
Abstract
As urbanization transforms landscapes around the globe, we are challenged to find ways for urban habitats to contribute to ongoing wildlife conservation efforts. Yet aside from basic distributions and relative abundance, we know little about the population dynamics of most species occupying urban habitats and almost nothing about the extent to which local populations serve as source or sink habitats. We analyzed the spatial synchrony and persistence of local populations of 16 avian species breeding in 14 mature riparian forests in central Ohio, USA during 2005–2011. In addition, we examined demographic growth rates (λ) for 2 focal species (northern cardinal, Cardinalis cardinalis, and Acadian flycatcher, Empidonax virescens) to understand source-sink structure. Our results showed high rates of population turnover for the majority of species and negative growth rates for both focal species, indicating that most populations functioned as sinks. Local populations showed little evidence of population synchrony, as we found consistently low or negative spatial autocorrelation and relatively high rates of population turnover for both migratory and resident species. Collectively our results suggest that long-term persistence of regional populations of birds breeding in Midwestern riparian forest fragments may be facilitated by metapopulation dynamics.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Spurred by rapid expansion of urban land, a variety of agencies, NGOs, and land trusts have ramped up efforts to protect even small parcels of habitat in an effort to conserve biodiversity within urbanizing landscapes. Few studies, however, have examined the extent to which small populations in such remnants might be demographically viable. Although small populations in fragmented habitats can be vulnerable to extinction (Wilcox and Murphy 1985; Newman and Pilson 1997; Gyllenberg et al. 1993; Fahrig 2002; Hanski and Ovaskainen 2003), the literature shows that in some cases regional populations can persist if structured as a metapopulation where subpopulations fluctuate independently (i.e. not in synchrony) with possibility of recolonization should they become locally extinct (Hanski and Gilpin 1991; Koenig 1998). Indeed metapopulation structure is known to reduce risk of extinction in fragmented landscapes across a wide range of taxa including plants (Kiviniemi and Lofgren 2009), invertebrates (Hanski et al. 1995; Drechsler et al. 2003; Guiney et al. 2010; Harrison et al. 1988), fish (Smedbol and Wroblewski 2002; Figueira 2009), herptiles (Marsh and Trenham 2001; Hels and Nachman 2002; Templeton et al. 2011), mammals (Sweanor et al. 2000; Magle et al. 2010), and birds (Busch et al. 2000; Cattadori et al. 1999; Scheiman et al. 2007; Ringsby et al. 2002). Metapopulations within urbanizing landscapes remain largely unexamined (Niemelä 1999), despite the fact that reproductive rates, population turnover, and landscape permeability are strongly affected by urban land uses (Hanski and Gilpin 1991; Opdam 1991; Soule et al. 1988, Reed and Levine 2005).
Assuming that spatially subdivided populations existing within fragmented urbanizing landscapes function as metapopulations can overestimate persistence and viability of populations. Unfortunately, the empirical evidence required to demonstrate a metapopulation structure, such as dispersal rates among patches, is difficult to attain. Population synchrony is one of the easier methods to obtain metrics that can be used to evaluate the landscape level structure of a population. If dynamics among subpopulations are highly synchronous, the likelihood that an increasing subpopulation will be able to compensate for a decreasing one is significantly reduced (Hanski and Gilpin 1991; Wilson 2005, Munkemuller and Johst 2006). Consequently, asynchronous population dynamics are considered both prerequisites for and indicators of potential metapopulation dynamics (Driscoll 2007).
Measures of spatial autocorrelation, the tendency for ecological variables at nearby points in space and time to have more similar values than would be expected by chance, can be powerful tools for quantifying population synchrony. Spatial autocorrelation has been documented throughout nature in species ranging from large mammals to fungal pathogens (Lichtenstein et al. 2002; Liebhold et al. 2004). Despite the ubiquity of autocorrelation in ecological systems, the underlying mechanisms remain unclear. Three commonly cited mechanisms hypothesized to cause spatial synchrony are: (a) correlated environmental conditions that drive population dynamics [Moran effects; Moran 1953; Ranta et al. 1997], (b) dispersal or movement between populations, and (c) interspecific interactions such as predator/prey relationships (Koenig 2002; Liebhold et al. 2004; Kiviniemi and Lofgren 2009; Rosenstock et al. 2011). Any of these mechanisms may produce a pattern of declining synchrony with increasing distance (Bellamy et al. 2003).
