
Abstract
Grazing constitutes a selective pressure on vegetation recruitment through modification of the seed banks. Here we address changes in seed bank density and its life history trait composition in century-old pastures, where contrasting reindeer densities have developed during the last decades. We cover the actual scales used by these wide-roaming herbivores by sampling 70 productive tundra habitats over 7,421 km2 in the reindeer summer pastures of Northern Norway. Results showed that the seed bank density was significantly lowered where reindeer densities had increased in recent decades, whereas the century-long history of grazing probably explains the main seed bank traits typical of grazing tolerant plants. The dominant trait characteristics were small seeds (<0.5 mg), seeds lacking dispersal mechanism and individuals with a graminoid growth form. Finally, differences between regions introduced trait variation independent of grazing history, reinforcing the importance of using scales covering several contexts when designing ecological studies.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Seed bank accumulation is one of the most important strategies for plant survival and propagation in disturbed and variable environments, such as grazed ecosystems (Chambers and Macmahon 1994). It is well documented that the vertebrate grazing changes the seed abundance and species composition of the seed banks (e.g., Chippindale and Milton 1934; O’Connor and Pickett 1992; Jutila 1998; Mayor et al. 2003; Sternberg et al. 2003; Eskelinen and Virtanen 2005). However, studies on seed bank trait composition in relation to grazing are few, in spite of the scientific evidence showing that plant life history traits influence plant tolerance to grazing (Diaz et al. 2001; Sternberg et al. 2003).
Plant life history traits are used to monitor the effects of herbivores on the vegetation (e.g., Wardle et al. 1998; McIntyre et al. 1999; Rusch et al. 2003; Cornelissen et al. 2004; Bråthen et al. 2007b). Because the seed bank is an important vegetation regeneration stage, the study of its life history traits could give information on the direction to which the vegetation is heading in case of disturbance. Studies on ungulate grazing in northern areas have shown that the use of growth forms to describe the vegetation reliably capture plants’ different responses to grazing (White and Trudell 1980; Chapin et al. 1996; Bråthen and Oksanen 2001; Bråthen et al. 2007b). This, in turn is also likely to be manifested as changes in the seed bank. Changes in seed bank structure associated with grazing could also be influenced by processes related to seed and dispersal unit characteristics, because of their critical role in determining species responses to disturbance (Navie and Rogers 1997; Osem et al. 2006). Grazing is known to favor plants with small and round seeds lacking dispersal assisting structures (Eriksson and Eriksson 1997; Navie and Rogers 1997). Thus, in our approach to study the seed bank under grazing conditions, we consider both plant traits (growth form) and seed traits (seed mass and diaspore morphology).
Based on hierarchical foraging theory (Senft et al. 1987) it can be expected that the effect of large herbivores on the seed banks may be highly spatially variable and dependent on type of grazer. However, the effect of grazers on seed banks has previously only been addressed in small extent studies (Jutila 1998; Eskelinen and Virtanen 2005; Olofsson 2006; Osem et al. 2006), which may be highly dependent on specific ecological context (Noda 2004) and which fail to include enough spatial variation in their design (Hobbs 2003). Here we apply a design including several ecological contexts and covering the actual scales used by a wide-roaming large herbivore that covers hundreds of km2 while grazing its pastures.
Reindeer (Rangifer tarandus tarandus) are often the most abundant large-roaming herbivore found in the circumpolar region (Batzli et al. 1980; Klein 1999). Reindeer are known to pick their food non-randomly by selecting plant reproductive parts and some species before others (White and Trudell 1980; Klein 1990; Wada 1999; Bråthen and Oksanen 2001; Cooper and Wookey 2003). As a result, reindeer can change the seed bank density (Cooper 2006) and potentially shape the composition of the plant life history traits present in the seed bank.
Reindeer have been present in coastal northern Norway since the deglaciation of 10,000–15,000 years BP (Skogland 1994). Herds migrate seasonally between inland winter pastures and coastal summer pastures. Since the sixteenth century, reindeer herds in Finnmark have become semi-domesticated and are now entirely herded by the indigenous Sámi people, although the seasonal migration pattern is maintained (Muga 1986). The summer pastures are naturally delineated by the coastal line into distinct geographic areas, and these are separated by fences as to form discrete management districts. These summer districts vary in terms of pasture area and reindeer density. Over the last two to three decades significant density-dependent reductions in calf growth have existed between districts (Fauchald et al. 2004; Tveraa et al. 2007), probably as a result of the deteriorated quality of the grazing grounds where densities have been high in the last two to three decades (Bråthen et al. 2007b). Yet, no study has evaluated at a large scale the reindeer grazing impact on the potential recruitment of the vegetation.
