
Abstract
Anurans and birds rely on sound for a number of social behaviors. Species that use roadside habitats are exposed to traffic noise that can mask important social signals and directly affect the community diversity and composition. We evaluate the impact of traffic noise on anuran and bird species richness, species occurrence, and composition in Puerto Rico, where there is a high density of highways and cars that generate high levels of noise pollution. We compared paired forest sites near (100 m, n = 20, dB > 60) and far (>300 m, n = 20, dB < 60) from highways, with similar vegetation structure, but different levels of noise. We found that the anuran community was not affected by traffic noise. In contrast, bird species richness and occurrence were significantly lower in sites near the highway, and bird species composition also varied significantly. Bird species with low-frequency songs were only detected in sites far from highways. The differences in the ecology and communication behavior between anurans and birds could explain these results. Anurans mainly call at night, when traffic activity was low. In contrast, bird singing activity occurs during the day and overlaps with the high levels of traffic noise. In addition, in natural habitats, Puerto Rican anurans occur at high densities and form noisy choruses (>80 dB), which may allow them to tolerate high levels of anthropogenic noise.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Many animal species use acoustic communication for a wide range of essential functions, including territorial defense and mating, but also for navigation, nurturing, detection of predators, and foraging. Sound is a very effective way to communicate over relatively long-distances; however, acoustic signals can be masked or interfered by various sources of noise (Amézquita et al. 2006; Boeckle et al. 2009; Brumm 2004; Brumm and Slabbekoorn 2005). Natural sources of background noise include wind, rain, and waterfalls can generate continuous background noise at low frequencies (e.g. under 4 kHz) (Brumm and Slabbekoorn 2005; Slabbekoorn 2004). In addition, arthropods, birds and frogs can produce high levels of background noise in a wide frequency range (0.1–10 kHz) (Gerhardt and Huber 2002).
Human activities such as industry, construction, and transportation are another important source of noise that occurs mainly at low frequencies (<5 kHz). Traffic is considered the most extensive source of anthropogenic noise, particularly in urban areas (Barber et al. 2010; Slabbekoorn and Ripmeester 2008; Sun and Narins 2005; Warren et al. 2006). The transformation of natural habitat into urban areas not only reduces suitable habitat, but also increases noise levels (Slabbekoorn and Ripmeester 2008; Warren et al. 2006), particularly for individuals that use habitats near highways where traffic noise can interfere or mask important social signals.
Studies have shown that animals can respond in different ways to minimize the effect of anthropogenic noise, including altering the amplitude, frequency, timing, and duration of signals to minimize acoustic competition (e.g. Bermúdez-Cuamatzin et al. 2009; Brumm et al. 2009; Parris and Schneider 2008; Slabbekoorn and Peet 2003; Wood and Yezerinac 2006). Some species modify their calling rate, usually increasing the calling rate (Brumm and Slater 2006; Kaiser and Hammers 2009; Sun and Narins 2005), and others may shift the time of calling to early in the morning before traffic begins (Bergen and Abs 1997).
Although some species are able to “adapt” or alter their vocalizations, it is likely that these changes have costs that could affect their survival and reproductive success (Parris et al. 2009; Warren et al. 2006). For example, the increase in urban noise reduced the distance that a robin’s song could propagate through the environment and presumably resulted in attracting fewer mates (Hoelzel 1986). In addition, traffic noise reduced the ability of tree frog females (Hyla chrysoscelis) to detect male advertisement calls (Bee and Swanson 2007). Furthermore, traffic noise can interfere with bird communication during incubation and fledgling phases of reproduction (Forman and Deblinger 2000; Slabbekoorn and Ripmeester 2008).
