
Abstract
Urbanization is thought to lead to the loss of biodiversity both because of habitat disturbance and the increased abundance of invasive species. However, most studies of biodiversity in cities are conducted on a short time scale, usually less than 3 years, and so miss the long-term dynamics of communities inhabiting these ecosystems. Here we use a study performed in the early 70’s on North Carolina State University (Raleigh, USA) as a baseline to evaluate the long term effects of disturbance and introduced species on native ant communities. Ant species were sampled almost 40 years later using a variety of sampling techniques in order to maximize species collection. Our results show that while the number of exotic species increased, including three major invasive ants, native ant species richness remained high. Furthermore, our survey was able to add several new records for the area considered, in comparison of the 70’s study, for a total of 89 species known from NCSU campus. After comparison with other studies, our results represent one of the most species-rich urban environments monitored and thus open encouraging perspective on how urban ecosystems could contribute to the preservation of the biodiversity of small-bodies organisms such as ants.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Over the past decades the number and size of cities continuously increased (Seto et al. 2011) and urbanized areas are predicted to triple by 2030 (relative to 2000) (Seto et al. 2012). Many authors have expressed concern over the fate of native biodiversity in these urban areas (Seto et al. 2012; McIntyre 2000; Chace and Walsh 2006; McKinney 2008), the same areas where the public is increasingly most likely to experience biodiversity (Dunn et al. 2006). Natural areas in urban areas are often disturbed relative to more natural habitats (McKinney 2002), with consequences for biodiversity. Most studies have emphasized on the importance of spatial components such as area sizes, disturbance levels or how the different types of urban habitats (residential, urban parks, forest remnants) sustain species diversity and abundance (Menke et al. 2011). Urbanization also facilitates the spread of introduced species (McIntyre 2000; McKinney 2008; Chytrý et al. 2008; Pyšek et al. 2010) which can strongly affect native species communities (Bolger et al. 2000). Urban environments combine both habitat disturbance (Rebele 1994) and biological invasions, inarguably the two most important drivers of the loss of global biodiversity (Vitousek et al. 1997).
Clearly, in urban areas, biodiversity has the potential to be victim to the juggernaut of disturbance and biological invasions. However, urban habitats are also characterized by heterogeneity (Savard et al. 2000). This heterogeneity may offer refuge to many species unable to persist in the “average” urban conditions (McKinney 2008). While many large animals, such as mammals or birds can require large habitat patches, small patches may be sufficient for the smaller organisms that constitute the majority of species (McKinney 2008). In this context, it is not surprising that the effects of urbanization on plant and animals communities are diverse (McIntyre 2000; McKinney 2008). To the extent that predictions are possible, we might predict that smaller organisms might tend to be less negatively affected by urbanization than larger organisms. For bird communities, urbanization typically leads to decrease species richness, an increase of biomass and a shift toward generalist species (Chace and Walsh 2006). For other groups of vertebrates, most studies tend to observe a general decrease of species richness with increasing urbanization levels (McIntyre 2000; McKinney 2008). Ant communities found within urban environments are usually considered to have impoverished in native species richness (Bernard 1958; Dauber 1997; Antonov 2008), though studies usually compare only a subset of habitat types rather than searching exhaustively for the persistence (or lack thereof) of species. Because smaller species may need smaller habitats for persistence they may also, almost by definition, be easier to miss.
Invasive species are recognized as one of the main factors responsible for the reduction or disappearance of specific native species, whether in light of disturbance such as urbanization or more generally (Elton 1958; Mack et al. 2000). In ants, several of the well-studied invasive ant species thrive in urban and other disturbed habitats (Holway et al. 2002). Invasive species such as these might preempt the ability of urban habitats to maintain biodiversity. On the other hand, King and Tschinkel (2008) have also argued that the decrease of abundance and richness observed in native communities is sometimes more strongly associated with the effect of disturbance than the effects of invasion, at least in the case of Solenopsis invicta. Long term datasets within urban settings have the potential to allow the through time comparison of the effects of invasion and disturbance on native species. Independent of the effects of invasive species on native species, McIntyre (2000) hypothesizes that change in invasive species assemblages themselves might be predictable, with the species accumulation of exotic species increasing with time. This pattern has already observed for other groups of organisms, especially plants (Tait et al. 2005).
