
Abstract
Understanding how competition from invasive species and soil conditions individually and interactively affect native performance will increase knowledge of invasion dynamics and can be used to improve the success of restoration plans. This study, conducted in Reno NV, USA, uses a two-phase plant–soil feedback experiment coupled with a target-neighbor competition design to examine the individual and interactive effects of both soil conditions and invasive neighbors on native performance. Study species include invasive species (Bromus tectorum and Agropyron cristatum) and native species (Elymus elymoides and Pseudoroegneria spicata). Results indicate that both plant performance and competitive interactions were influenced by species-specific soil conditioning. Specifically, invasive B. tectorum generated a larger competitive effect on natives than invasive A. cristatum; however, only A. cristatum conditioned soil in a manner that increased competitive effects of conspecifics on natives. Native P. spicata was relatively unaffected by soil conditioning and conversely, E. elymoides was strongly affected by soil conditioning. Few previous studies have examined soil conditioning and the interaction of soil conditioning and neighbor effects that both are potentially important mechanisms in structuring plant communities and influencing plant invasion.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Restoration of native plants into areas currently occupied by invasive plants is frequently attempted to control the negative impact of plant invasion on native species diversity (Grice 2009) albeit often with limited success (Eviner and Hawkes 2008; Jordan et al. 2008; Davies and Sheley 2011). Both aboveground effects (e.g., competition, Humphrey and Schupp 2004; Fansler and Mangold 2011) and belowground effects (e.g., plant–soil feedbacks, Jordan et al. 2008; Grman and Suding 2010) of invasive species limit the success of native species restoration. The competitive abilities of invasive species (Elton 1958; Vila and Weiner 2004; Blank 2010) and the impact of this competition during restoration is fairly well established (Eliason and Allen 1997; Cox and Allen 2011; Wainwright et al. 2012). However, the importance of plant–soil feedbacks on the success of native species restoration and on the competitive relationships between invaders and natives is less understood.
Plant–soil feedbacks (PSF) occur when plants alter soil conditions, such as soil biological community composition (Van der Putten et al. 1993) and soil nutrient availability (Dijkstra et al. 2006; Meisner et al. 2011; Perkins et al. 2011a) and thus influence subsequent plant performance (Bever 2003; Kulmatiski et al. 2008; Perkins and Nowak 2012). Research and meta-analysis indicate that, in general, most plants create PSFs that decrease subsequent conspecific performance (Kulmatiski et al. 2008). However, an increasing number of studies suggest that invasive species tend to either be unaffected by PSF or create PSFs that increase subsequent conspecific performance (van Grunsven et al. 2007; Perkins and Nowak 2012) and decrease subsequent heterospecific performance (Jordan et al. 2008). Previous research on how alteration of soil conditions affects competitive relationships is limited (Wardle et al. 2004; Bever et al. 2010; Kulmatiski et al. 2011). In addition, “aboveground implications of belowground processes” are not easy to predict (Wardle et al. 2004), may exert large effects on plant–plant interactions (Bever et al. 2010), and are needed to better understand invasion and restoration dynamics.