Our knowledge of noise as an ecological disturbance is limited, but as urban areas expand worldwide the average and peak intensity of noise is increasing, mainly in highly developed regions (Barber et al. 2010; Berglund and Lindvall 1995; Fuller et al. 2007; Slabbekoorn and Ripmeester 2008). So far our knowledge of the influence of traffic noise is limited to individual species (e.g. song frequency and duration), however traffic noise may impact the dynamics of an entire community (Benitez-López et al. 2010; Francis et al. 2009; Peris and Pescador 2004; Rheindt 2003; Stone 2000; Sun and Narins 2005). To better understand the effects of anthropogenic noise at the community level, we compared anuran and bird species richness, species occurrence, and community composition in areas near and far away from major highways in Puerto Rico
Materials and methods
Study areas
The study was conducted in the San Juan - Caguas metropolitan area of northeastern Puerto Rico (18°26′45.04″–18°14′47.29″N and 66°28′28.65″–66°01′35.74″W). The area is located in the subtropical moist forest life zone (Ewel and Whitmore 1973), with annual average rainfall of 1,400 mm and 27.4°C of temperature (NOAA 2010). Puerto Rico is part of the Caribbean biodiversity hot spot (Myers et al. 2000), and has many endemic species of anurans (14 of 25) and birds (17 of 335) (Raffaele 1989; Rivero 1998).
In addition, Puerto Rico has a very high density of highways and high levels of noise pollution. In Puerto Rico the number of cars has increased from 1 million in 1980 to 2.8 millions in 2005 (DTOP 2009), and noise level in the metropolitan areas are consistently above the maximum recommended levels established by the government (day:60 dB; night:50 dB; Fig. 1) (Alicea-Pou et al. 2006).

Noise level in San Juan Metropolitan area, measured in 57 urban stations by the Junta de Calidad Ambiental de Puerto Rico (Alicea-Pou et al. 2006). The solid black line represents the maximum permitted noise level, and the open circles represent the mean of the actual noise level (Leq). (This figure was modify and used with the authors permission)
Sites were located in two habitat types: karst forest (KF) and lowland forest (LF). These represent the most common habitats that are associated with highways and high levels of traffic in metropolitan area (annual average daily traffic >100,000) (DTOP 2009). The highways selected were PR-2 and PR-22 in karst forest and PR-1 and PR-52 in lowland forest. These habitat types have different abiotic characteristics that directly influence the forest structure and tree species composition, and these differences are reflected in differences in a composition of the bird community (Acevedo and Restrepo 2008; Wunderle et al. 1987). In contrast amphibians are mainly generalists and no difference in species composition has been reported between these forest types. Furthermore most species are terrestrial, and have direct development.
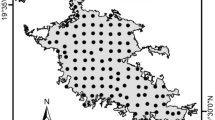
We selected 10 large forest areas (>5 ha, >500 m long and >100 m wide) within each habitat type, adjacent to major highways (Fig. 2). The level of anthropogenic noise in each site was measured using a Digital Sound Level Meter (RadioShack Model No 33-2055 A) set for A-weighting. In each site two areas were sampled, one area near the highway (<100 m), with high levels of anthropogenic noise (>60 dB); and another site at least 400 m from the highway with low levels of anthropogenic noise (<60 dB). Measurements were made between 8:00 and 11:00 am. To estimate the average noise in each site, we took one measurement every 10 s for 1 min (Fig. 3). The anthropogenic noise threshold (60 dB) was based on Dooling and Popper (2007) who studied the negative influence of noise on birds.