It is hypothesized in this study that reindeer grazing has modified the seed bank density of the pastures where reindeer density has increased over the last decades. Also, it is hypothesized that this grazing constitutes a selective pressure on the composition of life history traits in the seed bank. To test out these hypotheses, we paired neighboring reindeer herding districts with contrasting histories of reindeer densities. The opportunity to implement a quasi-experimental (Shadish et al. 2002) study design, with both a temporal dimension (i.e., different histories of reindeer densities) and a spatial dimension (i.e., contrasting large-scale areas of different histories) is not common. As such, this seed bank study represents a pioneering approach.
Methods
Study area
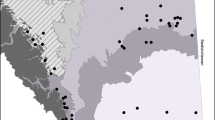
The study was carried out in the coastal alpine tundra of Finnmark County, Northern Norway, between latitudes 77°91′N–30°90′E and 78°54′N–64°28′E (Fig. 1). The study sites were located in the low alpine zone, where snow cover remains until late June and the main vegetation types are grass and herbaceous dicotyledon meadows, Empetrum- and Vaccinium-dominated heaths and Eriophorum and Carex marshes. The main herbivore in these areas are semi-domestic reindeer (Rangifer tarandus tarandus), followed in a less extent by moose (Alces alces), and locally, domestic sheep (Ovis aries). Also widespread are Norwegian lemmings (Lemmus lemmus), ptarmigan (Lagopus lagopus), and gray sided voles (Clethrionomys rufocanus), although high peak spring densities have been lacking during the last decades (Ims and Fuglei 2005). Finnmark County is characterized by climatically steep gradients from west to east as well as from coast to inland (Hanssen-Bauer 1999), caused by the NE Atlantic Current that gradually declines from west to east. The northernmost coast of Finnmark is classified as Arctic (Walker et al. 2005). Average (range in parenthesis) precipitation in the summer months is 65 (52–84), 54 (47–60), and 48 (38–55) mm in the region west, region middle, and region east of Finnmark, respectively. Average (range in parenthesis) temperature is 10.6 (8.5–12.1), 10.1 (7.9–11.6), and 8.7 (6.2–10.5)°C, in the region west, region middle, and region east, respectively (30 year normal, average for June, July, and August; Norwegian Meteorological Institute 2006).

Study design and location of the study. One pair of districts (stable reindeer density district and increased reindeer density district) was selected in three Finnmark regions (region west, region middle, and region east). Each of the six districts were further divided in hierarchical levels, first in 37 landscape areas (20 in increased density districts and 17 in stable density districts), and then in 70 habitats (38 in increased density districts and 32 in stable density districts). Each habitat contained between 1 and 5 gradients down slope (91 in increased density districts and 83 in stable density districts), along where the samples were collected in plots (175 in increased density districts and 205 in stable density districts). One sample was collected per plot
Study design
For this study six large-scale reindeer herding districts, ranging in area from 300 to 4,000 km2 (total area 7,421 km2), were paired according to their history of reindeer density. The two districts forming a pair had maintained constant reindeer numbers from the 1950’s to the 1980’s, after which, one of the districts in the pair increased its density from 100 to 200% (i.e., increased reindeer density district), while the other district maintained its density constant (i.e., stable reindeer density district) (Anonymous 2004) (Fig. 2). Density changes are to a large extent determined by spatio-temporally heterogeneous management practices (Tveraa et al. 2007; Naess et al. 2009). Feces counts confirmed that the focal habitats of the increased reindeer density districts had a higher animal density than habitats from stable density districts. Furthermore, both districts forming a pair had to be neighboring districts in order to reduce climatic and geoedaphic differences sensu Kruckeberg (2002). Under these criteria, three pairs of districts were identified. The pairs were located in three different regions, covering the gradient from the westernmost to the easternmost parts of Finnmark and termed region west, region middle, and region east (Fig. 1).

History of reindeer density (mean number of reindeer per km2 with 95% confidence intervals) for districts in the west, middle, and east region from 1945 to 1980 and from 1980 to 2003. This reindeer density development provides the basis for the contrasting between districts with a history of increased or stable reindeer density districts, respectively
In order to randomly choose representative habitats, each district was divided into nested levels (Fig. 1). The first level was landscape areas of 2 × 2 km2. Landscape areas containing major roads, sea, lakes, glaciers, or more than 50% forests were discarded.