We used patterns of population synchrony and rates of extinction and recolonization to provide insight into the structure of populations and the extent to which they operate as a metapopulation (Fig. 1). There were two scenarios considered to be consistent with metapopulation structure, cases with (1) high negative spatial autocorrelation and high rates of extinction and recolonization, or (2) low spatial autocorrelation coupled with high rates of extinction and recolonization. Within this context, we also evaluated the extent to which synchrony was associated with characteristics of the landscape matrix and the migratory status of species. For two focal species we also examined the extent to which local populations operated as sinks or sources, as well as the synchrony among population growth rates.

Decision tree illustrating how patterns of spatial autocorrelation and rates of extinction & recolonization can be used to provide insight into population structure. Shaded boxes indicate scenarios that are consistent with the theoretical underpinnings of metapopulation dynamics
Methods
Study system and field methodology
We studied bird communities in 14 mature riparian forest fragments located within the greater Columbus metropolitan areas of Franklin and Delaware Counties, Ohio, USA. Fragments were separated by > 2 km and consisted of mature forests that were of comparable size, shape, and configuration across a rural–urban gradient (Rodewald and Shustack 2008a). Common trees and woody understory plants included sycamore (Platanus occidentalis), boxelder (Acer negundo), sugar maple (Acer saccharum), black walnut (Juglans nigra), ash (Fraxinus spp.), Amur honeysuckle (Lonicera maackii), tall paw-paw (Asimina triloba), and Ohio buckeye (Aesculus glabra). Fragments were separated by > 2 km and consisted of mature forests that were of comparable size, shape, and configuration across a rural–urban gradient that had building densities ranging from 10–727 buildings/km2 (0.1–7.3 buildings per ha). A principle components analysis was performed on landscape metrics (e.g., percent impervious surface, number of buildings, etc.) for each site, the first principle component (hereafter referred to as ‘urban index’) accounted for 80 % of the variation within the data set and was positively correlated with increasing urbanization (Rodewald ad Shustack 2008a). Where possible, we treated urbanization as a continuous variable; however, when our analysis required categories urban index values were used to categorize sites as either urban (urban index > 0) or rural (urban index < 0). These two categories of landscapes were qualitatively and quantitatively distinct. Rural landscapes averaged 80 buildings/km2 (10–260/ha range) and were dominated by cropland, pasture, managed grasslands, and farms. Urban landscapes, in contrast, averaged 541 buildings/km2 (391–728/ha range) and were dominated by residential areas, commercial development, and roads.
Study organisms
We studied 16 common bird species breeding on our study sites, that represented two migratory guilds: 8 Neotropical migratory species (Acadian flycatcher – Empidonax virescens, Baltimore oriole – Icterus galbula, blue-gray gnatcatcher – Polioptila caerulea, great-crested flycatcher – Myiarchus crinitus, gray catbird – Dumetella carolinensis, indigo bunting – Passerina cyanea, red-eyed vireo – Vireo olivaceus, and wood thrush – Hylocichla mustelina), and 8 resident species (American robin – Turdus migratorius, brown-headed cowbird – Molothrus ater, Carolina chickadee – Poecile carolinensis, Carolina wren – Thryothorus ludovicianus, downy woodpecker – Picoides pubescens, northern cardinal – Cardinalis cardinalis, red-bellied woodpecker – Melanerpes carolinus, and tufted titmouse – Baeolophus bicolor). In addition to these 16 species, the Acadian flycatcher and northern cardinal were used as focal species for a more intensive demographic examination. Species also represented two very different responses to urbanization (Rodewald and Bakermans 2006). Migrants tended to be strong urban avoiders, such that their numbers declined as landscapes surrounding parks urbanized. In contrast, the residents were urban adaptors and were most numerous in urban landscapes. Of the urban avoiders and urban adapters, the Acadian flycatcher and northern cardinal were used as focal species for a more intensive demographic examination given that they were sufficiently common to achieve adequate sample sizes and they shared open cup nests and understory habitats.
We used previously collected field data generated from standardized surveys of breeding brds within 2-ha grids within each site. Density estimates were calculated from spot-map surveys (Bibby et al. 2000) conducted at each site from May through early July 2005 through 2011. From April – August of these same years we searched for nests and monitored season-long reproductive output of our two focal species, Acadian flycatchers and northern cardinals (see Rodewald et al. 2013 for details). To aid in tracking success of territorial pairs, flycatchers and cardinals were banded with both a USGS leg band (steel for cardinals, aluminum for flycatchers) and a unique combination of three color bands for individual identification. Territories of banded individuals were closely monitored with special care given to locating nests and all fledged young over the course of the season in order to establish an accurate estimate of reproductive productivity.