Location of study sites in karst forest (KF) and lowland forest (LF) in the San Juan - Caguas metropolitan area in Puerto Rico. Land cover classes were extracted from Gould et al. 2007

Noise level (dB) near and far from the highway in karst forest and lowland forest in Puerto Rico. (Paired t-test, KF: T = 8.05, p < 0.00; LF: T = 11.5, p < 0.00). Box plots illustrate the median (horizontal line within the box), 25–75th percentiles (the box), 10–90th percentiles (T-bar) and the values greater than the 10–90th percentiles (the points)
In addition to the influence of noise, other factors can limit a species distribution in areas around highways, such as habitat loss and fragmentation as well as changes in vegetation structure and complexity (Eigenbrod et al. 2008; Rheindt 2003). Given the potential of these factors to influence the animal communities near and far from roads, we characterized the habitat of each site in a 5 × 50 m plot. In the plot we identified all vegetation >10 cm diameter at breast height (dbh). Additionally, we measured woody plants >1 cm of dbh in a 1 × 50 m transect. In this transect, we also located 10, 1 × 1 m plots, every 5 m. In each of these plots, we determined percent of canopy cover and percent of ground cover (leaf litter, vegetation, soil, rocks). We estimated the canopy height and the presence of foliage in eight layers (0–2 m, 2–4 m, 4–6 m, 6–8 m, 8–10, 10–15 m, 15–20 m) and we also estimated the understory vegetation cover used contact point at two layers (0–1 m, 1–2 m). In addition we measured the depth of leaf litter and soil pH. Google Earth was used to measure the distance to the nearest edge for each site.
Sampling
To quantify the community composition of birds and amphibians we conducted acoustical surveys using Automated Digital Recording System (ADRS), developed by our research group Automated Remote Biodiversity Monitoring Network (ARBIMON) (For additional information see: www.arbimon.net). We used a total of 10 recording devices. Automated digital recorders allow the simultaneous survey of multiple sites, while obtaining data for extended periods of time, thus increasing the probability of detecting a given species (Acevedo and Villanueva-Rivera 2006; Dorcas et al. 2009).
In each site, we simultaneously recorded at the two points, near and far from the highway between May and September 2009. Each recording device was programmed to record 1 min every 20 min for three consecutive days for a total of 216 recordings per site. With this sample scheme, we generated a total of 8,640 recordings \( \left( {{\hbox{two}}\,{\hbox{habitats}} \times {1}0\,{\hbox{sites}}/{\hbox{habitat}} \times {\hbox{two}}\,{\hbox{points}}/{\hbox{sites}} \times {\hbox{three}}\,{\hbox{days}} \times {72}\,{\hbox{recordings}}/{\hbox{day}}} \right) \). All recordings have been archived at www.arbimon.net
Analysis
A Mann Whitney U test was used to compare habitat variables between sites near and far from the road for each habitat type. We analyze recording data from 2 days from each study site (n = 40). We listened to one recording per hour and a total of 1,920 recordings were analyzed for species presence or absence. All analyses were done separately for each habitat type. Species richness was defined as the total species per habitat and site (near and far from the highway). Species occurrence was calculated as the number of sites where a species was detected within each habitat type (n = 10). A paired t-test was used to compare the species richness and species occurrence between sites near and far from the highway. We used a multiple regression analysis to determine the relationship between species richness and noise level (dB), distance to nearest edge, % of canopy cover, canopy height, number of layers of leaf litter and pH. The regression was run separately for amphibians and birds.
For the ordination of the community composition each habitat was analyzed separately using Non metric Multidimensional Scaling Analysis (NMS). Only the bird community was used for this analysis, because the anuran community composition was very similar; all sites shared four of six amphibian species. The NMS analyses compare the dissimilarity or distance between sites based on species composition (McCune and Grace 2002). For the NMS analyses, we used Sorensen (Bray-Curtis) distance, a random starting configuration with maximum of six axes, 50 permutations with real data, Monte Carlo test based on 50 permutations, and stability criterion = 5 × 10−5.
To determine if there were differences in species composition between sites near and far from the highway, we used a Multi-Response Permutation Procedure (MRPP). This nonparametric test is based on analysis of the distance matrix and it evaluates the null hypothesis of no difference in species composition between groups using a randomization procedure (McCune and Grace 2002; Zimmerman et al. 1985). All multivariate analyses were made using the software PC-ORD 5.
Results
There were no significant differences for any habitat variables between sites near and far from the road in both forest types (Appendix 1). Noise level (KF: U = 00.0, P < 0.001; LF: U = 00.0, P < 0.001) and distance to nearest edge (KF: U = 89.0, P = 0.003; LF: U = 92.0, P = 0.001), varied significantly between sites near and far from the highway.