Within each landscape area, suitable habitats were located based on topographic curvature, slope, and altitude above sea level (asl) in order to stratify our sampling to relatively productive habitats in the low-alpine zone. Areas with concave curvature accumulate moisture and nutrients, hence, they were assumed to be relatively more productive. Furthermore, snow beds in concave habitats have been shown to attract reindeer (Edenius et al. 2003) and accumulate seeds (Larsson and Molau 2001). Therefore, concave habitats with a mean slope between 7.5° and 30° (between 5° and 30° in the Region East which is less steep) were chosen. Terrain above 600 m asl in Region West, above 500 m asl in Region Middle Finnmark, and above 300–350 m asl in Region East, was excluded. Altitudinal belts above these heights generally represent the middle to high alpine zone, which contains sparse vegetation. Potential habitats were located selecting pixels with the specified topographic measures from a terrain model with 25 × 25 m resolution using GIS software (ArcGIS 2004). Each habitat was located in the field with geographical coordinates (longitude and latitude) provided by the GIS programme.
Landscape areas that contained the largest number of potential habitats were visited, 20 and 17 landscape areas for increased and stable districts, respectively. Once located in the field, the habitat had to meet certain conditions. First, the vegetation cover had to be higher than 75%, and second, the study site had to be large enough to include at least two gradients (each no less than 10 m long) and should not contain paths, electric posts, marsh, or bog. If a habitat failed to meet any of these conditions, it was discarded and the next one closest to it was visited and inspected for possible analysis. Once the habitat was selected, between two and five gradients were laid out down slope with a 5-m separation between them. Gradient length varied between 10 and 50 m and was limited by the habitat size since sampling was stopped before entering new convex area with heath vegetation. Samples were gathered in plots every tenth meter of each gradient in order for data to be representative of the habitat. Between 1 and 5 plots were sampled per gradient depending on its length. If the plot had more than 50% stones, it was discarded and the plot in the next 2 m was taken instead. If that plot also failed to meet the conditions, the plot 2 m before the original plot was taken. If none of those options was successful, the sample was not collected. The amount of gathered samples per habitat varied between 2 and 14.
The two districts of a pair were sampled simultaneously in order to avoid differences in plant phenology. Data were assembled over 4 weeks, starting in the region west, during July and August 2004. At that time, current year seeds were dispersing. It is therefore possible that the samples contained both transient seeds, i.e., with a life span of less than a year, and persistent seeds, i.e., at least 1-year-old (Thompson et al. 1997). Plant nomenclature follows Lid and Lid (1994).
Sampling
In each plot, a 2.5-cm diameter soil core was collected with a 10-cm long metal cylinder placed down at soil level, below the humus layer, on the bottom right corner of the studied plot. The core depth varied in accordance with the thickness of the organic soil layer of the plot in order for all seeds, recent and old, to be collected. Top layer samples, i.e., moss and litter layer down to the humus layer, were taken by collecting a 15 × 15 cm sample from each studied plot. All samples were stored in paper bags and frozen no later than 8 h after collection. Samples were frozen at −18°C in order to simulate a stratification period. After 2 months they were taken out to thaw in dark chambers at 2°C before the experiment started. A total of 380 soil core samples and 384 top layer samples were collected from the 70 visited habitats.
Soil core samples were crumbled onto Petri dishes (diameter 8.7 cm, depth 1.4 cm) with a filter paper in the bottom to keep moisture. About half of every top layer sample (10 × 10 cm) was put in a transparent box (15 × 10 cm), also with filter paper in the bottom. All samples were randomly placed in two phytotron chambers where optimal germination conditions for most northern alpine plants were simulated: 24 h of daylight split into 12 h at 20°C and 12 h at 15°C. All samples were kept moist by water spraying every third day. Emerged seedlings were transplanted to seedling boxes and kept until identification was possible. After 4 months, all samples were set in refrigerated dark chambers at 2°C in order to simulate a winter period. Soil cores were crumbled again and returned together with the top layer samples to the growth chambers for the germination to continue after 3 weeks of chilling. To further stimulate seed germination, samples were sprayed after the second chilling period once a week, during 2 weeks, with KNO3 (10 mM; Giba et al. 2003). Seedling emergence was monitored during a total of 7 months, until no more seedlings emerged from the samples. Seedlings dying before identification were classified in general groups: dicotyledon, graminoid, ericoid shrub, or “plant.” Some graminoids were cold treated (0.5°C) to initiate flowering, in order to identify them. Measurements of soil pH were taken with pH papers from every soil sample. The range in pH was between 3.6 and 5.5 in all the regions. All samples were set up at the same time and received the same conditions and treatment during the germination period.