Data analysis
We quantified the degree of ecological similarity among spatially distinct sites using Mantel’s tests for spatial autocorrelation (Mantel 1967), which tests for similarity between (1) a matrix of geographic distances between locations, and (2) a matrix of ecological “distances” (i.e. differences) between environmental parameters. The null hypothesis of the Mantel test is that values of the environmental parameters are distributed randomly throughout space, whereas, statistical significance indicates that locations in space differ significantly from random, or, that environmental values are not randomly distributed in space (Mantel 1967). The test results in a normalized Mantel statistic, the correlation of all pairwise elements of matrices X and Y, which, like any product–moment coefficient ranges from 1 to −1 (Fortin and Gurevitch 2001). Mantel tests were run with PASSaGE – Pattern Analysis Spatial Statistics and Geographic Exegesis, a program used for spatial analysis in natural sciences (Rosenberg and Anderson 2011). We also ran a modified form of the Mantel test known as the Mantel correlogram which shows the strength of correlation between the two matrices across a range of five distance categories (Cushman et al. 2005).
For each species we created three unique geographic distance matrices, one for all sites (n = 14), one for rural sites (n = 6), and one for urban sites (n = 8). Each being a symmetrical, square n × n matrix of Euclidian straight-line distance. Each i,j component of the matrix was the distance in meters between sites.
For each species PASSaGE was used to create a correlation matrix of demographic fluctuations from log transformed [ln(x + 1)] changes in species density from year to year. The result was a matrix of correlations in year-by-year territory density fluctuations for each i,j site comparison. As with the geographic matrices, we created three autocorrelation matrices for each species, one for all sites (n = 14), one for rural sites (n = 6), and one for urban sites (n = 8).
We also tested for spatial autocorrelation of the population growth rates (lambdas) of Acadian flycatchers and northern cardinals. Lambda values were calculated using previously published survival rates for our study system with the Leslie matrix procedure (Caswell 2001). The Leslie matrix is an age-classified population-modeling matrix in which each column represents an age class of the population. A unique Leslie matrix was constructed for each of my focal species at each site in every year, resulting in a total of 98 Leslie matrices per species. As commonly applied to bird populations, we used a three age-class structure that included post-fledging, juvenile, and adult. We did not include egg or nestling survival (prior to fledging) because rates of nest predation and brood parasitism in our system are relatively high, as a result the number of successfully fledged young is a far better indicator of territory specific fecundity than number of eggs laid. Leslie matrices included previously published demographic parameters, territory specific fertility of the adult reproductive age class, which was calculated from the number of young fledged per territory per year (see Rodewald et al. 2013). Post fledging survival (P) estimates were obtained from a previous radio-telemetry study of fledged birds in our system. Cumulative survivorship during the post-fledging period was 0.44 ± 0.077 for cardinals and 0.72 ± 0.077 for flycatchers (Ausprey 2010; Ausprey and Rodewald 2011). Because post-fledging survival did not vary with urbanization, we used the same estimates for both urban and rural sites. Juvenile survivorship (J), between post fledging and the age of reproduction, was estimated as post fledging survival*0.5, 0.36 for flycatchers, and 0.22 for cardinals, as is commonly used in the literature (Noon and Saur 2001).
Estimates of adult survival were based on mark-recapture (resight) data of banded birds at our study sites that were analyzed using program MARK. Cardinal and flycatcher survivorship was 0.64 ± 0.039 and 0.53 ± 0.056 respectively, and did not differ between urban and rural landscapes (Rodewald and Shustack 2008b, Rodewald and Shustack 2008a). Because fecundity estimates varied by site and year, an individual Leslie matrix was constructed for each site in each year. Lambda was then calculated as the dominant eigenvalue of the Leslie matrix using the program MATLAB. Autocorrelation matrices of year-by-year changes in flycatcher and cardinal lambdas were computed using the same methods described for spot map data.
Using annual spot map data we estimated extinction and recolonization rates for all 16 study species over the 7 year study period. An extinction event was said to have occurred when a patch was occupied in year x and unoccupied in year x + 1. Likewise, a recolonization event was declared when a patch went from unoccupied in year x to occupied in year x + 1. The number of possible extinction/recolonization events was tallied as the total number of occupied (or unoccupied) sites where the density of the subsequent year was also estimated. Finally, the probability of extinction or recolonization was calculated as the number total of extinction/recolonization events divided by the number of possible extinction/recolonization events.
