
Abstract
Despite the accelerating global urbanization and its associated implications for wildlife and humans, we know little about the biology of urban ecosystems. Here, we investigated colonization and habitat selection of the European hare (Lepus europaeus), a declining farmland species, in urban areas in Denmark, using a combination of citizen science data and transect counts. Further, we estimated the population density of urban hares in Aarhus, Denmark’s second largest city. Our results provide the first evidence that hares have established populations in urban areas, potentially in response to decreasing habitat quality in rural areas due to agricultural intensification. The hare density in Aarhus was ca. 8 hares per km2, which is comparable to or slightly higher than hare abundance estimates from various pastural areas in Europe, suggesting that urban areas provide suitable habitat for hares. Hare habitat selection was generally associated with areas consisting of large lawns, such as high buildings and parks, which potentially provide high-quality forage throughout the year. Considering the increasing expansion of urban areas and deteriorating habitat quality of agricultural areas, urban planning that incorporates habitat requirements for wildlife could help to support urban animal populations, especially for species of conservation concern.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Urbanization increases globally and is a major driver of environmental change (Grimm et al. 2008). For example, the majority of carbon emissions, residential water use, and industrial timber use are attributed to cities (Brown 2001), negatively affecting ecosystems globally. Many animal populations go extinct in built-up areas, because they are not able to adapt to the rapidly changing conditions (McKinney 2006). However, urban areas can constitute habitat for various wildlife populations that were previously restricted to rural environments (Ditchkoff et al. 2006). In fact, population densities of some species can be higher in urban compared to surrounding agricultural areas, e.g. due to increased food availability and thermal shelter, as shown for hedgehogs (Erinaceus europaeus) (Hubert et al. 2011). The process of urban colonization and adjustment of animal populations to specific conditions of the urban environment is defined as synurbanization (Luniak 2004). Although there is an increasing research focus on urban environments (Magle et al. 2012), we still know little about the biology of urban species, such as the mechanisms of colonization (Rutz 2008), adaptations to urban environments (Lowry et al. 2013), and habitat selection (Chambers and Dickman 2002; Marks and Bloomfield 2006). The ultimate reason for synurbanization of animal populations is the expansion of urban areas (Luniak 2004), whereas proximate mechanisms include behavioral adaptations towards novel habitats and human disturbance (Honda et al. 2018). Generally, individuals with a high behavioral flexibility are more likely to settle in urban environments, and in effect, urban wildlife often shows different behaviors compared to individuals in rural environments (Lowry et al. 2013). Further, wildlife might settle in urban areas after an increase in population size, as shown for red fox (Vulpes vulpes) (Wandeler et al. 2003) and wild boar (Sus scrofa) (Stillfried et al. 2017), or due to improved habitat conditions in urban areas (Altwegg et al. 2014), e.g. increased food availability and reduced predation risk. Conversely, decreasing habitat quality or human persecution in rural areas might force individuals to colonize urban areas (Rutz 2008).
Especially for uncommon or declining native animals living in urban environments, the identification of preferred habitats can be an invaluable conservation tool, given the expansion of urbanization. Citizen science can be a useful tool to collect large amounts of data, but has its challenges due to varying quality of the data, variation in sampling effort both in space and time, and sampling bias (Dickinson et al. 2010). Urban areas are well suited for citizen science studies due to the high density of potential observers and the high accessibility via a dense road network, reducing variation in sampling effort. For example, citizen science data was used to understand human-coyote (Canis latrans) encounters in urban areas (Wine et al. 2015) and to map occupancy of Eastern screech owls (Megascops asio) in suburban areas (Nagy et al. 2012).
Here, we investigated the colonization history and habitat selection of the European hare (Lepus europaeus, hereafter hare, Fig. S1) in urban areas in Denmark to shed light on the mechanisms driving the synurbanization of farmland species. Hares evolved in open steppe grassland and now predominantly occur in agricultural landscapes, being more abundant in arable compared to pastoral areas (Vaughan et al. 2003). Home range size usually varies between 10 and 100 ha depending on resource availability (Tapper and Barnes 1986; Ullmann et al. 2018), and hares feed on a variety of plant species, including most crops and many wild plants (Frylestam 1986), whereas weeds, grasses, and crops rich in fat are generally selected for (Schai-Braun et al. 2015). The main predator of hares is the red fox (Vulpes vulpes) (Knauer et al. 2010). Hare populations have declined throughout Europe since 1960 (Smith et al. 2005). The main cause of this decline is attributed to agricultural intensification, such as increased field sizes and more monocultures, reducing habitat heterogeneity, and widespread pesticide use, reducing the availability of wild herbs and weeds (Edwards et al. 2000; Smith et al. 2005; Storkey et al. 2011). Further, industrial meat production has led to the farming of more arable crops for animal feed at the expense of pastures (Foley et al. 2011; Peyraud et al. 2014), and high vegetation (such as cereals and maize) was shown to exclude hares from agricultural fields during parts of the year (Mayer et al. 2019; Mayer et al. 2018), potentially bringing them in closer proximity to human settlements. Hares in agricultural areas generally avoid paved roads (Roedenbeck and Voser 2008), and a simulation study indicated that urbanization might decrease hare abundance (Lundström-Gilliéron and Schlaepfer 2003). However, apart from a master thesis (Schulze 2012), to our knowledge no published record of hare occurrence and habitat selection in urban areas exists.