Species richness and species occurrence
In the karst forest sites (KF), five species of anuran (including two endemic species) and 13 species of birds (including six endemic species) were detected. In the lowland forest sites (LF), six species of anurans (including two endemic species) and 17 species of birds were detected (including seven endemic species) (Table 1). Anuran species richness was similar between near and far sites in both habitat types (KF:T = −0.55, p = 0.59; LF:T = −0.70, p = 0.49) (Fig. 4a), but bird species richness was higher in the sites far from highway (KF:T = −3.08, p = 0.013; LF:T = −7.75, p < 0.001) (Fig. 4b). In addition the multiple regression analyses, showed that amphibian species richness was not affected by noise, distance to edge or the habitat variables, but the analysis for birds species richness showed a significant negative relationship with noise level (dB) (Table 2).

Species richness near and far from the highway in karst forest and lowland forest in Puerto Rico. a Amphibians and b birds
The species occurrence of anuran species was similar between sites near and far from the highway in both habitat types (p > 0.05) (Table 1, Fig. 5a). Three amphibian species (Eleutherodactylus antillensis, E. coqui and E. cochranae) occurred in all sites, and they had high species occurrence (above 0.6). E. brittoni was present in all sites, but it occurred at lower species occurrence. Leptodactylus albilabris and Lithobates grylio have low frequency calls (<3 kHz), and they were detected only in sites far from the highway (Table 1).

Differences in species occurrence between sites near and far from the highway in karst forest and lowland forest in Puerto Rico. a Amphibians and b birds. Relative frequency was calculated for each species as the number of sites where a species was detected within each habitat type (n = 10). Negative values represent a higher species occurrence in sites far from the highway
In the two habitats types bird species occurrence was significantly different between sites near and far from the highway (KF: T = −5.84, p < 0.00; LF:T = −6.35, p < 0.001) (Table 1, Fig. 5b). Sites far from the highway had more species with higher species occurrence than sites near the highway. Coereba flaveola was the most frequent bird species in all sites. The second most common species was Vireo altiloquus in karst forest sites and Tyrannus dominiscensis in lowland forest sites.
In karst forests three species (Patagioenas squamosa, Megascops nudipes and Myarchus antillarum) were detected exclusively in sites far from the highway. In lowland forest sites seven species (Crotophaga ani, Melanerpes portoricensis, Todus mexicanus, Vireo latimeri, P. squamosa, M.nudipes and M. antillarum) were identified exclusively in sites far from the highway (Table 1). Five of the species only detected in sites far from the highway are endemic to Puerto Rico (M. nudipes, M. portoricensis, M. antillarum T. mexicanus and V. latimeri).
Bird community composition
Bird community composition showed significant differences between near and far sites in both habitats (Fig. 6). The first axis explained 44% of the variance in karst forest sites and the second axis explained 36%. Multi-Response analysis (MRPP) showed that sites at different distance from highway (near and far) were significantly different (A = 0.044, p = 0.049). The presence of Coccyzus veilloti, M. nudipes, and P. squamosa were important in separating sites. In the lowland forest sites, the first axis explained 55.1% of the variance in the community distance matrix and the second axis explained 18.9%. Multi-Response analysis showed that sites at different distance from highway (near and far) were significantly different (A = 0.088, p = 0.0039). Six species (C. ani, M. nudipes, M. portoricensis, M. antillarum, T. mexicanus, and V. latimeri) were important in separating sites at different distances. The species that we only detected in sites far from the highway have vocalizations below 6 kHz and diurnal vocal activity, with the exception of the Puerto Rican Screech-Owl (M. nudipes), which calls at night.