Results
Species densities
Only 3 of 16 species (18.8 %) exhibited significant (P < 0.10) positive spatial autocorrelation across the entire study area (14 sites), whereas seven species (43.7 %) showed negative spatial autocorrelation. Though migrants and residents showed similar degrees of autocorrelation as a whole (migrants r = 0.014 ± 0.0585 vs. residents r = 0.067 ± 0.0475; p = 0.494), 5 of 8 migrant species showed negative spatial autocorrelation compared to only 2 of 8 resident species (Table 1). The mean scale of positive spatial synchrony was 25.5 km for the entire study area and did not differ between migratory guilds.
Spatial autocorrelation in rural sites ranged from r = −0.371 (gray catbird) to r = 0.391 (blue-gray gnatcatcher), with only the gnatcatcher (P = 0.08) showing marginal significance. Autocorrelation in urban sites ranged from r = −0.206 (Baltimore oriole) to r = 0.471 (Acadian flycatcher), with Acadian flycatchers (P = 0.03) and Carolina wrens (P = 0.07) showing positive significant autocorrelation. Mean autocorrelation of migrant and resident birds did not differ from each other in rural sites (migrants r = −0.094 ± 0.099 vs residents r = 0.060 ± 0.0611; p = 0.211) or urban sites (migrants r = 0.053 ± 0.0792 vs. residents r = −0.0238 ± 0.051; p = 0.429).
Guild densities
Migrants
Density of migrants was negatively autocorrelated for the study area (r = −0.117; left-tailed P = 0.287) and rural sites (r = −0.443; left-tailed P = 0.027), but showed positive spatial autocorrelation in urban sites (r = 0.129; P = 0.154). Mantel correlograms showed that the degree of autocorrelation varied among distance classes. Autocorrelation differed between rural and urban sites at the smallest spatial scale (rural = −0.341 vs. urban =0.084: P = 0.023), but not at larger scales (rural = 0.0124 vs. urban = 0.0162; P = 0.935).
Residents
Residents showed weakly negative autocorrelation in both rural (r = −0.095; P = 0.549) and urban sites (r = −0.05131; P = 0.0569), while the entire landscape yielded positive and significant spatial autocorrelation (r = 0.312; P = 0.038). Like migrants, autocorrelation of residents differed between rural and urban landscapes at the smallest spatial scale (p = 0.007). Autocorrelation also differed between migrant and resident birds at the smallest distance category in both rural (p = 0.017) and urban (p = 0.036) landscapes.
Extinction and recolonization probability
Over 7 years of spot mapping territories (2005–2011) we observed 109 local extinction events and 123 colonization events. The northern cardinal lacked any extinction or colonization events over the course of the study, while the Carolina wren had the greatest number of extinction (22) and colonization (19) events of any species. The Baltimore oriole had the highest probability of extinction over the course of the study (0.789), and was also the only species with a probability of extinction higher than probability of recolonization (0.373).
Probability of extinction ranged from 0.0–0.789 with mean 0.166 ± 0.0522; probability of recolonization ranged from 0.0–1.0 with mean 0.652 ± 0.0815. Probability of extinction (migrants = 0.2283; residents = 0.1027, p = 0.2457) and recolonization (migrants = 0.5747; residents = 0.7293, p = 0.3613) was similar among migratory guilds.
Demography
Mean population growth rate of flycatchers for each landscape was; All = 0.788 ± 0.027; Rural = 0.853 ± 0.038; Urban = 0.7387 ± 0.036. Mean population growth rates for cardinals were; All = 0.868 ± 0.011; Rural = 0.866 ± 0.021; Urban = 0.871 ± 0.012. Spatial autocorrelation for the Acadian flycatcher was landscape-dependent, with subpopulations being negatively autocorrelated in rural landscapes (r = −0.212; p = 0.862), but significantly positively autocorrelated in urban landscapes (r = 0.511, p = 0.035). Autocorrelation across the entire landscape approached zero (r = 0.08102; p = 0.25). Spatial autocorrelation of northern cardinals was similarly negative across the study area as a whole (r = −0.148) and within rural (r = −0.21776) and urban sites (r = −0.237).
Discussion
Despite growing efforts to protect natural areas within urbanizing landscapes, few have evaluated the extent to which population dynamics within and among habitat patches might allow for regional population persistence. We found weak synchrony (i.e., positive spatial autocorrelation) of population densities and demographic rates of flycatchers and cardinals coupled with high rates of population turnover, which provides some of the first evidence that local avian subpopulations may function as metapopulations. Metapopulation structure has been suspected in many avian species on the basis of patchy distributions, without additional supporting data on population structure (Lahaye et al. 1994; Donovan et al. 1995a; Ringsby et al. 2002; Ainley et al. 2003; Schippers et al. 2011). Our work is distinct from previous studies in that we examined population turnover as well as population synchrony for both densities and demography.