This study investigates (1) how performance of two native grass species is influenced by PSFs and by competition from invasive plants and (2) if PSF changes these competitive relationships. Specifically, this study questions how performance of natives Elymus elymoides and Pseudoroegneria spicata are affected by competing neighbors of invasive Bromus tectorum and Agropyron cristatum in soils previously occupied by either the native or the invaders (Fig. 1). Species were chosen so as to include two common native grasses and two common invasive grasses found within the Great Basin of the western US. Although A. cristatum has been intentionally seeded within the Great Basin during reclamation efforts, it meets the definition of an invasive species (non-native, able to expand populations over a considerable distance, and produce negative effects on native ecosystems, Richardson et al. 2000; Fansler and Mangold 2011). Our previous work on PSF dynamics of the study species (Perkins and Nowak 2012) indicates that both natives E. elymoides and P. spicata influence soil in a manner that benefits heterospecifics more than conspecifics. Invasive A. cristatum influences soil in a manner that benefits conspecifics more than heterospecifics. Finally, invasive B. tectorum does not alter soil in a manner that significantly affects subsequent plant performance. These invasive species (B. tectorum and A. cristatum) do generate larger competitive effects than the native species in this study (E. elymoides and P. spicata), especially in seedling stages (Humphrey and Schupp 2004; Blank 2010; Gunnell et al. 2010). Competition from invasives has been observed to significantly decrease both E. elymoides and P. spicata seedling biomass in a glasshouse setting (Blank 2010; Gunnell et al. 2010) and in the field (Humphrey and Schupp 2004). Therefore, we hypothesized that natives would be negatively affected individually by invader presence and by conspecific PSF. We also hypothesized that the invader A. cristatum would have a larger competitive effect in soils previously occupied by conspecifics and that both invaders would have larger competitive effects on natives in soil previously occupied by the natives than in previously unoccupied soil. Because of limited knowledge regarding effects of soil conditions on competitive relationships, we generated our hypotheses on the straightforward reasoning that (1) if a plant has increased performance in conspecific-conditioned soil, it should also generate larger competitive effects on neighbors in that soil; and (2) if a species has decreased performance in conspecific-conditioned soil, it should also be more vulnerable to competition from neighbors in that soil.

Diagram of the experimental design. One individual plant of either target species (native grass species, E. elymoides and P. spicata) were grown with one individual of either neighbor species (invasive grass species, B. tectorum or A. cristatum), a conspecific, or with no neighbor in soil conditioned by either the target, the neighbor, or in unconditioned soil
Materials and methods
To examine effects of competition and plant–soil feedback on native species performance, a glasshouse experiment in two phases was conducted. During the soil conditioning phase, a first generation of plants were grown to condition (induce species-specific changes in) soil. The soil conditioning treatments include conditioning by each of the natives (E. elymoides and P. spicata), each of the neighbors (B. tectorum and A. cristatum), and an unconditioned control. For the unconditioned soil treatment, pots were filled with soil, but left unplanted and otherwise treated the same (e.g., received the same watering regime) as the planted pots. The second phase was the bioassay phase, wherein a target-neighbor design was established to determine competitive relationships in each of the soil conditioning treatments (Fig. 1). The experimental design for the bioassay phase included native species (E. elymoides and P. spicata) as targets growing in (1) unconditioned soil with either no neighbor, a conspecific neighbor, a B. tectorum neighbor, or an A. cristatum neighbor; (2) conspecific-conditioned soil with either no neighbor, a conspecific neighbor, a B. tectorum neighbor, or an A. cristatum neighbor; (3) B. tectorum-conditioned soil with either no neighbor, a conspecific neighbor, or a B. tectorum neighbor; and (4) A. cristatum-conditioned soil with either no neighbor, a conspecific neighbor, or an A. cristatum neighbor. Because the effect of the two invasive neighbors on each other was not of interest, the combination of A. cristatum as a neighbor in B. tectorum-conditioned soil and B. tectorum as a neighbor in A. cristatum-conditioned soil were not included (Fig. 1). The entire design was replicated 10 times.
Soil used for this project was native soil collected from Bedell Flats, a natural area approximately 40-km northeast of Reno, NV, USA. The soil is a Durinodic Xeric Haplargid (USDA, NRCS) and is a moderately fertile sandy loam (Perkins et al. 2011a). Major species in the area include sparse Artemisia tridentata and Chrysothamnus nauseosus although soil was collected from unvegetated areas. The top 20 cm of soil was collected, homogenized, and potted into RLC 3 containers (49 ml volume, Stuewe & Sons, Corvallis OR USA) in a glasshouse in Reno, NV USA. The glasshouse was maintained to reflect typical growing season conditions, i.e., between 7 and 24 °C with ambient light. Distilled water was used to maintain pots near field capacity without allowing any leaching or overflow. Pots were randomly assigned to soil conditioning treatment and target-neighbor species, and pots were periodically rearranged to compensate for any environmental variation in the glasshouse.