One of the biggest limitations in our understanding of the response of species to urbanization, be they native or introduced, is the dearth of long-term studies. This is particularly true for insects for which museum collections and historic records are typically sparse. For instance in a review of 79 studies by McIntyre (2000), no study looked at the effect of urbanization for more than three consecutive years. This represents a real problem as the short and long-term effects of urbanization may be very different. For this reason, early studies of the biota in cities represent very valuable benchmarks of the effects of urbanizations, particularly if data from those studies can be compared to more recent samples. From 1971 to 1973 Nuhn and Wright (1979) studied the ant community present on the campus of North Carolina State University in downtown Raleigh, North Carolina through the use of extensive collection methods (hand collecting, pitfall traps, Berlese traps and Malaise trap) and reported the presence and relative commonness of fifty nine species including five exotic species, including the invasive ant Pachycondyla chinensis Emery (Guénard and Dunn 2010). Forty years later, we resampled the same habitats originally considered by Nuhn and Wright (1979). During the intervening years, the population of Raleigh has gone from 125,572 inhabitants in 1971 to over 405,192 inhabitants in 2009 (official city of Raleigh website), and during the same period the density of students increased from 20,000 students to more than 34,000 nowadays (North Carolina State University Historical Sketch website). Due to constant modification of the habitat through construction, landscape management and the presence of new invasive species on the studied area, we hypothesized that the diversity of native ant species should be less than it was 40 years ago, whereas that of introduced species should be greater.
Methods
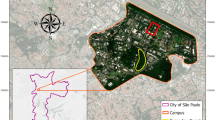
We collected ants on the North Carolina State University (NCSU) campus within the limits of the area defined by Nuhn and Wright (1979) from July 2007 to August 2011 (Fig. 1). The section studied of NCSU campus (35.7865 N, −78.6721 W), represents an area of about 1.3 km2 (or 130 ha), and is located on the south-eastern portion of the city of Raleigh, North Carolina, USA. It is surrounded on its northern, western and southern edges by residential and commercial areas, while its eastern portion is bordered by a city park. North Carolina State University campus is characteristic of urban ecosystem, composed of a heterogeneous mosaic of habitats that include areas heavily built and with intense human activity, several thousands of individuals using the area of study daily, many recreational areas with landscaped vegetation, large isolated trees surrounded by mowed grassy field courts and remnants of forest patches with old-growth trees. Furthermore as for many urban ecosystems (McIntyre 2000), this area experiences perpetual light to heavy disturbances which range from people activity (soil compaction), vegetation landscaping (mowing, pruning, plantation, plant removal, pest treatments …), to the construction or modifications of buildings and infrastructures. To illustrate this, since 1973 (the end of the initial sampling) up to 2006 (the beginning of our sampling), no fewer than seven new buildings have been built and 26 renovations events occurred in the area considered (North Carolina State University Building index website)(Fig 2).

Location of the sampling sites on the northern part of the North Carolina State University campus. The following collection methods were used: baiting stations (small diamonds), hand collection (small circle), pitfall traps (pin symbol) and Winkler extractors (large circle)

Mean number of native ant (gray) and exotic ant (dark) species collected on baits for the different habitats sampled at the grain scale. Standard Error bars are presented for each habitat for native species (gray bars) and exotic species (dark bars). Letters indicate significance among groups for exotic species (on top of graph) and for native species (on the bottom part of graph)
We chose sampling methods to maximize the number of species collected. Toward this end, we used several collection techniques, hand collecting, baiting, pitfall traps and Winkler extractors (Fig. 1). The techniques used are also comparable to those used in Nuhn and Wright (1979): hand collecting, baiting and leaf litter extractions. It should be noted however that we did not use Malaise traps which were used Nuhn and Wright (1979). In addition, we used a passive trapping method not used in the previous study, pitfall traps..
Baiting sampling
Ants were collected at baits from June to August 2011. Ants were surveyed each day in one area of NCSU campus on five different sub-habitats. 1) Group of trees, defined as a remnant of forest presenting at least 5 trees clumped together; 2) Isolated tree, defined as a tree standing isolated from any other tree; 3) Shrubs, defined as ornamental shrubs that provide shade but which do not exceed 2 m in height; 4) Lawn, defined as herbaceous open areas that are regularly mowed; and 5) Pavement, defined as highly modified human habitat covered by concrete or bricks. Each sub habitats were replicated thirteen times and for each site a total of 10 baits stations were used, separated at least by 3 m (and up to 5 m) from each other for a total of 650 bait stations. Baits were left for an hour. Each bait was composed of cookie crumbs within a 15 mL plastic vial (11.5 × 1.5 mm). After an hour, the vials were covered and any ants were frozen. The ants were then sorted and placed in alcohol until they could be identified.