NMS ordination for bird community composition in karst forest and lowland forests in Puerto Rico. The species listed were the most important separating sites along axis 1. Circles represent sites near the highway and triangle represent sites far from the highway
Discussion
In both the karst and moist forest bird community composition was affected by traffic noise, but amphibian community composition was not. In sites near the road, bird species richness and bird species occurrence were lower than in sites far from the road, and the community composition was also different. The differences in response to anthropogenic noise between anurans and birds are probably related to differences in their ecology and communication behavior.
Anuran species in Puerto Rico are nocturnal and although they can call during the day, the majority of their activity begins after dusk (18–19 h), and there is a little overlap with traffic noise. In contrast, most of the bird singing activity is during the daylight hours (6–18 h), and this overlaps with high levels of traffic noise in the metropolitan area (Fig. 1). The combination of high levels of traffic noise overlapping with the period of calling activity is making birds more vulnerable to the masking effect of traffic noise (Slabbekoorn and Ripmeester 2008). However, if background noise masks a bird’s song, they can move to other sites with a lower levels of noise (Reijnen et al. 1995; Slabbekoorn and Ripmeester 2008), and this can change the community composition.
In the present study, traffic noise altered the bird community composition in sites near the highway. Species with low frequency songs or calls were only detected in quieter sites far from the highway. Other studies have also shown that species with low-frequency calls (e.g. doves, owls, corvids, woodpeckers, herons and rails) are absent in areas with traffic noise (<5 KHz) (Francis et al. 2009; Parris and Schneider 2008; Rheindt 2003).
In contrast, anuran species have restricted ranges and low dispersal capacity; therefore, if background noise disrupts acoustic communication these animals cannot move easily to quieter sites (Gerhardt and Huber 2002). But it appears that anurans species in Puerto Rico may be adapted to high levels of noise because in natural conditions they often occur at very high population densities (20,000 animals per hectare) (Stewart and Woolbright 1996), and males can emit calls between 90 and 95 dB (Narins 1995), which can create very noisy condition in the island’s forests.
Alternative hypotheses that could explain the differences in species diversity between sites near and far from the highway include our sampling method and edge effects. In this study we sampled the anuran and bird communities using automated recording devices. This method will only detect species that are calling, and the sampling area will depend on the frequency and power of the call. We assumed that the sampling radius was approximately 20 m; however, it is possible that the recorders detected individuals more than 20 m away. Some recording sites near the highway were located between 20 and 30 from the forest edge; therefore if the recorders sampled a larger area (i.e. >20 m radius), then the sampling area would have extended outside of the forest. Despite these limitations, the use of automated recorder devices has been shown to be more effective for detecting species in comparison with traditional methodology (e.g. point counts) (Acevedo and Villanueva-Rivera 2006). Furthermore, these recorders permit simultaneous sampling in multiple sites.
Edge effect cause alterations in biotic and abiotic conditions and can change species abundance, distribution, and interactions between them. In this study, all forest sites near and far from the highway were similar in vegetation structure and microclimatic characteristics, but there were differences in the distance to the nearest edge. Sites near the highway were on average 47 m from the nearest edge and sites far from the highway were on average 160 m from the nearest edge (Appendix 1). This difference in distance to the forest edge could be important for birds that are restricted to interior forest habitat, but all bird species detected only in sites far from the highway are species present in areas with high rate of forest fragmentation and do not appear to avoid edges (Pardieck et al. 1996; Suarez-Rubio and Thomlinson 2009; Wunderle et al. 1987). Although the sampling scheme and edge effect could influence the results, our analyses show that anthropogenic noise was the major factor affecting the distribution and composition of birds in these urban forests. By interfering or masking important social signals, traffic noise is reducing the diversity of birds near roads; specifically species with low frequency songs.
