliable supply of water. However, longer-term experiments are necessary if we are to identify more specifically how the presence of litter influences the establishment of seedlings in semiarid grasslands.
References
Baskin CC, Baskin JM (2001) Seeds, ecology, biogeography and evolution of dormancy and germination. Academic Press, San Diego, p 15
Bissels S, Donath TW, Hölzel N, Otte A (2006) Effects of different mowing regimes on seedling recruitment in alluvial grasslands. Basic Appl Ecol 7:433–442
Bosy JL, Reader RJ (1995) Mechanisms underlying the suppression of forb seedling emergence by grass (Poa pratensis) litter. Funct Ecol 9:635–639
Carson WP, Peterson CJ (1990) The role of litter in an old-field community: impact of litter quantity in different seasons on plant species richness and abundance. Oecologia 85:8–13
Chambers JC (2000) Seed movements and seedling fates in disturbed sagebrush steppe ecosystems: implications for restoration. Ecol Appl 10:1400–1413
Donath TW, Eckstein RL (2010) Effects of bryophytes and grass litter on seedling emergence vary by vertical seed position and seed size. Plant Ecol 207:257–268
Eckstein RL, Donath TW (2005) Interactions between litter and water availability affect seedling emergence in four familial pairs of floodplain species. J Ecol 93:807–816
Eckstein RL, Ruch D, Otte A, Donath TW (2012) Invasibility of a nutrient-poor pasture through resident and non-resident herbs is controlled by litter, gap size and propagule pressure. PLoS ONE 7:e41887
Evans RA, Young JA (1970) Plant litter and establishment of alien weed species in rangeland communities. Weed Sci 19:697–702
Facelli JM, Pickett STA (1991) Plant litter: its dynamics and effects on plant community structure. Bot Rev 57:1–32
Fowler NL (1986) Microsite requirements for germination and establishment of three grass species. Am Midl Nat 115:131–145
Gao JL (2004) Research report on improving, regenerating and managing techniques of Xiashitan grassland. Pratacult Sci 21:6–8 (in Chinese, English summary)
Hovstad KA, Ohlson M (2008) Physical and chemical effects of litter on plant establishment in semi-natural grasslands. Plant Ecol 196:251–260
Jensen K, Gutekunst K (2003) Effects of litter on establishment of grassland plant species: the role of seed size and successional status. Basic Appl Ecol 4:579–587
Jensen K, Meyer C (2001) Effects of light competition and litter on the performance of Viola palustris and on species composition and diversity of an abandoned fen meadow. Plant Ecol 155:169–181
Jia SX (1987) Chinese grassland and forage plants records, vol 1. China Agricultural Press, Beijing
Liu GX, Mao PS, Huang SQ, Sun YC, Han JG (2008) Effects of soil disturbance, seed rate, nitrogen fertilizer and subsequent cutting treatment on establishment of Bromus inermis seedlings on degraded steppe grassland in China. Grass Forage Sci 63:331–338
Liu GX, He F, Wan LQ, Li XL (2015) Management regime and seeding rate modify seedling establishment of Leymus chinensis. Rangel Ecol Manag 68(2):204–210
Loydi A, Eckstein RL, Otte A, Donath TW (2013) Effects of litter on seedling establishment in natural and semi-natural grasslands: a meta-analysis. J Ecol 101:454–464
Miglécz T, Tóthmérész B, Valkó O, Kelemen A, Török P (2013) Effects of litter on seedling establishment: an indoor experiment with short-lived Brassicaceae species. Plant Ecol 214:189–193
Quested H, Eriksson O (2006) Litter species composition influences the performance of seedlings of grassland herbs. Funct Ecol 20:522–532
Rotundo JL, Aguiar MR (2005) Litter effects on plant regeneration in arid lands: a complex balance between seed retention, seed longevity and soil–seed contact. J Ecol 93:829–838
Ruprecht E, Donath TW, Otte A, Eckstein RL (2008) Chemical effects of a dominant grass on seed germination of four familial pairs of dry grassland species. Seed Sci Res 18:239–248
Ruprecht E, Józsa J, Ölvedi TB, Simon J (2010) Differential effects of several “litter” types on the germination of dry grassland species. J Veg Sci 21:1069–1081
Wang RZ, Ripley EA (1997) Effects of grazing on a Leymus chinensis grassland on the Songnen plain of northeastern China. J Arid Environ 36:307–318
Wang QJ, Shi HL, Jing ZC, Wang CT, Wang FG (2004) Recovery and benefit analysis of ecology on degraded natural grassland of the source region of Yangtze and Yellow rivers. Pratacult Sci 21:37–41 (in Chinese, English summary)
Wellstein C (2012) Seed–litter–position drives seedling establishment in grassland species under recurrent drought. Plant Biol 14(6):1006–1010
Xiao XM, Wang YF, Jiang S, Ojima DS, Bonhan CD (1995) Interannual variation in the climate and above—ground biomass of Leymus chinensis steppe and Stipa grandis steppe in the Xilin river basin, Inner Mongolia, China. J Arid Environ 31:283–299
Xiong SJ, Nilsson C (1999) The effects of plant litter on vegetation: a meta-analysis. J Ecol 87:984–994
Zhao MQ, Sun JL (2004) Degraded situation and rehabilitation methods of Jiangjiadian grassland in Jilin province. Pratacult Sci 21:46–48 (in Chinese, English summary)
Acknowledgments
We are grateful for support from the Special Fund for Agro-Scientific Research in the Public Interest (201103005), the National Natural Science Foundation of China (Grant No. 31072069 and 31302015), China Forage and Grass Research System (CARS-35-12), and Cutting-edge and Characteristic Disciplines of Biology (Botany). We thank Priscilla Licht for correcting the English.
Author information
Authors and Affiliations
Corresponding author
Additional information
Communicated by Lori Biederman.
Rights and permissions
About this article

Cite this article
Liu, G., Wan, L., He, F. et al. Effects of litter, seed position, and water availability on establishment of seedlings for two semiarid grass species. Plant Ecol 217, 277–287 (2016). https://doi.org/10.1007/s11258-016-0569-x
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11258-016-0569-x