Eight other baiting sites were sampled in September 2009. These additional sites were not used for statistical analysis, but were used for the overall counts of species richness and occurrences.
Winkler extractor sampling
The Winkler extractor method is an efficient method to collect ant species present in the leaf litter and species foraging on the surface (Bestelmeyer et al. 2000). Leaf litters was collected randomly in order to maximize the number of micro-habitats sampled and thus collect a maximum of ant species (Ward 2000). Once sifted, the litter was brought to the laboratory to be placed in mini-Winkler extractor for drying for a length of 4 days. A total of 31 sites were sampled with this technique.
Pitfall traps sampling
Pitfall traps were used only in the patch of old-growth secondary forest in 2007 and 2008. Details about the methods used are provided in Guénard and Dunn (2010). Four sites of 16 pitfall traps were sampled with this technique which represented a total of 64 pitfall traps. Other methods than the pitfall traps were preferred as those are time demanding for a limited efficiency (many leaf litter species are not collected with this method) and difficult to operate in a landscaped habitat (e.g. removal problems). For these reasons, the use of leaf litter collection, which allows a direct collection through the Winkler extractor technique was preferred.
Literature survey
To evaluate the importance of urban ecosystems in preserving diversity, we compare the species richness known from several studies realized in urban habitat to the richness of ant species known from the entire region where the city is located. Species richness values were obtained from relevant publications and when not available for the region considered from a database developed by one of the authors (BG) which includes records from over 5000 publications compiled for the elaboration of a global ant survey project (Guénard et al. 2012a, ongoing projects). This literature database was used to extract data from articles specific to urban environments and for the relevant regional checklist. Number of exotic species present in those studies was calculated from the authors’ knowledge on species global distribution.
Analysis
We compared the mean species richness of native and exotic species collecting by the baiting method for each of the habitat sampled. We use the non-parametric Wilcoxon of multiple comparisons method as the data tested were not normally distributed.
Taxonomic changes and updates
Since the publication of the Nuhn and Wright article in 1979, several groups of ants collected at North Carolina State University have been revised. Those taxonomic updates were incorporated into the text and tables. When available, specimens collected by Nuhn and Wright (1979) and deposited at the NCSU Insect Museum collection were also checked for species identification.
Nuhn and Wright (1979) noted the collection of Camponotus americanus, however upon examination their specimen, deposited at NCSU museum, is actually C. castaneus which we also collected several times in this study. Unless other real specimens of C. americanus have been deposited elsewhere, we consider this species to be absent on the North Carolina State University campus. The presence of this species on this area is nonetheless possible, just not yet confirmed (Table 1). The identification of Tetramorium guineense by Nuhn and Wright (1979) is due to a misidentification of Tetramorium bicarinatum which is known from other parts of North Carolina (Guénard et al. 2012b) and is known to have been often misidentified (Wetterer 2009). The ant Tetramorium caespitum presented in Nuhn and Wright (1979) is likely to be Tetramorium sp. E, an undescribed species of the Tetramorium caespitum complex and introduced from Europe (Schlick-Steiner et al. 2006). However, for readability, we refer to it as T. caespitum, but note should be taken of the uncertain identification of this species. The ant species Ponera exotica, considered as exotic in Nuhn and Wright (1979) is considered here as a native species following the work of (Johnson 1987).
Results
We collected a total of 78 ant species during this survey. Combined with the results of Nuhn and Wright (1979), this represents a total of 89 species collected on the North Carolina State University campus since 1971 (Table 1). Eleven species collected by Nuhn and Wright (1979) were not captured during our sampling. Conversely, 32 of the species we collected were not encountered by Nuhn and Wright (1979) (Table 1).
The number of species collected on baits in the “group of trees” habitat was higher (17 species) than any other habitat type (Table 2). The mean number of native species collected per site in the group of trees habitat was higher than all other habitats except isolated trees (Table S1). The number of exotic species was highest in lawn and paved habitats. The lawn and pavement habitats, both of them highly anthropogenic had the fewest native species and most exotic species.