In the first phase (the soil conditioning phase), several seeds were planted in each pot, but only the first emergent was allowed to grow. After 80 days of growth, aboveground biomass of the conditioning plants was removed. To transition to the second phase (target-neighbor bioassay phase), all soil conditioned by an individual species within one replicate was combined, homogenized, and then repotted. Seeds for the bioassay phase were planted within 1 day of the end of the soil conditioning phase. Several seeds of each target species and neighbor species were planted, and the first emergent of each species was allowed to grow. Date of emergence for each species within each pot was recorded. The growing season for the bioassay phase lasted for 80 days, after which all aboveground biomass was collected, dried at 60 °C for 24 h, and weighed.
A relative interaction index (R ii, Armas et al. 2004) was adapted to calculate the net effect of soil conditioning (PSF response) and then the net balance of the interaction between plants grown alone and plants grown with neighbors within each soil conditioning treatment (Competition effect). The formulae for PSF response and for Competition effect are
where B is the aboveground biomass of each species (growing without a neighbor) under each soil conditioning treatment (B c) and the mean biomass of that species under other soil conditioning treatments (B a) within each replicate.
where B is the aboveground biomass of a single plant growing with a neighbor (B w) and alone (B o). Competition effect was calculated separately for each soil conditioning treatment, i.e., the biomass of a target species growing with a neighbor in a given soil conditioning treatment and the biomass of a target growing alone in that same soil conditioning treatment were used in the calculations.
A positive value indicates that a plant produced more biomass due to soil conditioning, or with a neighbor; a negative value indicates that a plant produced less biomass when grown in conditioned soil or without a neighbor.
Data were analyzed with PASW Statistics 18 (PASW for Windows, Rel. 18.0.0. 2009. Chicago: SPSS Inc.). Box-plots were used to examine for outliers, and Shapiro–Wilk was used to evaluate normality (no data were dropped and no transformations were necessary). Multivariate analysis was used to examine the effect of soil conditioning treatment, neighbor species, and the interaction of soil conditioning and neighbor species on target time to emergence, and examine biomass separately for each species. To minimize the potential for Type I error, MANOVA for overall effects and Tukey HSD comparisons for pairwise differences among soil conditioning treatments and neighbor species were used. In addition, separately for each species, univariate analyses (ANOVA) were conducted to examine the effect of soil conditioning treatment on PSF response and the effect of soil conditioning treatment, neighbor species, and the interaction of soil conditioning and neighbor species on competitive effect.
Results
Plant emergence was not affected by soil conditioning treatment or by neighbors (Table 1). Average time to E. elymoides emergence was 10.1 days (SE 0.2) and was not significantly affected by soil conditioning treatment, neighbor species (p = 0.11), or soil conditioning by neighbor interaction. Average time to P. spicata emergence was 8.7 days (SE 0.1) and was not significantly affected by soil conditioning species, neighbor species, or soil conditioning by neighbor interaction.
Soil conditioning had significant effects on both E. elymoides and P. spicata biomass (Table 1). The biomass produced by both E. elymoides and P. spicata was significantly different among soil conditioning treatments (Fig. 2). E. elymoides produced the most biomass in unconditioned soil, less biomass in A. cristatum- and B. tectorum-conditioned soils (22 and 35 % reduction, respectively), and the least biomass (57 % reduction) in its conspecific-conditioned soil. Pseudoroegneria spicata also produced the most biomass in unconditioned soil; however, the reduction of P. spicata biomass was only significant in conspecific-conditioned soil and B. tectorum-conditioned soil (32 and 26 % reduction respectively). Thus, the ability of both native species to produce biomass was significantly affected by soil conditioning treatments.