Data on geoedaphic and geographic factors per habitat were identified using GIS. Data on the major categories of bedrock types were identified from geological maps (Geological Survey of Norway 2004), and classified as poor, moderate, or rich in bedrock nutrient content based on Tarbuck and Lutgens (1992). Data on slope and altitude were identified from the 25 × 25 m pixel terrain model. There were generally only small differences in the geoedaphic and geographic factors between districts with a history of stable and increased reindeer density. However, bedrock nutrient content showed a gradient from rich bedrock in the region west, intermediately rich in region middle to mostly poor bedrock in the region east. Similarly, slope and altitude showed a gradient from steeper slopes at higher altitudes (average of approx. 18° and 350 m asl) in the region west, to intermediate steep slopes at intermediate altitudes (average of approx. 13° and 275 m asl) in the region middle, to less steep slopes and lower altitudes (average of approx. 8° and 200 m asl) in the region east.
Preparation of data for statistical analysis
Germination data were converted to seed density by calculating the seed density per m2. Soil seed bank and top layer samples were summed together because of the large amount of zeros present in the soil seed bank data. From now on they are referred to as the seed bank data. Germinating seeds were classified after three life history traits. Each life history trait was divided in several trait groups: (1) Growth form (graminoid, herbaceous dicotyledon, shrub); (2) Seed mass (<0.1 mg, >0.1 to 0.5 mg, >0.5 to 1 mg, >1 to 5 mg); (3) Diaspore morphologies [no appendages, animal dispersed appendages (hooks, fruits and elaiosome), wind dispersed appendages (wings and pappus)]. Data on seed mass and diaspore morphology were extracted from several internet data bank sources (Fitter and Peat 1994; Klotz et al. 2002), literature (Welling and Laine 2002; Welling et al. 2004), and personal observation. Growth form classification was based on Chapin et al. (1996) with some modifications. Due to the low number of species germinating, we grouped sedges and grasses as one group, called graminoids, while all herbaceous dicotyledons were another group. See Appendix in Supplementary material for a classification of the germinating species after the different traits.
Statistical analysis
All data were sampled at the plot level and were then averaged to habitat since this level was the focus of sampling. Data were transformed by loge(x + 1) and analyzed using linear mixed effects models (lme; Pinheiro et al. 2005). Lme models allow the usage in nested designs of explanatory variables that are a mixture of fixed effects and random effects (Pinheiro and Bates 2000). We tested for main effects and possible interactions of grazing history and life history traits on seed bank densities (seeds/m2). A separate analysis was carried out for each of the three focal life history traits (growth form, seed mass categories, and diaspore morphology), always including grazing history as the other main predictor variable in all analyses. We also included in all analyses region as a covariate, since we expected seed densities to differ according to context differences between regions. Grazing history was a categorical variable with two levels: increased or stable. Region had three categories: region west, region middle, and region east. In all analyses, district and landscape area were considered random effects. However, district was eventually dropped as a random factor as it explained very little variation (var = 0.05 ± 0.01) and this spatial level corresponds to the spatial level of grazing history and region.
All statistical output is presented as geometric means, i.e., back transforming results from the logarithmic scale. Data analyses were performed in the statistical environment R (version 2.4.0; R Development Core Team 2006).
Results
Out of 2,056 individuals that germinated from the seed bank samples, 70% were identified to species level and classified according to the selected traits to run the analyses. The mean seed density in the seed bank per habitat was 1,268 ± 470 (95% CI, n = 70) seeds/m2 and 62 species were encountered (see Appendix).
The results from the three statistical analyses showed that the seed bank density (seeds/m2) per habitat was significantly affected by grazing history and region, and that seed density differed significantly between growth forms, seed mass categories, and diaspore morphologies (Table 1). Habitats in districts with a history of increased reindeer density had lower seed bank densities (Fig. 3). Between the regions, the seed bank densities were higher in the region middle and the region west than in the region east (Fig. 3). The distribution of life history traits was independent of history of reindeer grazing (Table 1), while there was an interaction with region (Table 1). For instance, the region east had a higher proportion of seeds that are wind dispersed and of shrub growth form (Table 2). Overall, small seeds (<0.5 mg), seeds with no appendages, and seeds belonging to graminoid growth form were the most common traits found in the seed bank (Fig. 4). The estimated effects of the random factors in the mixed models were generally low between the landscape areas compared to habitats within landscape areas for all life history traits (standard deviation for landscape areas vs. habitat within landscape areas were 1.89 vs. 9.18 for growth forms, 1.86 vs. 8.58 for seed mass categories, and 2.03 vs. 9.91 for diaspore morphologies).