Exotic and invasive species
A total of 10 exotic species have been reported from North Carolina State University campus and a total of 12 exotic species are known from Wake County more generally (Table 1). Nine exotic species were collected on NCSU campus during this survey. One exotic species collected by Nuhn and Wright (1979), Tetramorium bicarinatum (presented as T. guineense), was not collected during this survey. Two new invasive species, the fire ant, Solenopsis invicta, and the Argentine ant, Linepithema humile which were both absent in the 1970’s are now common species on NCSU campus, especially in open habitats (Table 2). Other exotic species previously collected and still present included the Dacetine ant, Pyramica membranifera (5 collections), the pavement ant, Tetramorium sp. E. (8 collections), and the Asian needle ant, Pachycondyla chinensis (17 collections). This last species was quite common on many locations of campus when tree cover was present (Table 2) in contrast to of Nuhn’s work (1977) in which the species was found just once.
Three exotic species represent new outdoor collections for this site. The fire ant, Solenopsis invicta, and the Argentine ant, Linepithema humile, are now among the most common species found. Brachymyrmex patagonicus was found at only one location, at the base and foraging on a willow oak, but with a high population density of foraging workers. Two species collected only indoors, Pheidole moerens and Hypoponera punctatissima represent new exotic records for NCSU campus. Those two species were probably introduced with plant materials and were restricted to the area surrounding ornamental plants. Pheidole moerens was observed foraging on scale insects growing on plants and also feeding on some Diplopoda found within the soil of plants.
Discussion
For small organisms, urban ecosystems offer a heterogeneous and complex mix of open and canopy-closed habitats of many types, including both elements similar to surrounding habitats, such as patches of trees, and elements that are entirely different, such as patches of cement (McKinney 2008). Thus while the study area considered here is small in absolute size (1.3 km2) it is large enough to include a wide variety of habitat types. In this area, Nuhn and Wright (1979) collected a very diverse ant fauna, in part because of their use of a wide variety of sampling regimes. Since that time, the area considered has been dramatically transformed and is now much more urban and surrounded by a much larger city. For these reasons, one might expect to see a decrease in diversity since 1979. Instead, we found that most species collected in the early 70’s were still present almost 40 years later, and several other species, native and exotic, are now present in this urban area. In most cases, it is impossible to say whether these new species were present in 1979, but undetected by Nuhn and Wright (1979) or if they have colonized this area since the initial study. For a few exotic ants, it seems likely that their introduction in this region occurred between our two studies. The fire ant, Solenopsis invicta is among those. Callcott and Collins (1996) documented the establishment of this species in North Carolina in 1995. Another invasive species, P. chinensis, while present in 1979 was rare, recorded in just one sample. This species is now very common and the comparison of our study and that of Nuhn and Wright (1979) benchmarks the timeline of its spread, which more recent studies (Guénard and Dunn 2010; Rice and Silverman 2013), suggest can be rapid.
A-priori we predicted based on McIntyre (2000) that the number of introduced species would increase through time. While five exotic species were reported by Nuhn and Wright (1979), this number has now doubled 40 years later. The new exotic species reported include the two invasive species L. humile and S. invicta which are considered to have detrimental effects on native ant communities (Holway et al. 2002). The fire ant, S. invicta, was especially common and abundant in open habitat. Specimens collected within more forested habitats tended to be from colonies on the forest edge (personal observation), such that edge effects due to fire ants in the study area may be common. A similar phenomenon was also described for the Argentine ant in the same region (Rowles and Silverman 2009). Brachymyrmex patagonicus was especially abundant at the only site it was collected and represents a new exotic species for NCSU campus. To our knowledge, this record represents the northernmost record of B. patagonicus in North America (MacGown et al. 2007; 2010; Martinez et al. 2011). Tetramorium sp. E was one of the most common and abundant species encountered in Nuhn and Wright (1979) study. While this species is still regularly encountered in our study site (Table S2), our results indicate that this species seems less conspicuous now than 40 years ago. Hypoponera opaciceps and P. membranifera, two hypogaeic species, were collected only through Winkler extractor method, in respectively one and five locations. The presence of P. membranifera in urban habitats is not unusual, and this species was previously recorded from anthropogenic habitats such as public parks in the southern part of Spain (Espadaler and López-Soria 1991; Ordonez-Urbano et al. 2008). Finally, we failed to find Tetramorium bicarinatum, known previously from only one location near greenhouses (Nuhn 1977; Nuhn and Wright 1979) that were destroyed several years ago. However, this species is still present in North Carolina with new records reported recently (Guénard et al. 2012b) and therefore could still be present in the Raleigh area.