Biomass produced by native grasses E. elymoides (a) and P. spicata (b) when grown alone and with neighbors in soils either previously unoccupied, or occupied by a conspecific or an invader (either B. tectorum or A. cristatum). Bars with different letters indicate a significant difference among neighbor species within one soil conditioning treatment. Bars with different numbers indicate a significant difference in the performance of the native grown with no neighbor among soil conditioning treatments. Significance was determined at a p < 0.05. Error bars are one SE
Neighbors had significant effects on both E. elymoides and P. spicata biomass (Table 1). The effects of neighbors on target performance was examined using biomass and competition effect values produced by each target species in the unconditioned soil (soil conditioning treatment “none” in Figs. 2, 4). Patterns of competitive effects of neighbors on target biomass was common to both target species: (1) the presence of any neighbor significantly reduced target biomass; (2) the reduction in target biomass was not significantly different between conspecific and A. cristatum neighbors (46 and 52 % reduction of E. elymoides and 56 and 50 % reduction of P. spicata respectively); and (3) B. tectorum as a neighbor reduced target biomass the most (70 % reduction of E. elymoides and 71 % reduction of P. spicata). In general, B. tectorum produced larger competitive effects than A. cristatum.
Plant biomass in soils without neighboring species was used to test PSF response (Fig. 3). The stronger response of E. elymoides biomass production to soil conditioning treatments without neighbors resulted in a significant PSF response among soil conditioning treatments. Elymus elymoides experienced a significant negative PSF response in conspecific soil (Fig. 3). However, the response of P. spicata biomass production to soil conditioning treatments without neighbors was not enough to generate a significant PSF response (Fig. 3).

Index showing the effect of soil conditioning (PSF) on native plant (E. elymoides and P. spicata) performance grown without neighbors. A positive value indicates that a plant produced more biomass in a given soil conditioning treatment than its mean biomass in all the treatments. Bars with different letters indicate a significant difference among soil conditioning treatments. Significance was determined at a p < 0.05. Error bars are one SE
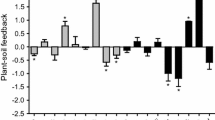
The interaction of soil conditions and neighbors were examined using competition effect values (Fig. 4). Neither native species was affected by neighbors more in their own conditioned soil than in the other soil conditioning treatments. Native E. elymoides was affected less by neighbors in its own conditioned soil than in unconditioned soil (represented by smaller competition effect values in the conspecific soil conditioning treatment than in other treatments, Fig. 2e), and native P. spicata tended to affected similarly by neighbors in its own conditioned soil compared to the unconditioned soil (Fig. 4). The effect of B. tectorum on P. spicata was consistently large and did not change among soil conditioning treatments (Fig. 4a). However, for E. elymoides, the competitive effect of B. tectorum was significantly different among soil conditioning treatments. Bromus tectorum produced the largest competitive effect in the unconditioned soil and the least in E. elymoides conspecific-conditioned soil (Fig. 4a). The competitive effect of A. cristatum on native E. elymoides (Fig. 4b) was significantly larger in A. cristatum-conditioned soil than in E. elymoides conspecific-conditioned soil. The competitive effect of A. cristatum on native P. spicata was not different among soil conditioning treatments (Fig. 2b).

Index showing the effect of neighbors on native plant performance among the soil conditioning treatments. A negative value indicates that native plant biomass was reduced by neighbor’s presence. Bars with different letters indicate a significant difference among soil conditioning treatments. Significance was determined at a p < 0.05. Error bars are one SE
Discussion
Performance of native species was affected by both aboveground (competition) and belowground (PSF) effects of invaders. More importantly, PSF changed competitive relationships between natives and invaders. This evidence that soil conditioning can influence competitive relationships has implications both for understanding plant population dynamics, plant community interactions, invasion dynamics, and ecological restoration (Wardle et al. 2004; Bever et al. 2010; van de Voorde et al. 2011). The effect of soil conditioning on competitive relationships can influence invasion dynamics as either an ability of the invader or as a characteristic of the potentially invaded system (Perkins et al. 2011b). The ability of some species, such as A. cristatum, to create PSFs that increase conspecific performance and competitive ability may increase their invasive potential (Klironomos 2002; Kulmatiski et al. 2008). Further, sites occupied by natives, such as E. elymoides, that condition soils in a manner that decreases subsequent conspecific performance or increase heterospecific performance (e.g., P. spicata, Perkins and Nowak 2012) might be especially vulnerable to invasion. Restoration of sites occupied by invaders that benefit from their own soil conditioning (e.g., A. cristatum) may require more attention to belowground conditions (Grman and Suding 2010) than sites occupied by invaders that do not benefit from soil conditioning (e.g., B. tectorum).