Among the most striking patterns in our system were high rates of local population turnover, which are foundational elements of the metapopulation concept (Hanski and Gilpin 1991; Gyllenberg et al. 1993; Hanski 1999). With the sole exception of northern cardinals, every species examined experienced at least one local extinction and colonization event. Extinction rates for bird populations in fragmented systems are quite variable depending on factors such as habitat patch size and quality, and population size within the patch (Bellamy et al. 1996; Boulinier et al. 2001; Doherty et al. 2003; Huste and Boulinier 2007). Unlike others reporting that extinction rates in fragmented or disturbed landscapes can be as much as two times greater than rates of recolonization (Crooks et al. 2000) we found that each subpopulation had a 16.6 % chance of extinction with nearly 70 % of extirpated sites being recolonized during the course of our research.
Associations between population dynamics and urbanization also differed between migratory guilds, which is consistent with reports of contrasting population responses of many migratory and resident birds to urban development (Miller et al. 2003; Dunford and Freemark 2005; Stratford and Robinson 2005; Rodewald and Bakermans 2006; Minor and Urban 2010). Populations of Nearctic-neotropical migrant birds in our system were twice as likely to experience local extinction as were resident species perhaps due to their sensitivity to urbanization. Huste and Boulinier (2007) described a similar pattern of extinction for migrant and resident birds in Paris, France, showing that rates of extinction for migratory birds were significantly higher (residents extinction rate = 0.03 ± 0.01 vs. migrants = 0.20 ± 0.03) than those of resident birds.
Among migratory species, low levels of positive spatial autocorrelation, combined with high rates of local extinction and recolonization are consistent with the assumptions of metapopulation theory (Harrison and Quinn 1989; Hanski and Gilpin 1991). Although the two most distant sites in our study system, North Galena and Public Hunting, are only 61 km apart, well within the maximum range of dispersal for most bird species (Hosner and Winkler 2007), the heavily disturbed urban matrix may functionally isolate sites for birds that are sensitive to urbanization (Kennedy et al. 2010; Tremblay and St Clair 2011). Sensitive bird species may be hesitant to move through a heavily urbanized landscape matrix due to factors such as low levels canopy cover in the matrix (Tremblay and St Clair 2011), greater density of predator species (Rodewald and Kearns 2011), and higher mortality due to building collisions (Chace and Walsh 2006). Tremblay and St Clair (2011) showed that urban sensitive bird species required 20–40 % canopy cover for movement between sites, while urban adaptive birds moved through landscapes with as little as 2–4 % cover. More moderate levels of spatial autocorrelation among populations of resident bird species may reflect their readiness to move through and breed within the urban/suburban landscape matrix in our system (Malpass and Rodewald unpublished data).
Demographic data for cardinals and flycatchers highlight the importance of considering habitat type and life history characteristics in source-sink metapopulation dynamics, as most populations appeared to be in decline (λ < 1). Patterns of spatial autocorrelation suggest that the potential for rescue events differed between the two species. High population synchrony (i.e. positive spatial autocorrelation) among sinking flycatcher populations (λ < 1) in urban landscapes provides little hope that a collection of remnants within cities will persist in the long term, whereas source-sink dynamics may operate less disturbed landscapes with higher average growth rates. Source-sink dynamics have been documented in woodlots fragmented by an agricultural matrix (Donovan et al. 1995b; Robinson et al. 1995; Brawn and Robinson 1996; Foppen et al. 2000; Hoover et al. 2006), but fewer studies have examined source-sink dynamics in urban systems. Our research suggests that not only fragmentation, but also land cover (i.e. urbanization) can mediate source – sink dynamics.
Although high density of individuals might suggest that urban sites improve reproductive output of cardinals, mean lambda was below zero (negative growth rate) for every site, with only 8/112 calculated lambdas being >1 (positive growth rate). Theoretical and experimental work has shown that populations comprised entirely of long-term sinks may persist given temporal and spatial variation (asynchrony) in habitat quality and reproductive success (Matthews and Gonzalez 2007). Growth rates of cardinals were negatively autocorrelated in all landscape types, and therefore may persist in spite of regionally poor reproductive success.