References
Acevedo MA, Restrepo C (2008) Land-cover and land-use change and its contribution to the large-scale organization of Puerto Rico’s bird assemblages. Diversity and Distributions 14:114–122
Acevedo MA, Villanueva-Rivera IJ (2006) Using automated digital recording systems as a effective tools for the monitoring of birds and amphibians. Wild Life Society Bulletin 34:211–214
Alicea-Pou J, Viñas-Curiel O, Cruz-Vizcarrondo W, Alomar O (2006) Monitoring of the environmental noise level in San Juan, Puerto Rico. Junta de Calidad Ambiental de Puerto Rico
Amézquita A, Höld W, Lima AP, Castellanos L, Erdtmann LK, Araújo MC (2006) Masking interference and the evolution of the acoustics communication system in the Amazonian dendrobatid frog Allobates femoralis. Evolution 60:1874–1887
Barber JR, Crooks KR, Fristrup KM (2010) The cost of chronic noise exposure for terrestrial organisms. Trends in Ecology and Evolution 25:180–189
Bee MA, Swanson EM (2007) Auditory masking of anuran advertisement calls by highway traffic noise. Animal Behaviour 74:1765–1776
Benitez-López A, Alkemade R, Verweij PA (2010) The impact of roads and other infrastructure on mammal and bird populations: a meta-analysis. Biological Conservation 143:1307–1316
Bergen F, Abs M (1997) Etho-ecological study of the singing activity of the blue tit Parus caeruleus, great tit Parus major and chaffinch Fringilla coelebs. Journal für Ornithologie 138:451–467
Berglund B, Lindvall T (1995) Community noise. Archives of the Center for Sensory Research 2:1–195
Bermúdez-Cuamatzin E, Ríos-Chelén AA, Gil D, Garcia CM (2009) Strategies of song adaptation to urban noise in the house finch: syllable pitch plasticity or differential syllable use. Behaviour 146:1269–1286
Boeckle M, Preininger D, Höld W (2009) Communication in noisy environments I: acoustic signals of Staurois latopalmatus Boulenger 1887. Herpetologica 65:154–165
Brumm H (2004) The impact of environmental noise on song amplitude in a territorial bird. Journal of Animal Ecology 73:434–440
Brumm H, Slabbekoorn H (2005) Acoustic communication in noise. Advances in Study of Behavior 35:151–209
Brumm H, Slater PJB (2006) Ambient noise, motor fatigue, and serial redundancy in chaffinch song. Behavior Ecology and Sociobiology 60:475–481
Brumm H, Schmidt R, Schrader L (2009) Noise-dependent vocal plasticity in domestic fowl. Animal Behavior 78:741–746
Departamento de Transportación y Obras Públicas de Puerto Rico DTOP (2009) http://www.dtop.gov.pr/
Dooling RJ, Popper A (2007) The effect of highway noise on birds. The California Department of Transportation Division of Environmental Analysis, Sacramento
Dorcas ME, Price SJ, Walls SC, Barichivich WJ (2009) Auditory monitoring of anuran populations. In: Dood CK Jr (ed) Amphibian ecology and conservation: a hand book of techniques. Oxford University Press, Oxford, pp 281–298
Eigenbrod F, Hecnar SJ, Fahrig L (2008) The relative effects of highway traffic and forest cover on anuran populations. Biological Conservation 141:35–46
Ewel JJ, Whitmore JL (1973) The ecological life zones of Puerto Rico and the U.S. Virgin Islands. Forest Service, U.S. Department of Agriculture. Institute of Tropical Forestry, Rio Piedras, Puerto Rico.