Despite differences between the sampling efforts and methods used between this study and that of Nuhn and Wright (1979), some changes in the commonness of species are conspicuous. Probably the most striking result is the quasi disappearance of Lasius neoniger, recorded from a single occurrence represented by 2 collected workers in our study. It was the most common species in the 70’s (Nuhn and Wright 1979). The population size of this species is now greatly reduced, the reasons for which are enigmatic. Here we suggest two hypotheses. Nuhn (1977) noticed the presence of a high rate of infection by a fungus tentatively identified as Laboulbenia formicarum on L. neoniger workers. Presence of fungal infestation of L. neoniger has also been noticed in a population collected about 1 km from the studied area of NCSU campus in August 2011 (B.G. personal observation). While the presence of this fungus on L. neoniger worker is well known (Wheeler 1910), its level of pathogenicity is presumed to be low (Smith 1946; Herraiz and Espadaler 2007) such that fungal die off seems unlikely, although cannot be ruled out. A second possibility is the replacement or displacement of L. neoniger by other species. Lasius neoniger favors open sunny habitats (Carter 1962; Nuhn and Wright 1979) and might have been displaced by the fire ant, S. invicta which now commonly occupy open habitats such as grasslands and pavements (Table 2). Other species that are open-habitat specialists, Forelius pruinosus and Pheidole bicarinata (Carter 1962; Nuhn and Wright 1979), show some evidence of having experienced similar fates. Forelius pruinosus was encountered at only two baits and collected only one other time by hand during the entire sampling, while Pheidole bicarinata, the third most common species in the 1970’s (Nuhn and Wright 1979) was only found in less than 11 % of the baiting sites in our study (Fig. 3). The hypothesis that these species were displaced by S. invicta is supported, albeit anecdotally, by the absence of S. invicta in the area where L. neoniger, F. pruinosus and P. bicarinata have been observed to be common outside campus. Furthermore the negative impact of the fire ant, S. invicta on native communities has been well described (Porter and Savignano 1990; Epperson and Allen 2010). The mysterious decline in the abundance of L. neoniger and other open-habitat species deserves further study.

Comparison of the 8 most common species collected in 1971–73 (from Nuhn and Wright 1979) and in 2009–11. Exotic species are framed in red and native species framed in green. Species are from left to right: 1970’s) Lasius neoniger, Nylanderia sp., Pheidole bicarinata, Pheidole dentata, Tetramorium sp. E, Monomorium minimum, Forelius pruinosus and Prenolepis imparis; 2000’s) Solenopsis invicta, Monomorium minimum, Nylanderia sp., Pachycondyla chinensis, Aphaenogaster carolinensis, Pheidole dentata, Tapinoma sessile, and Solenopsis molesta. Numbers in the 1970’s series represent the total number of occurrences per species for all collecting methods and, in the 2010’s series the percentage of occurrences per species on baits (the calculation of percentage was impossible for the 70’s series as the total number of collections in uncertain). Photos @ Alex Wild & Benoit Guénard
Conversely, several species that were not encountered in the 1970’s are now common on the North Carolina State University campus. In part, this change may be an artifact of improving taxonomy. Some species can now be identified that were difficult or impossible to identify in 1979. Nuhn and Wright (1979), clumped several Aphaenogaster species together (probably A. carolinensis, A. picea, and A. rudis) as well as several species of the genus Nylanderia (then called Paratrechina parvula complex). Secondly, our sampling effort was designed to target principally hypogaeic and epigaeic species living in the leaf litter strata, while at the opposite, the use of leaf litter samples in Nuhn and Wright (1979) was reduced and their sampling emphasis was carried on ground dwelling species. This might explain the new records from campus of hypogaeic species such as Amblyopone pallipes (7 occurrences), Discothyrea testacea (6 occurrences), Proceratium silaceum (6 occurrences), Lasius flavus (2 occurrences), or Pyramica pergandei (2 occurrences), as well as the high frequency of occurrences of Solenopsis molesta (33 occurrences), or of other hypogaeic species such as Hypoponera opacior (24 occurrences), Myrmecina americana (19 occurrences), Pyramica ornata (16 occurrences), Strumigenys louisiannae (16 occurrences) and Ponera pennsylvanica (14 occurrences).