Initially, we investigated correlates of urban colonization by hares, testing different hypothesis that are not mutually exclusive to understand the mechanisms driving hare synurbanization. We used citizen science data to track the colonization of urban areas and tested the hypotheses that urban colonization was related to agricultural intensification, increasing human observers in urban areas, changing hare population size, and the expansion of urban areas. Further, we estimated the hare abundance and density in Aarhus, Denmark’s second largest urban area, using transect counts. Moreover, using citizen science data and transect counts, we describe hare occurrence in urban areas in Denmark and investigated the factors affecting habitat selection in urban areas. We predicted that hares were more likely to occur in green, open and comparatively larger areas resembling agricultural landscape, such as parks and areas with large lawns, allowing for both foraging opportunities and predator detection. We discuss the implications of our findings for urban planning and conservation of hares.
Materials and methods
Study area and landscape data
Our study area comprised all major urban areas of Denmark, defined as all urban morphological zones derived from the European Environment Agency (http://ftp.eea.europa.eu/www/umz/v4f0/UMZ2000.zip) that were > 10 km2 in size (Fig. 1). To describe changes in rural land use and farming intensity over time, we obtained data regarding the number of farms in Denmark, the total agricultural area, and the area of cereal fields from Statistics Denmark (https://www.dst.dk/). Further, we obtained the proportion of the human population living in urban areas from the World Bank database (https://data.worldbank.org/indicator/SP.URB.TOTL.IN.ZS?locations=DK) and the annual rate of change of urban areas (in percent), defined as urbanization rate, from the United Nations database (https://population.un.org/wup/Download/). To get a measure of the Danish hare population size over time, we used annual hunting bag data (http://fauna.au.dk/jagt-og-vildtforvaltning/vildtudbytte/udbyttet-online-siden-1941/soejlediagram/). To describe urban areas, we obtained data of land cover types, main roads, buildings, and property sizes from the Danish Map Supply (https://www.kortforsyningen.dk/). Further, we obtained the normalized difference vegetation index (NDVI) as measure of green vegetation obtained from remote sensing 30-m-resolution images from the Landsat archive (http://earthexplorer.usgs.gov) and provided by Engemann et al. (2019) (https://drive.google.com/drive/folders/1FRAU-6UhAsUEw2dzvxntLMAzRmrdo72c). Land cover types were categorized into forest, industrial areas, areas with low buildings, areas with high buildings, city centers, water bodies, and parks (Table 1). Forests were characterized by very large property sizes and NDVI values. The category ‘parks’ mostly consisted of areas without buildings, such as parks, sport fields, railroads, fallow land etc., and consisted of comparatively large property sizes and NDVI values. Areas with high buildings were characterized by intermediate property sizes and NDVI values. Industrial areas were of comparable property sizes than high buildings, but had lower NDVI values. City centers and areas with low buildings consisted of small property sizes and low (city centers) and high (areas with low buildings) NDVI values. Finally, we obtained the human population density at a 1 × 1 km resolution from the Central Person Register (www.cpr.dk).

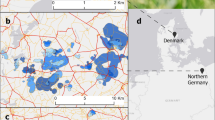
Maps of Aarhus, Denmark. Map a shows property sizes (grey lines), line transects (black lines) and European hare observations during transect counts (black dots). The small map shows all large urban areas in Denmark (grey shading). Map b shows the 1 × 1 km grid cells (black lines), land cover types (see color code in figure legend) and citizen science hare observations (black dots)
Urban hare observations
To obtain a measure of hare abundance and population density, we conducted line transect counts in Aarhus, Denmark’s second largest city (273 K inhabitants). Data sampling was conducted in April and May 2018. We walked line transects on foot just after dusk, when hares are most active (Schai-Braun et al. 2012), because other methods, such as spotlight counts from a vehicle, were not feasible in a city. Sixty-one transects, ranging between 1.0 and 3.1 km in length (mean ± SD: 2.1 ± 0.4 km, sum = 127.7 km), were laid randomly along the course of roads and streets, ranging over the entire city (Fig. 1). The transects were representative of the land cover types of Aarhus, i.e., the proportion of each land cover type covered by the transects was comparable to the respective proportion of that land cover type in the entire city (Table S1). Individual transects were walked by a single observer at normal walking pace (ca. 3–5 km/h). The exact location of each observed hare was recorded using a handheld GPS. Further, we obtained hare observations from two citizen science databases, naturbasen.dk and the Global Biodiversity Information Facility (GBIF.org (04 June 2019) GBIF Occurrence Download https://doi.org/10.15468/dl.mrqqtu). The database ‘naturbasen’ is a countrywide database for the collection of biodiversity data in Denmark. Citizens can report species observations using an APP (since 2012), which automatically records the location of an observation, or alternatively through a web-based module at a home computer. Records are continually validated by the aid of photographic documentation and a panel of mainly amateur experts. GBIF is an international database, providing open access data about all types of life on earth.
Data preparation and statistical analyses
We conducted four main analyses (Table S2): (1) investigating possible causes for urban colonization using citizen science data, (2) estimating the abundance and population density of hares based on transect counts, (3) analyzing the factors affecting number of hares per urban grid cell using citizen science data, and (4) analyzing habitat selection of urban hares using both citizen science data and transect counts.
Correlates of urban colonization
To analyze the correlates of urban colonization by hares we estimated the proportion of hare observations in urban areas compared to all observations separately for each year from when citizen science data were available (from 1978). We used this measure to control for potential variation in sampling effort between years. We used a generalized linear model (GLM) with a binomial response variable and a logit link to analyze the proportion of hares in urban areas. As independent variables, we used the number of farms in Denmark, the total area of agricultural land and the area of cereal as measure of agricultural intensification, the proportion of the human population living in urban areas as a measure of changing sampling effort, the urbanization rate as measure of increasing urbanization, and the annual hunting bag numbers as measure of hare population size. Most of these variables were highly correlated (Spearman rank correlation r > 0.6). Thus, we created single-effect models and compared them using Akaike Information Criterion corrected for small sample size (AICc) (Arnold 2010). We scaled and centered all independent variables to be able to compare the relative effect sizes. After we found that the proportion of urban hare observations could have been driven by declining hare populations in rural areas, we also analyzed the number of urban hare observations (dependent variable) using GLMs with a quasipoisson response and a log link to test the robustness of the our previous findings. We used the same independent variables and additionally included the total human population size to account for changing potential observer numbers.