Interestingly, the invaders showed two different interactive relationships between competitive ability and plant–soil feedbacks. Invader B. tectorum had consistently high competitive ability in all but one of the soil conditioning treatments (Figs. 2, 4). Bromus tectorum did not produce significant PSFs (Fig. 3) and its competitive ability did not change substantially among most soil conditioning treatments. Conversely, invader A. cristatum only produced higher competitive effects than the native’s conspecific neighbors in its conspecific-conditioned soil (Figs. 2, 4). Thus, the invasive potential (Perkins et al. 2011b) of B. tectorum may be derived from competitive ability and not soil conditioning and the invasive potential of A. cristatum may be derived from soil conditioning more than competitive ability. Like A. cristatum, other invasive plants are also thought to be successful invaders due to the changes they create in their soil environment more than their competitive ability. For example, Typha × glauca invasion in coastal marshes has been attributed to its ability to alter soil conditions in a manner that decreases heterospecific performance and increases conspecific performance more than its competitive ability (Farrer and Goldberg 2009).
The plant–soil feedback generated by invader A. cristatum is worthy of note. Soil conditioned by A. cristatum resulted in increased E. elymoides performance indicated by a positive PSF response value (Fig. 3). This result is consistent with previous work that indicates that A. cristatum may produce plant–soil feedbacks that increase subsequent plant growth (Jordan et al. 2008; Perkins and Nowak 2012). However, this benefit to heterospecifics may only manifest when they are growing alone. Agropyron cristatum has been observed to produce a conspecific-positive feedback type, meaning all subsequent plant growth is promoted, but conspecifics have a larger response than heterospecifics (Perkins and Nowak 2012). Although soil conditioned by A. cristatum might benefit E. elymoides growing alone, the benefit to subsequent A. cristatum performance (increased biomass production, Perkins and Nowak 2012) and competitive ability (Fig. 4) outweigh any benefit to a native growing alongside the invader.
These results have several implications for restoration and provide an example that “one way fits all” restoration plans are not sufficient (Kardol and Wardle 2010). Native E. elymoides performed especially poorly in conspecific-conditioned soil; therefore, seeding E. elymoides into soils that previously had E. elymoides present might be less successful than into other soils. Native P. spicata was impacted by neighbor presence, but not by PSF; therefore, restoration of P. spicata should focus on neighbor presence more than soil conditioning. Based on the result that invader B. tectorum induced a large competitive effect on native performance but did not have more of an effect in its own conspecific-conditioned soil, a restoration plan involving B. tectorum may need to address the invader’s presence more than soil conditions. Conversely, invader A. cristatum did not produced competitive effects that were significantly different from the competitive effect produced by the native’s conspecific neighbor except in its own conditioned soil; therefore, a restoration plan involving A. cristatum may need to address soil conditions as well as invader presence. Restoration strategies that address soil conditioning may include planting native species that are less affected by soil conditioning (van de Voorde et al. 2011) or amending the soil to remove the conditioning effects (Grman and Suding 2010). Also, reclamation plans that involve intentionally introducing A. cristatum for site stabilization should be cautiously considered as native species at these sites may be negatively impacted by soil conditioning.