Eleven species collected by Nuhn and Wright (1979) were not captured during our sampling. Among the species not collected in this study, several were collected in 2008 elsewhere in the city of Raleigh (Menke et al 2011). Those include Crematogaster vermiculata, Dorymyrmex bureni, Formica dolosa, Lasius subglaber, Neivamyrmex texanus and Trachymyrmex septentrionalis (Menke et al 2011), and Ponera exotica (unpublished result). It seems likely then that those species might still occur on the North Carolina State University campus or nearby. Little can be said about the other four species collected in 1979 but missing from our collections other than that these species are interesting ones for further study.
Results of baiting indicate that more overall and native species are found in closed-canopy habitats while less native species but more exotic species are encountered in open, heavily disturbed habitats. The importance of trees in disturbed habitats to maintain ant species diversity has been emphasized in several studies (Dunn 2000; Yasuda and Koike 2009). Our studies also support this conclusion and further suggest that the presence of trees may limit the impact of invasive species on native ant communities. While the overall species richness was higher in groups of trees than near isolated trees (Table 2), the mean number of species per sample was similar (Fig. 2).
To our knowledge, the species richness found on North Carolina State University campus, with 89 species, is the most diverse ant fauna known from an urban habitat anywhere in the world with the exception of Bogor, on Java island, Indonesia where 94 species were collected (Rizali et al. 2008). The ants on the North Carolina State University campus represent 79 and 46 % of the species known from Wake County and from North Carolina respectively (Guénard et al. 2012b). These estimates are higher than those from studies of ants in other cities. The number of ant species collected from cities represent 3 to 79 % (mean of 20 %) from the species regional pool known from the entire region where the city is located (Table S3). In part this is likely due to the focus of urban studies of ants. Most studies focus on standardized ecological sampling of ants, but such sampling while useful for comparative purposes is unlikely to find all of the species present (see Espadaler and López-Soria 1991). For example, Menke et al. (2011) sampled Raleigh using a standardized method and found just 53 % of the species we encountered on campus. The high number of species collected in this study suggests that cities may play a more important role in conservation than generally appreciated, a role that could be better understood without much work. Most researchers, interested in ant community ecology and diversity, could easily survey their local ant fauna and compare their results to ours herein.
In conclusion, high diversity of native ant species can persist even in urban environments in which many invasive species are present (Holway et al. 2002; Guénard and Dunn 2010). Such persistence, we speculate, is facilitated by habitat heterogeneity, even if such heterogeneity is at a small spatial grain. The persistence of biodiversity in urban environments may be greater for small organisms such as ants that are not typically the target of conservation. However, because small organisms represent the majority of life by any accounting, this persistence is of great relevance to the conservation of biodiversity more generally.
References
Antonov IA (2008) Ant assemblages of two cities with different ecological conditions in southern Cisbaikalia. Russ J Ecol 39:454–456
Bernard F (1958) Fourmis des villes et fourmis du bled entre rabat et tanger. Bull Soc Sci Nat Maroc 38:131–142
Bestelmeyer BT, Agosti D, Alonso LE, Brandao CRF, Brown WL Jr, Delabie JHC, Silvestre R (2000) Field techniques for the study of ground-dwelling ants: an overview, description, and evaluation. In: Agosti D, Majer JD, Alonso LE, Schultz TR (eds) Ants: standard methods for measuring and monitoring biodiversity. Smithsonian Institution Press, Washington, pp 122–144