Density of hares in Aarhus city
To avoid biases in the probability of detection based on the distance from the observer, we corrected hare abundance using distance sampling (Fig. 2). Our line transects met the assumptions of distance sampling, i.e., complete detection of objects on the transect line, that objects do not move before detection (hares generally did not react to pedestrians passing them; MM, personal observation) and that measurements are exact (Thomas et al. 2010). The perpendicular distance of each hare location to the line transect was calculated using the R package ‘raster’ (Hijmans et al. 2015). Line transect density estimates with their log-normal confidence intervals were computed using the R package ‘Distance’ (Miller et al. 2016). We did not truncate data, because we had no extreme outliers. The maximum observed distance from the transect was 90 m (Fig. S2). Thus, we set the observed transect width to 100 m, which is likely an overestimation of the surveyed area (due to buildings reducing visibility). We used three a priori models: a uniform key function, a half-normal key function, and a Hazard rate key function, all with cosine adjustment. We initially also fitted the models with a polynomial and a Hermite polynomial adjustment, but excluded them due to worse model fit based on AICc. We used a goodness of fit test for distance sampling models using the R package ‘mrds’ (Laake et al. 2015) to assess model fit.

The proportion of urban European hare observations compared to all observations (a), and the number of hare observations in Denmark during the different months of the year separately for rural and urban areas (b). Hare observations were obtained from citizen science data in Denmark
Number of hares per urban grid cell
To avoid large variation in data quality, we only selected citizen science hare locations that had a spatial accuracy of <250 m. Further, to standardize the spatial sampling effort and to obtain a relative measure of hare occurrence and abundance for the citizen science data, we created 1 × 1 km large grid cells over all urban areas, and counted the number of hares in each grid cell using the R package ‘rgeos’ (Bivand et al. 2018). Within each grid cell, we then calculated the proportion of each land cover type, the density of main roads (km of road per km2), the average property size, the mean area of all buildings, the building density (area of buildings per km2), the mean NDVI, and the human population size, using the R package ‘raster’ (Hijmans et al. 2015).
To analyze the number of hares per grid cell (dependent variable), we used generalized linear mixed models (GLMM) with a negative binomial distribution and a log link to account for overdispersion and zero-inflation of the count data, using the r package glmmADMB (Bolker et al. 2012; O’hara and Kotze 2010). We included the proportion of areas with high and low buildings, respectively, city centers, industry, forest and parks, the road density, average NDVI, human population density and grid cell size (which varied at the edges of urban areas) as fixed effects (the latter two variables were included to control for different sampling effort), and the urban ID as random intercept.
Habitat selection in urban areas
To analyze habitat selection within urban areas based on citizen data, we selected only urban areas that had sufficient hare observations (> 20). To get a measure of resource availability, we created 10 x the number of random positions than we had obtained from citizen science hare observations within each urban area. We then assigned each random and used (hare) position to the land cover type, property size and NDVI using the R package ‘raster’ (Hijmans et al. 2015). We did the same procedure for the hare data obtained from the transect counts. However, in this case, we defined our sampling area for random locations as the transects buffered by 100 m, because this was the area we could potentially observe hares, i.e. not to include areas that had not been sampled.
To analyze habitat selection (hare location = 1 versus random location = 0, dependent variable), we used GLMs for the transect count data and GLMMs of the R package ‘lme4’ (Bates et al. 2015) for the data obtained from citizen science data. We used different models, because we only analyzed one urban area (Aarhus) in the case of the transect counts, and multiple areas in the case of the citizen science data, and thus included the urban ID as random intercept in the latter analysis to control of non-independence of the data. Both analyses were run with a binomial distribution and a logit link. We included the land cover type (excluding water), NDVI, and property size (log-transformed to normalize residuals of the statistical models) as fixed effects. We had no observations of hares in forests in our transect count data and thus excluded this land cover type from the analysis (transect count analysis only). Further, we did not include the proximity to roads in the analysis as the transects were laid on roads and we thus had a higher detection probability in proximity to roads, and because citizen science data was too inaccurate and potentially also biased towards roads. We initially created single-effect models for the variables property size and NDVI to test if their relationship with the response was linear or quadratic, based on AIC values.
Model selection
For analyses 3 and 4, we initially tested for correlations between the independent variables (we created dummy variables for categorical variables), with correlated variables defined as Spearman rank correlation >0.6 (Zuur et al. 2010). We then created single effect models for the correlated variables, and removed the variable with lower fit, i.e. higher AICc value (Zuur et al. 2010). Model selection for all analyses was based on a stepwise variable selection using AICc, selecting the model with the lowest AICc (Murtaugh 2009), using the R package ‘MuMIn’ (Barton 2016). Parameters that included zero within their 95% CI were considered uninformative (Arnold 2010). We validated the most parsimonious models by plotting the model residuals versus the fitted values (Zuur et al. 2009). All statistical analyses were carried out in R 3.2.5 (R Core Team 2013).