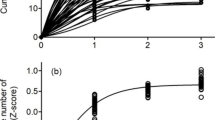
As one of few studies to use both spatial synchrony and population turnover rates, we provide evidence of metapopulation dynamics in bird communities in an urbanizing landscape. That said, we acknowledge some important limitations of our work. For example, we were unable to census all available habitat within the landscape, owing partly to logistical constraints and partly due to limited access to private property. This means that our estimates of occupancy are likely underestimated, which could lead to somewhat inflated rates of population turnover. In addition, like most other studies of highly vagile organisms, we were unable to directly observe dispersal events, which limits our understanding of the true connectivity (or independence) of subpopulations. However, in spite of these limitations, we were still encouraged by the frequency of local rescue events (recolonizations) that we observed, which suggests that even small populations might persist over the long term. In this way our study underscores the value of protecting known source habitats in urbanizing landscapes to ensure that nearby habitat remnants will retain some conservation value (Figs. 2 and 3).

Mantel correlograms of migrant and resident birds in rural and urban landscapes respectively (2005–2011). Autocorrelation of migratory guilds responded differently to landscape type. Autocorrelation of migrants and residents differed significantly from each other in both urban (p = 0.036), and rural (p = 0.017) sites

Growth rates of Acadian flycatchers and northern cardinals across the urban index. A more positive urban index indicates a more urban site, while negative urban index values indicate more rural sites. Flycatchers show a strong decreasing trend in growth rate as urbanization increases (r = −0.34), while reproductive success of cardinals remains unchanged with urbanization (r = 0.04)
References
Ainley DG, Ballard G, Barton KJ, Karl BJ, Rau GH, Ribic CA, Wilson PR (2003) Spatial and temporal variation of diet within a presumed metapopulation of adelie penguins. Condor 105(1):95–106
Ausprey IJ (2010) Post-fledging ecology of two songbird species across a rural-to-urban landscape gradient. Masters Thesis. The Ohio State University
Ausprey IJ, Rodewald AD (2011) Postfledging survivorship and habitat selection across a rural-to-urban landscape gradient. Auk 128(2):293–302
Bellamy P, Hinsley S, Newton I (1996) Local extinctions and recolonisations of passerine bird populations in small woods. Oecologia 108(1):64–71
Bellamy PE, Rothery P, Hinsley SA (2003) Synchrony of woodland bird populations: the effect of landscape structure. Ecography 26:338–348
Bibby CJ, Burgess NP, Hill DA, Mustoe S (2000) In bird census techniques, 2nd edn. Academic, London
Boulinier T, Nichols JD, Hines JE, Saur JR, Flather CH, Pollock KH (2001) Forest fragmentation and bird community dynamics: inference at regional scales. Ecology 82(4):1159–1169
Brawn JD, Robinson SK (1996) Source-sink population dynamics may complicate the interpretation of long-term census data. Ecology 77(1):3–12
Busch JD, Miller MP, Paxton EH, Sogge MK, Keim P (2000) Genetic variation in the endangered southwestern willow flycatcher. Auk 117(3):586–595
Caswell H (2001) Matrix population models: construction, analysis, and interpretation. 2nd edition. Sinaur Associates, Inc
Cattadori IM, Hudson PJ, Merler S, Rizzoli A (1999) Synchrony, scale and temporal dynamics of rock partridge (Alectoris graeca saxatilis) populations in the Dolomites. J Anim Ecol 68:540–549
Chace, J. F., & Walsh, J. J. (2006) Urban effects on native avifauna: A review. Landscape and Urban Planning, 74(1):46–69.