Bolger TD, Suarez AV, Crooks KR, Morrison SA, Case TJ (2000) Arthropods in urban habitat fragments in southern California: area, age and edge effects. Ecol Appl 10:1230–1248
Callcott AMA, Collins HL (1996) Invasion and range expansion of imported fire ants (Hymenoptera: Formicidae) in North America from 1918–1995. Fla Entomol 79:240–251
Carter WG (1962) Ants of the north Carolina piedmont. J Elisha Mitch Sci S 78:1–18
Chace JF, Walsh JJ (2006) Urban effects on native avifauna: a review. Landsc Urban Plan 74:46–69
Chytrý M, Jarosik V, Pysek P, Hájek O, Knollová I, Tichý L, Danihelka J (2008) Separating habitat invasibility by alien plants from the actual level of invasion. Ecology 89:1541–1553
Dauber J (1997) Ameisenfauna einer urbanen Landschaft. Naturschutz und Landschaftplanung 29:303–309
Dunn RR (2000) Isolated trees as centers of ant diversity in active and fallow fields. Biol Conserv 95:317–321
Dunn RR, Gavin M, Sanchez M, Solomon J (2006) The pigeon: dependence of global conservation on urban nature. Conserv Biol 20:1814–1816
Elton CS (1958) The ecology of invasions by animals and plants. Methuen, London
Epperson DM, Allen CR (2010) Red imported fire ant impacts on upland arthropods in southern Mississippi. Am Midl Nat 163:54–63
Espadaler X, López-Soria L (1991) Rareness of certain Mediterranean ant species: fact or artifact? Insect Soc 38:365–377
Guénard B, Dunn RR (2010) A new (old), invasive ant in the hardwood forests of Eastern North America and its potentially widespread impacts. PLoS One 5:e11614. doi:10.1371/journal.pone.00116147
Guénard B, Weiser MD, Dunn RR (2012a) Global models of ant diversity suggest regions where new discoveries are most likely are under disproportionate deforestation threat. Proc Natl Acad Sci U S A 109:7368–7373
Guénard B, McCaffrey KA, Lucky A, Dunn RR (2012b) Ants of North Carolina: an updated list (Hymenoptera: Formicidae). Zootaxa 3552:1–36
Herraiz JA, Espadaler X (2007) Laboulbenia formicarum (Ascomycota, Laboulbeniales) reaches the Mediterranean. Sociobiology 50:449–455
Holway DA, Lach L, Suarez AV, Tsutsui ND, Case TJ (2002) The causes and consequences of ant invasions. Annu Rev Ecol Syst 33:181–233
Johnson C (1987) Biogeography and habitats of Ponera exotica (Hymenoptera: Formicidae). J Entomol Sci 22:358–361
King JR, Tschinkel WR (2008) Experimental evidence that human impacts drive fire ant invasions and ecological change. Proc Natl Acad Sci U S A 105:20339–20343
MacGown JA, Hill JG, Deyrup MA (2007) Brachymyrmex patagonicus (Hymenoptera: Formicidae) an emerging pest species in the southeastern United States. Fla Entomol 90:457–464
MacGown JA, Hill JG, Brown LA (2010) Dispersal of the exotic Brachymyrmex patagonicus (Hymenoptera: Formicidae) in the United States. Proceedings: Imported Fire Ant Conference, Charleston, South Carolina, March 24–26, 2008, 80–86
Mack RN, Simberloff D, Lonsdale WM, Evans H, Clout M, Bazzaz FA (2000) Biotic invasions: causes, epidemiology, global consequences, and control. Ecol Appl 10:689–710
Martinez MJ, Wrenn WJ, Tilzer A, Cummings RF (2011) New records for the exotic ants Brachymyrmex patagonicus Mayr and Pheidole moerens Wheeler (Hymenoptera: Formicidae) in California. Pan-Pac Entomol 87:47–50
McIntyre NE (2000) Ecology of urban arthropods: a review and a call to action. Ann Entomol Soc Am 93:825–835
McKinney ML (2002) Urbanization, biodiversity, and conservation. Bioscience 52:883–890
McKinney ML (2008) Effects of urbanization on species richness: a review of plants and animals. Urban Ecosyst 11:161–176
Menke SB, Guénard B, Sexton JO, Weiser MD, Dunn RR, Silverman J (2011) Urban areas may serve as habitat and corridors for dry-adapted, heat tolerant species; an example from ants. Urban Ecosyst 14:135–163
North Carolina State University Building index. http://www.ncsu.edu/facilities/buildings/index.htm. Assessed on February 15 2013
North Carolina State University Historical Sketch. http://www.lib.ncsu.edu/scrc/university-historical-sketch. Accessed on April 26 2014
Nuhn TP (1977) A survey of the ants (Hymenoptera: Formicidae) on the campus of North Carolina State University at Raleigh. Master’s thesis