Results
Correlates of urban colonization
We obtained a total of 38,389 hare observations from citizen science data between 1978 and 2019. Of these, 2,925 observations came from urban areas (7.6% of observations). No hares were observed in urban areas before the year 1991, and the proportion of hare observations in urban areas increased from 0.4% in 1991 to 13.5% in 2018 (Fig. 2). The proportion of hare observations in urban areas was best explained by the number of farms (delta AICc to the next model = 55.9), i.e., the proportion of urban hare observations increased with decreasing number of farms (Table 2). However, the other single-effect models were also informative, suggesting that the proportion of urban hare observations also increased with the proportion of people living in urban areas and to a smaller degree the urbanization rate, and decreased with the hare hunting bag (as measure of hare population size) and the area of total agricultural land and cereal (Table 2). The number of urban hares was also best explained by the number of farms, whereas all other variables (same as above) were also informative (Table 2). The number of farms roughly halved from 1980 to 2018, whereas the area of total agricultural land and cereals was relatively stable (Fig. S2). The proportion of people living in urban areas increased from 84% in 1982 to 88% in 2018, and hare hunting bag size dropped from about 200,000 to 38,000 hares in the same period (Fig. S2).
Population density of hares in Aarhus city
We observed 62 hares on 61 transects (Fig. 1). All three key functions were within delta AICc <2, with the best model being the one with a uniform key function (Fig. 3). Model fit was good (χ2 goodness-of-fit test; χ2 = 3.58, df = 5, p = 0.61). The model estimate was 151 hares (SE = 20.6, 95% confidence interval (CI): 110–192 hares) in the covered area (22.2 km2 when using a search distance of 100 m). This corresponded to a hare density of 6.8 hares per km2 (95% CI: 5.0–8.6) and a total population size of 584 hares (95% CI: 428–739) in Aarhus (covering an area of 85.6 km2). Excluding the area covered by buildings (which cannot be utilized by hares) led to a population density estimate of 8.1 hares per km2 (95% CI: 5.9–10.2).

Detection model (line) with a Uniform key function and a cosine for European hares in Aarhus, Denmark. The histogram and black dots represent the observed detection distances of hares
Number of hares per grid cell
To analyze the number of hares per urban grid cell, we used data from 2009 to 2018. We obtained 1,261 hare observations from urban areas with an accuracy of <250 m. Urban hares were observed throughout the year, with most observations coming from the brighter months between April and September, which was also the case for rural hare observations (Fig. 2). Observations from the ‘naturbasen.dk’ database included an age category (adult versus juvenile) for each observation, and the percentage of juvenile hare observations in urban areas was 1.6% (10 of 619 observations), which was comparable to rural areas (1.9%; 73 of 3,727 observations).
The number of hare observations per grid cell ranged from 0 to 44 observations (mean ± SD = 0.5 ± 2.2; median = 0). Building density was correlated with the proportion of areas with high buildings and consequently removed from the analysis. Further, the average property size was negatively correlated with the proportion of areas with low buildings and parks and thus removed from the analysis. The number of hares increased with the proportion of areas with high buildings, city center, industrial areas and parks, the road density, and with the grid cell area (Table 3, Fig. 4). The quadratic function of NDVI revealed that the number of hares first decreased with increasing NDVI and then levelled at a mean NDVI of zero (Table 3, Fig. 4). Apart from the grid cell size, the proportion of areas with high buildings and parks had the largest effect on the number of hares (Table 3). Human population density, the proportion of areas with low buildings and the proportion of forest were not included in the best model.

The estimated number of European hares per grid cell (solid line) in relation to the (a) proportion of areas with high buildings, (b) proportion of parks, and (c) mean NDVI. The 95% confidence intervals are shown as grey shading. Data were obtained from citizen science data between 2009 and 2018
Habitat selection
The habitat selection of hare locations obtained from our transect counts was best explained by the land cover type and the interaction of NDVI and property size. With areas with low buildings as reference, hares selected for areas with high buildings and parks, whereas industrial areas were neither selected for nor avoided (Table 4). When properties were large (>10 ha), hares increasingly avoided greener areas (as expressed by NDVI; Table 4, Fig. 5). Conversely, when property sizes were smaller (<10 ha), hares increasingly selected for greener areas (Table 4, Fig. 5).

The effect of the interaction between property size and NDVI on habitat selection by European hares in Aarhus, Denmark. The 95% confidence intervals are given as grey shading. Data were obtained from transect counts in 2018
Habitat selection based on citizen science observations was best explained by the land cover type and quadratic function of property size (Table 4). With areas with low buildings as reference, hares selected for areas with high buildings, industrial areas and parks, and did neither select nor avoid city centers and forests (Table 4). Relative to random locations, hares selected for areas with high buildings, industrial areas and parks, and avoided areas with low buildings (Fig. 6). There was no selection for or against city centers or forests (Fig. 6). Further, hares selected for intermediate sized properties, and avoided smaller and larger properties (Table 4, Fig. 6). The effect of NDVI was uninformative.

The effect of (a) the log-transformed property size and (b) land cover type on habitat selection by European hares in urban areas in Denmark. The 95% confidence intervals are given as grey shading (a) and bars (b). Data were obtained from citizen science data between 2009 and 2018
Discussion
To our knowledge, we are the first to describe the colonization and habitat selection of hares in urban areas. Hares were observed in all major cities of Denmark, and we provide evidence that urban colonization correlated with agricultural intensification. The hare population density in Aarhus was estimated at 6.8 hares per km2 or 8.1 hares per km2 when excluding the area of buildings (i.e., areas inaccessible to hares), which is comparable to pastural areas in various parts of Europe (Smith et al. 2005). Hares mostly occurred and selected for areas with high buildings and parks (i.e., areas with comparatively large property sizes and lawns) and avoided the greenest areas, i.e., forests and low buildings.