Cushman SA, Chase M, Griffin C (2005) Elephants in space and time. Oikos 109:331–341
Doherty P, Sorci G, Royle J, Hines J, Nichols J, Boulinier T (2003) Sexual selection affects local extinction and turnover in bird communities. Proc Natl Acad Sci 100(10):5858–5862
Donovan TM, Lamberson RH, Kimber A, Thompson FR, Faaborg J (1995a) Modeling the effects of habitat fragmentation on source and sink demography of neotropical migrant birds. Conserv Biol 9(6):1396–1407
Donovan T, Thompson F, Faaborg J, Probst J (1995b) Reproductive success of migratory birds in habitat sources and sinks. Conserv Biol 9(6):1380–1395
Drechsler M, Frank K, Hanski I, O’Hara RB, Wissel C (2003) Ranking metapopulation extinction risk: from patterns in data to conservation management decisions. Ecol Appl 13(4):990–998
Driscoll DA (2007) How to find a metapopulation. Can J Zool 85:1031–1048
Dunford W, Freemark K (2005) Matrix matters: effects of surrounding land uses on forest birds near Ottawa, Canada. Landsc Ecol 20(5):497–511
Fahrig L (2002) Effect of habitat fragmentation on the extinction threshold: a synthesis. Ecol Appl 12(2):346–353
Figueira WF (2009) Connectivity or demography: defining sources and sinks in coral reef fish metapopulations. Ecol Model 220(8):1126–1137
Foppen RPB, Chardon LP, Lifeveld W (2000) Understanding the role of sink patches in source-sink metapopulations: reed Warbler in an agricultural landscape. Conserv Biol 14(6):1881–1892
Fortin MJ, Gurevitch J (2001) Mantel tests: spatial structure in field experiments. In: Scheiner SM, Gurevitch J (eds) Design and analysis of ecological experitments, 2nd edn. Oxfard University Press, UK, pp 308–326
Guiney MS, Andow DA, Wilder TT (2010) Metapopulation structure and dynamics of an endangered butterfly. Basic Appl Ecol 11:354–362
Gyllenberg M, Söderbacka G, Ericsson S (1993) Does migration stabilize local population dynamics? Analysis of a discrete metapopulation model. Math Biosci 118(1):25–49
Hanski I (1999) Metapopulation ecology. Oxford University Press, London
Hanski I, Gilpin M (1991) Metapopulation dynamics - brief-history and conceptual domain. Biol J Linn Soc 42(1–2):3–16
Hanski I, Ovaskainen O (2003) Metapopulation theory for fragmented landscapes. Theor Popul Biol 64(1):119–127
Hanski I, Pakkala T, Kuussaari M, Guangchun L (1995) Metapopulation persistence of an endangered butterfly in a fragmented landscape. Oikos 72(1):21–28
Harrison S, Quinn JF (1989) Correlated environments and the persistence of metapopulations. Oikos 56(3):293–298
Harrison S, Murphy DD, Ehrlich PR (1988) Distribution of the bay checkerspot butterfly, Euphydryas editha bayensis: evidence for a metapopulation model. Am Nat 132(3):360–382
Hels T, Nachman G (2002) Simulating viability of a spadefoot toad Pelobates fuscus metapopulation in a landscape fragmented by roads. Ecography 25:730–744
Hoover JP, Tear TH, Baltz ME (2006) Edge effects reduce the nesting success of Acadian flycatchers in a moderately fragmented forest. J Field Ornithol 77(4):425–436
Hosner PA, Winkler DW (2007) Dispersal distances of tree swallows estimated from continent-wide and limited-area data. J Field Ornithol 78(3):290–297
Huste A, Boulinier T (2007) Determinants of local extinction and turnover rates in urban bird communities. Ecol Appl 17(1):168–180
Kennedy CM, Marra PP, Fagan WF, Neel MC (2010) Landscape matrix and species traits mediate responses of Neotropical resident birds to forest fragmentation in Jamaica. Ecol Monogr 80(4):651–669
Kiviniemi K, Lofgren A (2009) Spatial (a)synchrony in population fluctuations of five plant species in fragmented habitat. Basic Appl Ecol 10:70–78
Koenig WD (1998) Spatial autocorrelation in California land birds. Conserv Biol 12(3):612–660
Koenig WD (2002) Global patterns of environmental synchrony and the Moran effect. Ecography 25:283–288
Lahaye W, Gutierrez R, Akcakaya H (1994) Spotted owl metapopulation dynamics in southern California rid. J Anim Ecol 63(4):775–785
Lichtenstein JW, Simons TR, Franzreb KE (2002) Landscape effects on breeding songbird abundance in managed forests. Ecol Appl 12:836–857
Liebhold A, Koenig WD, Bjornstad ON (2004) Spatial synchrony in population dynamics. Annu Rev Ecol Evol Syst 35:467–490
Magle SB, Reyes P, Zhu J, Crooks KR (2010) Extirpation, colonization, and habitat dynamics of a keystone species along an urban gradient. Biol Conserv 143:2146–2155
Mantel N (1967) The detection of disease clustering and a generalized regression approach. Cancer Res 27(1):209–220