Nuhn TP, Wright CG (1979) An ecological survey of ants (Hymenoptera: Formicidae) in a landscaped suburban habitat. Am Midl Nat 102:353–362
Official City of Raleigh Website, Raleigh Demographics. http://www.raleighnc.gov/home/content/PlanLongRange/Articles/RaleighDemographics.html. Accessed on August 4th 2013
Ordonez-Urbano C, Reyes-Lopez J, Carpintero-Ortega S (2008) Una especie aloctona puede ser “rara”? El caso de Pyramica membranifera (Hymenoptera: Formicidae). Bol Soc Entomol Aragonesa 42:321–323
Porter SD, Savignano DA (1990) Invasion of polygyne fire ants decimates native ants and disrupts arthropod community. Ecology 71:2095–2106
Pyšek P, Bacher S, Chytrý M, Jarošík V, Wild J, Celesti-Grapow L, Gassó N, Kenis M, Lambdon PW, Nentwig W et al (2010) Contrasting patterns in the invasions of European terrestrial and freshwater habitats by alien plants, insects and vertebrates. Global Ecol Biogeogr 19:317–331
Rebele F (1994) Urban ecology and special features of urban ecosystems. Global Ecol Biogeogr 4:173–187
Rice ES, Silverman J (2013) Propagule pressure and climate contribute to the displacement of Linepithema humile by Pachycondyla chinensis. PLoS One 8:e56281. doi:10.1371/journal.pone.0056281
Rizali A, Bos MM, Buchori D, Yamane S, Hans C, Schulze J (2008) Ants in tropical urban habitats: the myrmecofauna in a densely populated area of Bogor, West Java, Indonesia. Hayati J Biosci 15:77–84
Rowles AD, Silverman J (2009) Carbohydrate supply limits invasion of natural communities by Argentine ants. Oecologia 161:161–171
Savard JPL, Clergeau P, Mennechez G (2000) Biodiversity concepts and urban ecosystems. Landsc Urban Plan 48:131–142
Schlick-Steiner BC, Steiner FM, Moder K, Seifert B, Sanetra M, Dyreson E, Stauffer C, Christian E (2006) A multidisciplinary approach reveals cryptic diversity in Western Palearctic Tetramorium ants (Hymenoptera: Formicidae). Mol Phylogenet Evol 40:259–273
Seto KC, Fragkias M, Güneralp B, Reilly MK (2011) A meta-analysis of global urban land expansion. PLoS One 6:e23777. doi:10.1371/journal.pone.0023777
Seto KC, Güneralp B, Hutyra LR (2012) Global forecasts of urban expansion to 2030 and direct impacts on biodiversity and carbon pools. Proc Natl Acad Sci U S A 109:16083–16088
Smith MR (1946) Ant hosts of the fungus, Laboulbenia formicarum Thaxter. Proc Entomol Soc Wash 48:29–31
Tait CJ, Daniels CB, Hill RS (2005) Changes in species assemblages within the Adelaide metropolitan area, Australia, 1836–2002. Ecol Appl 15:346–359
Vitousek PM, Mooney HA, Lubchenko J, Melillo JM (1997) Human domination of earth’s ecosystems. Science 277:494–499
Ward PS (2000) Broad-scale patterns of diversity in leaf litter ant communities. In: Agosti D, Majer JD, Alonso LE, Schultz TR (eds) Ants: standard methods for measuring and monitoring biodiversity. Smithsonian Institution Press, Washington, pp 99–121
Wetterer JK (2009) Worldwide spread of the Penny ant, Tetramorium bicarinatum (Hymenoptera: Formicidae). Sociobiology 54:811–830
Wheeler WM (1910) Colonies of ants (Lasius neoniger Emery) infested with Laboulbenia formicarum Thaxter. Psyche 17:83–86
Yasuda M, Koike F (2009) The contribution of the bark of isolated trees as habitat for ants in an urban landscape. Landsc Urban Plan 92:276–281
Aknowledgments
The authors would like to thank Neil McCoy, Katherine Driscoll, Matthew Drew for their help in collecting. We would like to thank Terry Nuhn for discussion about his collection. Thank you also to Alex Wild for authorizing the use of his pictures. Finally, we’d like to thank two anonymous reviewers for their comments on a previous version of the manuscript. The work of RRD and BG was supported by a NASA Biodiversity Grant (ROSES-NNX09AK22G), support from a USGS grant to the Climate Science Center and an NSF-CAREER grant (09533390).
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article

Cite this article
Guénard, B., Cardinal-De Casas, A. & Dunn, R.R. High diversity in an urban habitat: are some animal assemblages resilient to long-term anthropogenic change?. Urban Ecosyst 18, 449–463 (2015). https://doi.org/10.1007/s11252-014-0406-8
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11252-014-0406-8