Potential mechanisms for urban colonization
As all independent variables were informative, no single explanation can be identified as the driver behind hare colonization in urban areas. Our most parsimonious model suggests that urban colonization was driven by agricultural intensification as the proportion of urban hare observations increased with agricultural intensification. The best proxy for agricultural intensification (and among all candidate models) was the number of farms, which steadily dropped over the last decades while the total area of agricultural area remained comparatively stable. This means that land use by individual farms intensified via increased agricultural field sizes, and the use of fertilizer and pesticides (Tscharntke et al. 2012), which led to a decline of hares and farmland birds in rural areas in Denmark (Heldbjerg et al. 2017; Schmidt et al. 2004) and elsewhere in Europe (Smith et al. 2005; Zellweger-Fischer et al. 2011). This is also indicated by the declining hunting bag numbers over time, although hunting bag statistics might not be a reliable estimate for population size (Kahlert et al. 2015). Previous research has shown that high vegetation (mainly cereal, maize and rape) excludes hares from agricultural fields, and more so when field sizes are larger (Mayer et al. 2018). This might be another proximate cause for synurbanization (apart from decreased habitat quality), because it brings hares into closer proximity to human settlements during the pre-harvest period when vegetation is generally high. Additionally, the increased proportion of urban hare observations could have partly been caused by an increasing proportion of people living in urban areas, leading to an increased number of potential observers in urban areas. However, the proportion of people living in urban areas increased by only 4% over the last 40 years, making it doubtful that this variable is biologically relevant. However, it is possible that the number of citizen science observers increased more in urban compared to rural areas. Moreover, the structure of urban areas both regarding housing density and vegetation cover might have changed over the last decades, as shown in urban areas in Australia (Luck et al. 2009), potentially providing better habitat for hares and thus facilitating colonization. However, the properties in Denmark have been subdivided in the end of the twentieth century, leading to an increased housing density (Attwell 2000; Guttu et al. 1997), which based on our habitat selection results would indicate that urban habitat quality rather declined over time. Further, lawns made up the majority of the vegetation cover in Danish urban areas already in in the end of the twentieth century and likely earlier (Attwell 2000), suggesting that urban areas had suitable habitat before the colonization by hares after 1990. The only hypothesis we can reject, is that urban colonization by hares was driven by increasing hare numbers in rural areas, because Danish hare populations declined over the last decades (Schmidt et al. 2004). Finally, there is no evidence that predator abundance in urban areas has declined over time, since urban fox populations have been stable (Copenhagen) or increasing (Aarhus) since 1990 (Pagh 2008). No data is available on the abundance of free-ranging cats, although the percentage of households with cats, estimated at 21% (Sandøe et al. 2018), probably has been fairly stable over time.
Population density
Our hare population density estimate for Aarhus (ca. 8 hares per km2) is comparable to hare density estimates from different pastural areas in Europe (Smith et al. 2005) and even slightly higher than estimates from pastural areas in Switzerland (Schai-Braun et al. 2013; Zellweger-Fischer et al. 2011). Further, our estimates were higher compared to two arable areas in Jutland, Denmark (3–6 hares per km2) (Wincentz Jensen 2009). This suggests that urban areas are at least of equal habitat quality compared to many pastural areas across Europe. However, the hare density in Aarhus was much lower compared to arable and mixed areas in other parts of Europe, where densities were on average 28 and 43 hares per km2, respectively (Smith et al. 2005). Usually hare abundance is estimated using spotlight counts from a car (Zellweger-Fischer et al. 2011) or via hunting bag statistics (Schmidt et al. 2004). Being in a city, we could not use spotlights and thus had to focus our sampling effort during dusk while there was still enough light to observe hares. This might have influenced the detection probability of hares, potentially leading to an underestimation of our hare density estimate. Further, differences in building density and height between the different urban land cover types might have influenced the detectability of hares, which also might have affected our hare density estimate.
Number of hares per urban grid cell and habitat selection
The number of hares per urban grid cell as well as habitat selection based on both data sets showed that hares selected for areas with high buildings. Areas with high buildings and parks generally have large property sizes and their vegetation predominantly consists of lawns without other plants (apart from occasional trees). This land cover type potentially resembles agricultural areas better compared to the other urban land cover types, which might explain the hares’ occurrence in and selection for these areas. In these urban areas, lawns are present throughout the year and are regularly mowed, providing high-quality forage due to the short vegetation height (Wilmshurst et al. 1995). Similarly, in an urban environment in Australia, long-nosed bandicoots (Perameles nasuta) used grassy areas for foraging (Chambers and Dickman 2002). In contrast, food availability in arable agricultural areas varies vastly throughout the year (Schai-Braun et al. 2015), depending on vegetation height and farming practice (ploughing, sowing, pesticide and fertilizer spraying, harvesting, etc.), making this habitat much less predictable. Further, short vegetation (lawns) and large property sizes allow for the early detection of approaching objects, allowing hares to outrun potential predators (Hewson 1977; Ogen-Odoi and Dilworth 1984; Weterings et al. 2016). Additionally, the number of hares decreased with increasing NDVI, which makes sense, because high NDVI values were associated with forests and low buildings, areas that were generally not selected for. This was also indicated by the results of our habitat selection analysis (based on transect counts), i.e., in larger properties hares increasingly selected for areas lower NDVI (thus, avoiding forests and selecting for areas with high buildings). Conversely, in smaller properties, hares increasingly selected for higher NDVI values, potentially because the majority of comparatively smaller properties consisted of city center, areas with low and high buildings, and NDVI was lowest in city canters, areas that were not selected for by hares. An additional explanation why hares might not select for areas with low buildings could be that dogs and cats are more abundant in those areas owned by single families (in many large buildings pets are not allowed).