Mean seed density (seeds per m2; geometric mean with 95% confidence intervals) for the two different grazing histories and for the three different Finnmark regions

Mean seed density (seeds per m2; geometric mean with 95% confidence intervals) for the different growth forms, and seed traits, i.e., seed mass categories and diaspore morphologies
Discussion
Districts with a history of increased reindeer densities had lower seed density in the seed bank as opposed to districts where reindeer densities had remained stable. However, no indications of selective pressure on life history trait distribution were found due to differences in this recent grazing history. Overall, individuals with a graminoid growth form, small seeds (<0.5 mg), and seeds lacking a mechanism for seed dispersal were the most common traits present in the seed banks from this alpine grazed ecosystem. These are traits common among grazing tolerant plants, and may reflect that the vegetation has adapted to a longer grazing history in Finnmark, where reindeer have been present for several centuries (Skogland 1994). Both seed density and the life history trait compositions were found unevenly distributed across the regions, indicating dependency on contexts varying at the large spatial scale of this study (Fig. 3; Table 1).
Lower seed densities in districts with a history of increased reindeer density indicate that the recruitment opportunities of this alpine vegetation are reduced. The decrease in seed density is likely because reindeer graze in a selective manner, by choosing plant reproductive parts when available (Klein 1990; Bråthen and Oksanen 2001; Cooper and Wookey 2003). Furthermore, there is also evidence that high grazing pressure not only decreases the amount of flowers, but also reduces the investment on sexual reproduction in many reindeer forage species (Ehrlén 1997; Wada 1999; Bråthen and Junttila 2006). Many alpine areas are known to be already seed limited (Turnbull et al. 2000; Eskelinen and Virtanen 2005) and recent studies on geese (Kuijper et al. 2006) and Svalbard reindeer (Cooper 2006) have discovered intensive grazing in the High Arctic to have depleted the seed banks, thus having a lasting impact in these plant communities.
Parallel with the grazing history effect, we found a regional gradient in seed density from region west and middle to region east. This regional gradient in seed density could be attributed to factors such as slope (Csontos et al. 2004), altitude (Molau and Larsson 2000; Cummins and Miller 2002), the physical environment of the soil (Chambers et al. 1991), or climate (Hill and Vander Kloet 2005). For instance, even small increases in temperature can accelerate flowering of some alpine species (Arroyo et al. 1981; Molau et al. 2005) increasing seed production and consequently increasing seed bank densities.
However, the recent history of reindeer grazing did not appear to be a selective force behind the distribution of the different life history traits. Similar trait dominance was found in both stable and increased districts. The general pattern reflected in both seed banks from increased and stable districts seemed to correspond to a more classic overall grazed ecosystem, i.e., small seeds lacking dispersal mechanisms (Navie and Rogers 1997) and mainly of graminoid growth form. Several graminoid species are known for their capacity of compensatory or even overcompensatory growth under grazing pressure (Wegener and Odasz 1997; Bråthen and Odasz-Albrigtsen 2000). Thus, the several centuries’ of long history of reindeer grazing in Finnmark have probably been a major factor in shaping the seed bank trait composition.
Inconsistently with our results, a recent study on reindeer in Finnmark found more graminoid and forb seedlings emerging in areas where reindeer densities were high (Olofsson 2006). However, results in Olofsson (2006) are based on findings close to reindeer fences while we found no effect of reindeer on graminoid seed density when embracing the summer pastures at their full extent. Thus, our results reinforce the importance of ecological context and scale. It is also worth noting that for vegetation composition responses to reindeer grazing in Finnmark, results found in a study at local scale (Olofsson et al. 2001) are contradicted by a study at larger scale (Bråthen et al. 2007b). Furthermore, reindeer dispersal of seeds by endozoochory has, in Finnmark, been found to be highly dependent on ecological scale and context (Bråthen et al. 2007a). Consequently, rather than choosing only one environmental context for our study, we believed it was important to know how seed bank densities were affected over a range of different environmental contexts for reindeer grazing. Among all studies on herbivore effects on seed banks, our study is the only one in line with the theoretical understanding that context-dependency can be approached by study scaling and design (Wiens 1989; Hobbs 2003; Noda 2004).