Marsh DM, Trenham PC (2001) Metapopulation dynamics and amphibian conservation. Conserv Biol 15(1):40–49
Matthews DP, Gonzalez A (2007) The inflationary effects of environmental fluctuations ensure the persistence of metapopulations. Ecology 88(11):2848–2856
Miller J, Wiens J, Hobbs N, Theobald D (2003) Effects of human settlement on bird communities in lowland riparian areas of Colorado (USA). Ecol Appl 13(4):1041–1059
Minor E, Urban D (2010) Forest bird communities across a gradient of urban development. Urban Ecosyst 13:51–71
Moran PAP (1953) The statistical analysis of the Canadian lynx cycle. II. Syncronization and meteorology. Aust J Zool 1:291–298
Newman D, Pilson D (1997) Increased probability of extinction due to decreased genetic effective population size: Experimental populations of Clarkia pulchella. Evolution 51(2):354–362
Niemelä J (1999) Is there a need for a theory of urban ecology? Urban Ecosyst 3(1):57–65
Noon B, Saur J (2001) Population models for passerine birds: structure, parameterization, and analysis. In: McCullogh DR, Barrett RH (eds) Wildlife 2001: populations. Elsevier Applied Science, New York
Opdam P (1991) Metapopulation theory and habitat fragmentation: a review of holarctic breeding bird studies. Landsc Ecol 5(2):93–106
Ranta E, Kaitala V, Lindstrom J, Helle E (1997) The Moran effect and synchrony in population dynamics. Nord Soc Oikos 78(1):136–142
Ringsby T, Saether B, Tufto J, Jensen H, Solberg E (2002) Asynchronous spatiotemporal demography of a house sparrow metapopulation in a correlated environment. Ecology 83(2):561–569
Robinson SK, Thompson FR, Donovan TM, Whitehead DR, Faaborg J (1995) Regional forest fragmentation and the nesting success of migratory birds. Science 267(5206):1987–1990
Rodewald AD, Bakermans MH (2006) What is the appropriate paradigm for riparian forest conservation. Biol Conserv 128(2):193–200
Rodewald, A.D., & Kearns, L.J. (2011) Shifts in dominant nest predators along a rural-to-urban landscape gradient. The Condor, 113(4):899–906.
Rodewald AD, Shustach DP (2008) Consumer resource matching in urbanizing landscapes: are synanthropic birds over-matching. Ecology 89(2):515–521
Rodewald AD, Shustack DP (2008) Urban flight: understanding individual and population level responses of Nearctic-Neotropical migratory birds to urbanization. J Anim Ecol 77:83–91
Rosenberg MS, Anderson CD (2011) PASSaGE: pattern analysis, spatial statistics and geographic exegesis. Version 2. Methods Ecol Evol 2:229–232
Rosenstock TS, Hastings A, Koenig WD, Lyles DJ, Brown PB (2011) Testing Moran’s theorem in an agroecosystem. Oikos 120:1434–1440
Scheiman DM, Dunning JB, With KA (2007) Metapopulation dynamics of Bobolinks occupying agricultural grasslands in the Midwestern United States. Am Midl Nat 158:415–423
Schippers P, Verboom J, Vos CC, Jochem R (2011) Metapopulation shift and survival of woodland birds under climate change: will species be able to track. Ecography 34:909–919
Smedbol RK, Wroblewski JS (2002) Metapopulation theory and northern cod population structure: interdependency of subpopulations in recovery of a groundfish population. Fish Res 55:161–174
Soule ME, Bolger DT, Allison CA, Wright J, Sorice M, Hill S (1988) Reconstructed dynamics of rapid extintions of chaparral-requiring birds in urban habitat islands. Conserv Biol 2(1):75–92
Stratford JA, Robinson WD (2005) Distribution of neotropical migratory bird species across an urbanizing landscape. Urban Ecosyst 8(1):59–77
Sweanor LL, Logan KA, Hornocker MG (2000) Cougar dispersal patterns, metapopulation dynamics, and conservation. Conserv Biol 14(3):798–808
Templeton AR, Brazeal H, Neuwald JL (2011) The transition from isolated patches to a metapopulation in the eastern collared lizard in response to prescribed fires. Ecology 92(9):1736–1747
Tremblay MA, St Clair CC (2011) Permeability of a heterogeneous urban landscape to the movements of forest songbirds. J Appl Ecol 48(3):679–688
Wilcox BA, Murphy DD (1985) Conservation strategy: the effects of fragmentation on extinction. Am Nat 125(6):879–887
Wilson TE (2005) Spatial synchrony in terrestrial breeding birds: implications for metapopulation processes. Thesis (M.S.) The Ohio State University
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article

Cite this article
Padilla, B.J., Rodewald, A.D. Avian metapopulation dynamics in a fragmented urbanizing landscape. Urban Ecosyst 18, 239–250 (2015). https://doi.org/10.1007/s11252-014-0390-z
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11252-014-0390-z