Both observations from transect counts and citizen science data were potentially biased towards active hares. Thus, the patterns described here likely relate to the habitat selection of active hares, and we can only speculate which habitat is selected for by resting hares in urban areas. Personal observations (MM) suggest that hares partly also use large grassy areas for resting during the day. Additionally, depending on the season and weather conditions (Mayer et al. 2019), inactive hares might select for areas with high vegetation as cover, such as small gardens (low buildings) with hedgerows and high vegetation cover, similar to selection in agricultural areas (Neumann et al. 2012; Pépin and Angibault 2007). For example, long-nosed bandicoots were shown to use dense scrub vegetation as resting places in urban areas (Chambers and Dickman 2002). GPS studies within urban areas could resolve the question where hares rest while being inactive, and where they give birth.
The citizen science data was likely biased, because active observers are not evenly distributed over the urban areas and because the number of active observers differed between years. However, we obtained similar results from our transect count data, which was obtained in a more systematic way. Thus, we argue that citizen science data can be a useful tool to study the occurrence and habitat selection of animals in urban areas.
Future perspectives and conclusions
Our results indicate that hares have established populations in urban areas. The question arises whether urban populations are only sinks that take up hares from surrounding populations which experience decreasing habitat quality and thus, increased emigration. Longer-term studies could answer the question if urban populations are increasing (as suggested by our results from the citizen science observations) and GPS-studies could shed light on dispersal and home range size of hares, shedding light on whether such populations are viable. We have limited evidence that hares reproduce in urban areas, but we have no data on mortality rates, which might be substantially higher in urbanized areas due to an increased probability of animal-vehicle collisions and potentially higher densities of predators. Citizen science data indicated that hares reproduce in urban areas as juvenile hares were observed, and their proportion of all observations was comparable to rural areas. Further, we also observed juvenile individuals in Aarhus (MM, personal observation).
Agricultural areas become increasingly hostile for wild animals and plants due to intensification of land use (Emmerson et al. 2016; Storkey et al. 2011; Tsiafouli et al. 2015). At the same time, urban areas are expanding and often have constitute a more heterogeneous landscape than rural areas. Thus, wildlife populations could benefit from urban planning that includes habitat requirements of animals. Parks play a vital role to provide habitat for various species adapted to urban areas (Fernandez-Juricic and Jokimäki 2001; Parker and Nilon 2008), as do residential gardens, especially if they constitute predominantly of native plants (Narango et al. 2018). In the case of the European hare, we found that comparatively large properties with extensive lawns provide suitable urban habitat. In order to aid urban wildlife populations, managers could create heterogeneous urban areas, containing both open areas providing foraging opportunities for animals relying on grassy areas, such as hares, geese, and hedgehogs, and at the same time provide more structured areas with vegetation cover, providing shelter for resting animals. Future research should also investigate occurrence patterns of multiple species and their interactions (Magle et al. 2012).
Change history
23 May 2020
In our paper, using distance sampling, we estimated the population density of hares in Aarhus city to 6.8 hares/km2.
References
Altwegg R, Jenkins A, Abadi F (2014) Nestboxes and immigration drive the growth of an urban Peregrine Falcon Falco peregrinus population Ibis 156:107–115
Arnold TW (2010) Uninformative parameters and model selection using Akaike's information criterion the. J Wildl Manag 74:1175–1178
Attwell K (2000) Urban land resources and urban planting—case studies from Denmark. Landsc Urban Plan 52:145–163
Barton K (2016) Package “MuMIn”: multi-model inference. R package, version 1.15. 6. Accessed,
Bates D et al. (2015) Package ‘lme4’
Bivand R et al. (2018) Package ‘rgeos’ R package v 03–24
Bolker B, Skaug H, Magnusson A, Nielsen A (2012) Getting started with the glmmADMB package available at glmmadmb r-forge r-project org/glmmADMB pdf
Brown L (2001) Building an economy for the earth earth policy institute
Chambers LK, Dickman CR (2002) Habitat selection of the long-nosed bandicoot, Perameles nasuta (Mammalia, Peramelidae), in a patchy urban environment. Austral Ecology 27:334–342
Dickinson JL, Zuckerberg B, Bonter DN (2010) Citizen science as an ecological research tool: challenges and benefits. Annu Rev Ecol Evol Syst 41:149–172
Ditchkoff SS, Saalfeld ST, Gibson CJ (2006) Animal behavior in urban ecosystems: modifications due to human-induced stress. Urban ecosystems 9:5–12
Edwards P, Fletcher M, Berny P (2000) Review of the factors affecting the decline of the European brown hare, Lepus europaeus (Pallas, 1778) and the use of wildlife incident data to evaluate the significance of paraquat Agriculture, ecosystems & environment 79:95-103
Emmerson M et al. (2016) How agricultural intensification affects biodiversity and ecosystem services. In: Advances in Ecological Research, vol 55. Elsevier, pp 43-97