Finally, because we have used a study design where reindeer history effects are studied at a scale that includes different ecological contexts, we believe our results to conservatively explain reindeer density effects on the seed banks. We suggest more effort should be made at studying the importance of differing ecological contexts and their interactions on seed banks. For instance, different contexts may be of high relevance for the effect of climate change on seed banks. Large-scale studies, such as the current, should then be of high priority.
References
Anonymous (2004) Ressursregnskap for reindriftsnæringen 1945–2003. Alta, Norway, p 135 (in Norwegian)
ArcGIS (2004) ESRI GIS and mapping software. Version 8.3.0
Arroyo MTK, Armesto JJ, Villagran C (1981) Plant phenological patterns in the high Andean Cordillera of central Chile. J Ecol 69:205–223
Batzli GO, White RG, MacLean SF, Pitelka FA, Collier BD (1980) The herbivore-based trophic system In: Brown J, Miller RG, Tieszen LL, Bunnell FL (eds) An Arctic ecosystem: the coastal tundra at Barrow, Alaska. US/IBP Synthesis Series. Dowden, Hutchinson and Ross, Stroudsburg, pp 335–410
Bråthen KA, Junttila O (2006) Infertile times: response to damage in genets of the clonal sedge Carex bigelowii. Plant Ecol 187:83–95
Bråthen KA, Odasz-Albrigtsen AM (2000) Tolerance of the arctic graminoid Luzula arcuata ssp. confusa to simulated grazing in two nitrogen environments. Can J Bot 78:1108–1113
Bråthen KA, Oksanen J (2001) Reindeer reduce biomass of preferred plant species. J Veg Sci 12:473–480
Bråthen KA, Gonzalez VT, Iversen M, Killengreen S, Ravolainen VT, Ims RA, Yoccoz NG (2007a) Endozoochory varies with ecological scale and context. Ecography 30:308–320
Bråthen KA, Ims RA, Yoccoz NG, Fauchald P, Tveraa T, Hausner V (2007b) Induced shift in ecosystem productivity? Extensive scale effects of abundant large herbivores. Ecosystems 10:773–789
Chambers JC, Macmahon JA (1994) A day in the life of a seed: movements and fates of seeds and their implications for natural and managed systems. Annu Rev Ecol Syst 25:263–292
Chambers J, Macmahon J, Haefner J (1991) Seed entrapment in alpine ecosystems—effects of soil particle-size and diaspore morphology. Ecology 72:1668–1677
Chapin FS, BretHart MS, Hobbie SE, Zhong HL (1996) Plant functional types as predictors of transient responses of arctic vegetation to global change. J Veg Sci 7:347–358
Chippindale HG, Milton WEJ (1934) On the viable seeds present in the soil beneath pastures. J Ecol 22:508–531
Cooper EJ (2006) Reindeer grazing reduces seed and propagule bank in the high Arctic. Can J Bot 84:1740–1752
Cooper EJ, Wookey PA (2003) Floral herbivory of Dryas octopetala by Svalbard reindeer. Arct Antarct Alp Res 35:369–376
Cornelissen J, Quested H, Gwynn-Jones D, Van Logtestijn R, De Beus M, Kondratchuk A, Callaghan T, Aerts R (2004) Leaf digestibility and litter decomposability are related in a wide range of subarctic plant species and types. Funct Ecol 18:779–786
Csontos P, Tamas J, Podani J (2004) Slope aspect affects the seed mass spectrum of grassland vegetation. Seed Sci Res 14:379–385
Cummins R, Miller GR (2002) Altitudinal gradients in seed dynamics of Calluna vulgaris in eastern Scotland. J Veg Sci 13:859–866
Diaz S, Noy-Meir I, Cabido M (2001) Can grazing response of herbaceous plants be predicted from simple vegetative traits? J Appl Ecol 38:497–508
Edenius L, Vencatasawmy CP, Sandstrom P, Dahlberg U (2003) Combining satellite imagery and ancillary data to map snowbed vegetation important to reindeer Rangifer tarandus. Arct Antarct Alp Res 35:150–157
Ehrlén J (1997) Risk of grazing and flower number in a perennial plant. Oikos 80:428–434
Eriksson A, Eriksson O (1997) Seedling recruitment in semi-natural pastures: the effects of disturbance, seed size, phenology and seed bank. Nord J Bot 17:469–482
Eskelinen A, Virtanen R (2005) Local and regional processes in low-productive mountain plant communities: the roles of seed and microsite limitation in relation to grazing. Oikos 110:360–368