How alteration of soil conditions affects competitive relationships in a natural setting may only marginally be anticipated by a target-neighbor competition in soils conditioned for one generation in a glasshouse setting. Studies on PSFs in a glasshouse usually find stronger effects than field studies (Kulmatiski et al. 2008), and PSFs may change with increasing plant density and neighbor identity (Blank 2010). Therefore, this project can be used to generate hypotheses (Freckleton and Watkinson 2000) for further testing. Appropriate hypotheses might address how soil conditioning changes with increasing conspecific and heterospecific neighbors, how PSFs compound and change over time, and how PSFs manifest under field (compared to controlled glasshouse) temperature and water availability conditions. This study only examined the target-neighbor relationship for 80 days (emergence and seedling life stages) and relationships that reduce target performance at the seedling stage may change to neutral or positive at later life stages (Leger and Espeland 2010; Farrer and Goldberg 2011). However, our results agree with other target-neighbor competition studies that include these species (Blank 2010; Gunnell et al. 2010). Blank (2010) also found P. spicata was more negatively affected by B. tectorum neighbors than conspecific neighbors, and Gunnell et al. (2010) also found E. elymoides and A. cristatum induce similar competitive effects. The negative effect of B. tectorum on E. elymoides has been observed to continue through 2 years in the field (Humphrey and Schupp 2004).
In summary, this project provides evidence that prior vegetation has potential to influence subsequent native plant performance and competitive relationships with neighbors. Specifically, both natives were negatively impacted by the presence of neighbors, and for E. elymoides, these competitive relationships changed with soil conditioning treatments. Invader A. cristatum generated a PSF that increased its competitive ability and B. tectorum created a consistently large competitive in most soil conditioning treatments.
References
Armas C, Ordiales R, Pugnaire FI (2004) Measuring plant interactions: a new comparative index. Ecology 85:2682–2686
Bever JD (2003) Soil community feedback and the coexistence of competitors: conceptual frameworks and empirical tests. New Phytol 157:465–473
Bever JD, Dickie IA, Facelli E, Facelli JM, Klironomos J, Moora M, Rillig MC, Stock WD, Tibbett M, Zobel M (2010) Rooting theories of plant community ecology in microbial interactions. Trends Ecol Evol 25:468–478
Blank RR (2010) Intraspecific and interspecific pair-wise seedling competition between exotic annual grasses and native perennials: plant–soil relationships. Plant Soil 326:331–343
Cox RD, Allen EB (2011) The roles of exotic grasses and forbs when restoring native species to highly invaded southern California annual grassland. Plant Ecol 212:1699–1707
Davies KW, Sheley RL (2011) Promoting native vegetation and diversity in exotic annual grass infestations. Restor Ecol 19:159–165
Dijkstra FA, Hobbie SE, Reich PB (2006) Soil processes affected by sixteen grassland species grown under different environmental conditions. Soil Sci Soc Am J 70:770–777
Eliason SA, Allen EB (1997) Exotic grass competition in suppressing native shrubland re-establishment. Restor Ecol 5:245–255
Elton C (1958) The ecology of invasions by animals and plants. University of Chicago Press, USA
Eviner VT, Hawkes CV (2008) Embracing variability in the application of plant–soil interactions to the restoration of communities and ecosystems. Restor Ecol 16:713–729
Fansler VA, Mangold JM ( 2011) Restoring native plants to crested wheatgrass stands. Restor Ecol 19:16–23
Farrer EC, Goldberg DE (2009) Litter drives ecosystem and plant community changes in cattail invasion. Ecol Appl 19:398–412
Farrer EC, Goldberg DE (2011) Patterns and mechanisms of conspecific and heterospecific interactions in a dry perennial grassland. J Ecol 99:265–276
Freckleton RP, Watkinson AR (2000) On detecting and measuring competition in spatially structured plant communities. Ecol Lett 3:423–432