Engemann K, Pedersen CB, Arge L, Tsirogiannis C, Mortensen PB, Svenning J-C (2019) Residential green space in childhood is associated with lower risk of psychiatric disorders from adolescence into adulthood. Proceedings of the National Academy of Sciences 116:5188–5193
Fernandez-Juricic E, Jokimäki J (2001) A habitat island approach to conserving birds in urban landscapes: case studies from southern and northern Europe. Biodiversity & Conservation 10:2023–2043
Foley JA et al (2011) Solutions for a cultivated planet. Nature 478:337–342
Frylestam B (1986) Agricultural land use effects on the winter diet of brown hares (Lepus europaeus Pallas) in southern Sweden. Mammal Rev 16:157–161
Grimm NB, Faeth SH, Golubiewski NE, Redman CL, Wu J, Bai X, Briggs JM (2008) Global change and the ecology of cities. Science 319:756–760
Guttu J, Nyhuus S, Saglie I, Thorén AH (1997) Boligfortetting i Oslo. Konsekvenser for grønnstrukturer, bokvaliteter og arkitektur. NIBR Prosjektrapport,
Heldbjerg H, Sunde P, Fox AD (2017) Continuous population declines for specialist farmland birds 1987-2014 in Denmark indicates no halt in biodiversity loss in agricultural habitats. Bird Conservation International 28:1–15
Hewson R (1977) Food selection by brown hares (Lepus capensis) on cereal and turnip crops in north-East Scotland. Journal of Applied Ecology 14:779–785
Hijmans RJ et al. (2015) Package ‘raster’ R package
Honda T, Iijima H, Tsuboi J, Uchida K (2018) A review of urban wildlife management from the animal personality perspective: The case of urban deer. Science of the total environment 644:576–582
Hubert P, Julliard R, Biagianti S, Poulle M-L (2011) Ecological factors driving the higher hedgehog (Erinaceus europeaus) density in an urban area compared to the adjacent rural area. Landscape and Urban Planning 103:34–43
Kahlert J, Fox AD, Heldbjerg H, Asferg T, Sunde P (2015) Functional Responses of Human Hunters to Their Prey—Why Harvest Statistics may not Always Reflect Changes in Prey Population Abundance. Wildlife biology 21:294–303
Knauer F, Küchenhoff H, Pilz S (2010) A statistical analysis of the relationship between red fox Vulpes vulpes and its prey species (grey partridge Perdix perdix, brown hare Lepus europaeus and rabbit Oryctolagus cuniculus) in Western Germany from 1958 to 1998. Wildl Biol 16:56–66
Lowry H, Lill A, Wong BB (2013) Behavioural responses of wildlife to urban environments. Biological reviews 88:537–549
Luck GW, Smallbone LT, O’Brien R (2009) Socio-economics and vegetation change in urban ecosystems: patterns in space and time. Ecosystems 12:604
Lundström-Gilliéron C, Schlaepfer R (2003) Hare abundance as an indicator for urbanisation and intensification of agriculture in Western Europe. Ecological modelling 168:283–301
Luniak M (2004) Synurbization–adaptation of animal wildlife to urban development. In: Proc. 4th Int. Symposium Urban Wildl. Conserv. Tucson. Citeseer, pp 50–55
Laake J, Borchers D, Thomas L, Miller D, Bishop J (2015) Mrds: mark–recapture distance sampling. R package version 2.1. 12
Magle SB, Hunt VM, Vernon M, Crooks KR (2012) Urban wildlife research: past, present, and future. Biol Conserv 155:23–32
Marks CA, Bloomfield TE (2006) Home-range size and selection of natal den and diurnal shelter sites by urban red foxes (Vulpes vulpes) in Melbourne. Wildlife Research 33:339–347
Mayer M, Ullmann W, Heinrich R, Fischer C, Blaum N, Sunde P (2019) Seasonal effects of habitat structure and weather on the habitat selection and home range size of a mammal in agricultural landscapes Landscape Ecology:1–16
Mayer M, Ullmann W, Sunde P, Fischer C, Blaum N (2018) Habitat selection by the European hare in arable landscapes: The importance of small-scale habitat structure for conservation. Ecology and Evolution 8:11619–11633
McKinney ML (2006) Urbanization as a major cause of biotic homogenization. Biol Conserv 127:247–260
Miller DL, Rexstad E, Thomas L, Marshall L, Laake JL (2016) Distance sampling in R BioRxiv:063891
Murtaugh PA (2009) Performance of several variable-selection methods applied to real ecological data. Ecology letters 12:1061–1068
Nagy C, Bardwell K, Rockwell RF, Christie R, Weckel M (2012) Validation of a citizen science-based model of site occupancy for eastern screech owls with systematic data in suburban New York and Connecticut. Northeastern Naturalist 19:143–159
Narango DL, Tallamy DW, Marra PP (2018) Nonnative plants reduce population growth of an insectivorous bird. Proceedings of the National Academy of Sciences 115:11549–11554
Neumann F, Schai-Braun S, Weber D, Amrhein V (2012) European hares select resting places for providing cover Hystrix, the Italian Journal of Mammalogy 22
O’hara RB, Kotze DJ (2010) Do not log-transform count data. Methods in Ecology and Evolution 1:118–122
Ogen-Odoi AA, Dilworth T (1984) Effects of grassland burning on the savanna hare-predator relationships in Uganda. African Journal of Ecology 22:101–106
Pagh S (2008) The history of urban foxes in Aarhus and Copenhagen. Denmark Lutra 51:51
Parker TS, Nilon CH (2008) Gray squirrel density, habitat suitability, and behavior in urban parks. Urban Ecosyst 11:243–255
Pépin D, Angibault JM (2007) Selection of resting sites by the European hare as related to habitat characteristics during agricultural changes. European Journal of Wildlife Research 53:183–189
Peyraud J-L, Taboada M, Delaby L (2014) Integrated crop and livestock systems in Western Europe and South America: a review. European Journal of Agronomy 57:31–42
R Core Team (2013) R: A language and environment for statistical computing
Roedenbeck IA, Voser P (2008) Effects of roads on spatial distribution, abundance and mortality of brown hare (Lepus europaeus) in Switzerland. Eur J Wildl Res 54:425–437
Rutz C (2008) The establishment of an urban bird population. J Anim Ecol 77:1008–1019