Fauchald P, Tveraa T, Yoccoz NG, Ims RA (2004) En økologisk bærekraftig reindrift Hva begrenser naturlig produksjon og høsting? NINA Fagrapport 76 (in Norwegian)
Fitter A, Peat H (1994) The ecological flora database. J Ecol 82:415–425. www.ecoflora.co.uk
Geological Survey of Norway (2004) Digital maps of bedrock and superficial deposits in Troms and Finnmark 1:250000
Giba Z, Gribisic D, Konjevic C (2003) Nitrogen oxides as environmental sensors for seeds. Seed Sci Res 13:187–196
Hanssen-Bauer I (1999) Klima i nord de siste 100 år. Ottar 99:41–48 (in Norwegian)
Hill NM, Vander Kloet SP (2005) Longevity of experimentally buried seed in Vaccinium: relationship to climate, reproductive factors and natural seed banks. J Ecol 93:1167–1176
Hobbs NT (2003) Challenges and opportunities in integrating ecological knowledge across scales. For Ecol Manag 181:223–238
Ims RA, Fuglei E (2005) Trophic interaction cycles in tundra ecosystems and the impact of climate change. Bioscience 55:311–322
Jutila H (1998) Seed banks of grazed and ungrazed Baltic seashore meadows. J Veg Sci 9:395–408
Klein DR (1990) Variation in quality of caribou and reindeer forage plants associated with season, plant part and phenology. Rangifer Spec Issue 3:123–130
Klein DR (1999) The role of climate and insularity in establishment and persistence of Rangifer tarandus populations in the high Arctic. Ecol Bull 47:96–104
Klotz S, Kühn I, Durka W (2002) Biolflor—eine datenbank zu biologisch-ökologischen merkmalen der gefäßpflanzen in deutschland. Schriftenreihe für Vegetationskunde 38. Bundesamt für Naturschutz, Bonn. www.biolflor.de
Kruckeberg AR (2002) Geology and plant life: the effects of landforms and rock types on plants. University of Washington Press, Seattle
Kuijper DPJ, Bakker JP, Cooper EJ, Ubels R, Jonsdottir IS, Loonen MJJE (2006) Intensive grazing by barnacle geese depletes high Arctic seed bank. Can J Bot 84:995–1004
Larsson E-L, Molau U (2001) Snowbeds trapping seed rain: a comparison of methods. Nord J Bot 21:385–392
Lid J, Lid DT (1994) Norsk flora. Det Norske Samlaget, Olso
Mayor MD, Bóo RM, Peláez DV, Elía OR (2003) Seasonal variation of the soil seed bank of grasses in central Argentina as related to grazing and shrub cover. J Arid Environ 53:467–477
McIntyre S, Lavorel S, Landsberg J, Forbes T (1999) Disturbance response in vegetation—towards a global perspective on functional traits. J Veg Sci 10:621–630
Molau U, Larsson E-L (2000) Seed rain and seed bank along an alpine altitudinal gradient in Swedish Lapland. Can J Bot 78:728–747
Molau U, Nordenhäll U, Eriksen B (2005) Onset of flowering and climate variability in an alpine landscape: a 10-year study from Swedish Lapland. Am J Bot 92:422–431
Muga DA (1986) A commentary on the historical transformation of the Sami communal mode of production. J Ethn Stud 14:111–121
Naess MW, Fauchald P, Tveraa T (2009) Scale dependency and the “marginal” value of labor. Hum Ecol 37:193–211
Navie SC, Rogers RW (1997) The relationship between attributes of plants represented in the germinable seed bank and stocking pressure in a semi-arid subtropical rangeland. Aust J Bot 45:1055–1071
Noda T (2004) Spatial hierarchical approach in community ecology: a way beyond high context-dependency and low predictability in local phenomena. Popul Ecol 46:105–117
Norwegian Meteorological Institute (2006). www.met.no
O’Connor TG, Pickett GA (1992) The influence of grazing on seed production and seed-banks of some African savannah grasslands. J Appl Ecol 29:247–260
Olofsson J (2006) Short- and long-term effects of changes in reindeer grazing pressure on tundra heath vegetation. J Ecol 94:431–440
Olofsson J, Kitti H, Rautiainen P, Stark S, Oksanen L (2001) Effects of summer grazing by reindeer on composition of vegetation, productivity and nitrogen cycling. Ecography 24:13–24
Osem Y, Perevolotsky A, Kigel J (2006) Size traits and site conditions determine changes in seed bank structure caused by grazing exclusion in semiarid annual plant communities. Ecography 29:11–20