Grice T (2009) Principles of containment and control of invasive species. Invasive species management: a handbook of principles and techniques. Oxford University Press Inc, New York, pp 61–76
Grman E, Suding KN (2010) Within-year soil legacies contribute to strong priority effects of exotics on native California grassland communities. Restor Ecol 18:664–670
Gunnell KL, Monaco TA, Call CA, Ransom CV (2010) Seedling interference and niche differentiation between crested wheatgrass and contrasting native great basin species. Rangel Ecol Manag 63:443–449
Humphrey LD, Schupp EW (2004) Competition as a barrier to establishment of a native perennial grass (Elymus elymoides) in alien annual grass (Bromus tectorum) communities. J Arid Environ 58:405–422
Jordan NR, Larson DL, Huerd SC (2008) Soil modification by invasive plants: effects on native and invasive species of mixed-grass prairies. Biol Invasions 10:177–190
Kardol P, Wardle DA (2010) How understanding aboveground-belowground linkages can assist restoration ecology. Trends Ecol Evol 25:670–679
Klironomos JN (2002) Feedback with soil biota contributes to plant rarity and invasiveness in communities. Nature 417:67–70
Kulmatiski A, Beard KH, Stevens JR, Cobbold SM (2008) Plant–soil feedbacks: a meta-analytical review. Ecol Lett 11:980–992
Kulmatiski A, Heavilin J, Beard KH (2011) Testing predictions of a three-species plant–soil feedback model. J Ecol 99:542–550
Leger EA, Espeland EK (2010) The shifting balance of facilitation and competition affects the outcome of intra- and interspecific interactions over the life history of California grassland annuals. Plant Ecol 208:333–345
Meisner A, de Boer W, Verhoeven KJF, Boschker HTS, van der Putten WH (2011) Comparison of nutrient acquisition in exotic plant species and congeneric natives. J Ecol 99:1308–1315
Perkins LB, Nowak RS (2012) Native and non-native grasses generate common types of plant–soil feedbacks by altering soil nutrients and microbial communities. Oikos. doi:10.1111/j.1600-0706.2012.20592.x
Perkins LB, Johnson DW, Nowak RS (2011a) Plant-induced changes in soil nutrient dynamics by native and invasive grass species. Plant Soil 345:365–374
Perkins LB, Leger EA, Nowak RS (2011b) Invasion triangle: an organizational framework for species invasion. Ecol Evol 4:610–625
Richardson DM, Pysek P, Rejmanek M, Barbour MG, Panetta FD, West CJ (2000) Naturalization and invasion of alien plants: concepts and definitions. Divers Distrib 6:93–107
van de Voorde TFJ, van der Putten WH, Bezemer TM (2011) Intra- and interspecific plant–soil interactions, soil legacies and priority effects during old-field succession. J Ecol 99:945–953
van der Putten WH, Vandijk C, Peters BAM (1993) Plant-specific soil-borne diseases contribute to succession in foredune vegetation. Nature 362:53–56
van Grunsven RHA, van der Putten WH, Bezemer TM, Tamis WLM, Berendse F, Veenendaal EM (2007) Reduced plant–soil feedback of plant species expanding their range as compared to natives. J Ecol 95:1050–1057
Vila M, Weiner J (2004) Are invasive plant species better competitors than native plant species? Evidence from pair-wise experiments. Oikos 105:229–238
Wainwright CE, Wolkovich EM, Cleland EE (2012) Seasonal priority effects: implications for invasion and restoration in a semi-arid system. J Appl Ecol 49:234–241
Wardle DA, Bardgett RD, Klironomos JN, Setala H, van der Putten WH, Wall DH (2004) Ecological linkages between aboveground and belowground biota. Science 304:1629–1633
Acknowledgments
This research was supported by the Nevada Agricultural Experiment Station. This manuscript was improved greatly by comments, advice, and feedback provided by anonymous reviewers and Scot Ferguson.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Perkins, L.B., Nowak, R.S. Soil conditioning and plant–soil feedbacks affect competitive relationships between native and invasive grasses. Plant Ecol 213, 1337–1344 (2012). https://doi.org/10.1007/s11258-012-0092-7
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11258-012-0092-7