Sandøe P, Nørspang AP, Kondrup SV, Bjørnvad CR, Forkman B, Lund TB (2018) Roaming companion cats as potential causes of conflict and controversy: A representative questionnaire study of the Danish public. Anthrozoös 31:459–473
Schai-Braun SC, Reichlin TS, Ruf T, Klansek E, Tataruch F, Arnold W, Hackländer K (2015) The European hare (Lepus europaeus): a picky herbivore searching for plant parts rich in fat. PLoS One 10:e0134278
Schai-Braun SC, Rödel HG, Hackländer K (2012) The influence of daylight regime on diurnal locomotor activity patterns of the European hare (Lepus europaeus) during summer. Mammalian Biology-Zeitschrift für Säugetierkunde 77:434–440
Schai-Braun SC, Weber D, Hackländer K (2013) Spring and autumn habitat preferences of active European hares (Lepus europaeus) in an agricultural area with low hare density. Eur J Wildl Res 59:387–397
Schmidt NM, Asferg T, Forchhammer MC (2004) Long-term patterns in European brown hare population dynamics in Denmark: effects of agriculture, predation and climate. BMC Ecol 4:15
Schulze R (2012) Development of a quantification method for European brown hares (Lepus europaeus) in urban areas on the example of Lichtenberg. GRIN Verlag, Berlin
Smith RK, Jennings NV, Harris S (2005) A quantitative analysis of the abundance and demography of European hares Lepus europaeus in relation to habitat type, intensity of agriculture and climate. Mammal Rev 35:1–24
Stillfried M et al (2017) Do cities represent sources, sinks or isolated islands for urban wild boar population structure? J Appl Ecol 54:272–281
Storkey J, Meyer S, Still KS, Leuschner C (2011) The impact of agricultural intensification and land-use change on the European arable flora. Proceedings of the Royal Society B-Biological Sciences 279:1421–1429
Tapper S, Barnes R (1986) Influence of farming practice on the ecology of the brown hare (Lepus europaeus). Journal of Applied Ecology 23:39–52
Thomas L et al (2010) Distance software: design and analysis of distance sampling surveys for estimating population size. Journal of Applied Ecology 47:5–14
Tscharntke T et al (2012) Global food security, biodiversity conservation and the future of agricultural intensification. Biol Conserv 151:53–59
Tsiafouli MA, Thébault E, Sgardelis SP, de Ruiter PC, van der Putten W, Birkhofer K, Hemerik L, de Vries FT, Bardgett RD, Brady MV, Bjornlund L, Jørgensen HB, Christensen S, Hertefeldt TD, Hotes S, Gera Hol WH, Frouz J, Liiri M, Mortimer SR, Setälä H, Tzanopoulos J, Uteseny K, Pižl V, Stary J, Wolters V, Hedlund K (2015) Intensive agriculture reduces soil biodiversity across Europe. Glob Chang Biol 21:973–985
Ullmann W, Fischer C, Pirhofer-Walzl K, Kramer-Schadt S, Blaum N (2018) Spatiotemporal variability in resources affects herbivore home range formation in structurally contrasting and unpredictable agricultural landscapes. Landscape Ecology 33:1–13. https://doi.org/10.1007/s10980-018-0676-2
Vaughan N, Lucas EA, Harris S, White PC (2003) Habitat associations of European hares Lepus europaeus in England and Wales: implications for farmland management. Journal of Applied Ecology 40:163–175
Wandeler P, Funk SM, Largiader C, Gloor S, Breitenmoser U (2003) The city-fox phenomenon: genetic consequences of a recent colonization of urban habitat. Molecular Ecology 12:647–656
Weterings MJ, Zaccaroni M, van der Koore N, Zijlstra LM, Kuipers HJ, van Langevelde F, van Wieren SE (2016) Strong reactive movement response of the medium-sized European hare to elevated predation risk in short vegetation. Animal Behaviour 115:107–114
Wilmshurst JF, Fryxell JM, Hudsonb RJ (1995) Forage quality and patch choice by wapiti (Cervus elaphus). Behavioral Ecology 6:209–217
Wincentz Jensen T-L (2009) Identifying causes for population decline of brown hare (Lepus europaeus) in agricultural landscapes in Denmark Unpublished PhD thesis, NERI, Aarhus University, Denmark
Wine S, Gagné SA, Meentemeyer RK (2015) Understanding human–coyote encounters in urban ecosystems using citizen science data: what do socioeconomics tell us? Environ Manag 55:159–170
Zellweger-Fischer J, Kéry M, Pasinelli G (2011) Population trends of brown hares in Switzerland: the role of land-use and ecological compensation areas. Biological conservation 144:1364–1373
Zuur A, Ieno E, Walker N, Saveliev A, Smith G (2009) Mixed effects models and extensions in ecology with R. Gail M, Krickeberg K, Samet JM, Tsiatis A, Wong W, editors New York, NY: Spring Science and Business Media
Zuur AF, Ieno EN, Elphick CS (2010) A protocol for data exploration to avoid common statistical problems. Methods in ecology and evolution 1:3–14
Acknowledgements
We thank Anders Fedder Kristensen, Carsten Swayne Storgaard, David Bækby Houborg, Emma Kaczmar, Hanne Juel Christiansen, Jonas Robert Andersen, and Mathias Damholt for assistance with fieldwork. Further, we thank the Danish Hunters Association for support and an anonymous reviewer for constructive feedback. We are grateful for economical funding form the Danish Environmental Agency.
Author information
Authors and Affiliations
Contributions
Martin Mayer: Conceptualization; Data collection; Formal analysis; Methodology; Visualization; Writing - original draft. Peter Sunde: Funding acquisition; Methodology; Project administration; Writing - review & editing.
Corresponding author
Ethics declarations
Conflict of interest
The authors declare no competing financial, personal or other conflict of interests.
Electronic supplementary material
ESM 1
(DOCX 1967 kb)
Rights and permissions
About this article

Cite this article
Mayer, M., Sunde, P. Colonization and habitat selection of a declining farmland species in urban areas. Urban Ecosyst 23, 543–554 (2020). https://doi.org/10.1007/s11252-020-00943-1
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11252-020-00943-1