Pinheiro J, Bates D (2000) Mixed-effects models in S and S-plus. Springer, New York
Pinheiro J, Bates D, DebRoy S, Sarkar D (2005) Nlme: linear and nonlinear mixed effects models R package version 3.1–6.4
R Development Core Team (2006) R: a language and environment for statistical computing. R Foundation for Statistical Computing, Vienna. www.R-project.org
Rusch GM, Pausas JG, Leps J (2003) Plant functional types in relation to disturbance and land use: introduction. J Veg Sci 14:307–310
Senft RL, Coughenour MB, Bailey DW, Rittenhouse LR, Sala OE, Swift DM (1987) Large herbivore foraging and ecological hierarchies. Bioscience 37:789–799
Shadish WR, Cook TD, Campbell DT (2002) Experimental and quasi-experimental designs for generalized causal inference. Houghton Mifflin, Boston
Skogland T (1994) Villrein: fra urinnvåner til miljøbarometer. Teknologisk Forlag, Oslo (in Norwegian)
Sternberg M, Gutman M, Perevolotsky A, Kigel J (2003) Effects of grazing on soil seed bank dynamics: an approach with functional groups. J Veg Sci 14:375–386
Tarbuck EJ, Lutgens FK (1992) The earth: an introduction to physical geology. Macmillan, New York
Thompson K, Bakker J, Bekker R (1997) The soil seed banks of North West Europe: methodology, density and longevity. Cambridge University Press, Cambridge
Turnbull LA, Crawley MJ, Rees M (2000) Are plant populations seed-limited? A review of seed sowing experiments. Oikos 88:225–238
Tveraa T, Fauchald P, Yoccoz NG, Ims RA, Aanes R, Høgda KA (2007) What regulate and limit reindeer populations in Norway? Oikos 116:706–715
Wada N (1999) Factors affecting the seed-setting success of Dryas octopetala in front of Brøggerbreen (Brøgger glacier) in the high arctic, Ny Ålesund, Svalbard. Polar Res 18:261–268
Walker DA, Raynolds MK, Daniels FJA, Einarsson E, Elvebakk A, Gould WA, Katenin AE, Kholod SS, Markon CJ, Melnikov ES, Moskalenko NG, Talbot SS, Yurtsev BA, Team C (2005) The circumpolar arctic vegetation map. J Veg Sci 16:267–282
Wardle D, Barker G, Bonner K, Nicholson K (1998) Can comparative approaches based on plant ecophysiological traits predict the nature of biotic interactions and individual plant species effects in ecosystems? J Ecol 86:405–420
Wegener C, Odasz AM (1997) Effects of laboratory simulated grazing on biomass of the perennial arctic grass Dupontia fisheri from Svalbard: evidence of overcompensation. Oikos 79:496–502
Welling P, Laine K (2002) Regeneration by seeds in alpine meadow and heath vegetation in sub-arctic Finland. J Veg Sci 13:217–226
Welling P, Tolvanen A, Laine K (2004) The alpine soil seed standing vegetation bank in relation to field seedlings and in subarctic Finland. Arct Antarct Alp Res 36:229–238
White RC, Trudell J (1980) Habitat preference and forage consumption by reindeer and caribou near Atkasook, Alaska. Arct Antarct Alp Res 12:511–529
Wiens JA (1989) Spatial scaling in ecology. Funct Ecol 3:385–397
Acknowledgments
We would like to thank the Norwegian Coast Guard, Leidulf Lund, Jarle Nilsen, and Kjell Nilsen for their help at the phytotron, Torstein Engelskjøn for help with species identification, Asle Lilletun, all the participants of Ecosystem Finnmark, and Tina Dahl, Alina Evans and Mette Nielsen for field assistance and Rolf Anker Ims and Nigel Yoccoz for discussions on the study design. This study is part of the “Ecosystem Finnmark” project, which was financed by the Norwegian Research Council.
Author information
Authors and Affiliations
Corresponding author
Electronic supplementary material
Below is the link to the electronic supplementary material.
Rights and permissions
About this article
Cite this article
González, V.T., Bråthen, K.A., Ravolainen, V.T. et al. Large-scale grazing history effects on Arctic-alpine germinable seed banks. Plant Ecol 207, 321–331 (2010). https://doi.org/10.1007/s11258-009-9676-2
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11258-009-9676-2