Forman RTT, Deblinger RD (2000) The ecological highway-effect zone of a Massachusetts U.S.A. suburban highway. Conservation Biology 14:36–46
Francis CD, Ortega CP, Cruz A (2009) Noise pollution changes avian communities and species interactions. Current Biology 19:1415–1419
Fuller RA, Warren PH, Gaston KJ (2007) Daytime noise predicts nocturnal singing in urban robins. Biology Letters 3:368–370
Gerhardt HC, Huber F (2002) Acoustic communication in insects and anurans: common problems and diverse solutions. University of Chicago Press, Chicago
Gould W, Alarcó C, Fevold B, Jiménez ME, Martinuzzi S, Potts G, Solórzano M, Ventosa E (2007) Puerto Rico Gap Analysis Project–Final Report. USGS, Moscow ID and the USDA FS International Institute of Tropical Forestry, Río Piedras, PR. 157 p and 8 appendices
Hoelzel AR (1986) Song characteristics and response to playback of male and female robins Erithacus rubecula. Ibis 128:115–127
Kaiser K, Hammers JL (2009) The effect of anthropogenic noise on male advertisement call rate in the neotropical treefrog, Dendropsophus triangulum. Behaviour 146:1053–1069
McCune B, Grace JB (2002) PC-ORD: multivariate analysis of ecological data. Version 5. User’s guide. MjM Software Design, Glendel Beach, Oregon
Myers N, Mittermeier RA, Mittermeier CG, da Fonseca GAB, Kent J (2000) Biodiversity hotspot for conservation priorities. Nature 403:853–858
Narins P (1995) Frog communication. Scientific American 273:62–67
National Oceanic and Atmospheric Administration NOOA (2010) http://www.weather.gov/climate/index.php?wfo=sju
Pardieck KL, Meyers JM, Pagán M (1996) Surveys of Puerto Rican Screech-Owl populations in large-tract and fragmented forest habitats. The Wilson Bulletin 108:776–782
Parris KM, Schneider A (2008) Impacts of traffic noise and traffic volume on birds of highway side habitats. Ecol Soc 14:29 http://www.ecologyandsociety.org/vol14/iss1/art29/
Parris KM, Velik-Lord M, North JMA (2009) Frogs call at a higher pitch in traffic noise. Ecol Soc 14:25 http://www.ecologyandsociety.org/vol14/iss1/art25/
Peris SJ, Pescador M (2004) Effects of traffic noise on passerine populations in Mediterranean wooded pastures. Applied Acoustics 65:357–366
Raffaele HA (1989) A guide to the birds of Puerto Rico and the Virgin Islands. Princeton University Press, Princeton, 254 p
Reijnen R, Foppen R, Ter Braak C, Thissen J (1995) The effects of car traffic on breeding bird populations in woodland. III. Reduction of density in relation to the proximity of main highways. Journal of Applied Ecology 32:187–202
Rheindt FE (2003) The impact of highways on birds: does song frequency play a role in determining susceptibility to noise pollution? Journal of Ornithology 114:295–306
Rivero JA (1998) Los anfibios y reptiles de Puerto Rico. Editorial de la Universidad de Puerto Rico. San Juan, Puerto Rico. 510 pp
Slabbekoorn H (2004) Habitat-dependent ambient noise: consistent spectral profiles in two African forest types. Journal of Acoustical Society of America 116:3727–3733
Slabbekoorn H, Peet M (2003) Birds sing at a higher pitch in urban noise. Nature 424:267
Slabbekoorn H, Ripmeester EAP (2008) Birdsong and anthropogenic noise: implications and applications for conservation. Molecular Ecology 17:72–83
Stewart MM, Woolbright LL (1996) Amphibians. In: Reagan DP, Waide RW (eds) The food web of a tropical rain forest. University of Chicago Press, Chicago, pp 273–320
Stone E (2000) Separating the noise from the noise: a finding in support of the ¨niche hypothesis¨, that birds are influenced by human-induced noise in natural habitats. Anthrozoos 13:225–231
Suarez-Rubio M, Thomlinson JR (2009) Landscape and patch-level factor influence bird communities in an urbanized tropical island. Biological Conservation 142:1311–1321
Sun JWC, Narins PM (2005) Anthropogenic sounds differentially affect de amphibian call rates. Biological Conservation 121:419–427
Warren PS, Katti M, Ermann M, Brazel A (2006) Urban bioacoustics: it’s not just noise. Animal Behaviour 71:491–502
Wood WE, Yezerinac SM (2006) Song sparrow Melospiza melodia song varies with urban noise. Auk 123:650–659
Wunderle JM, Diaz A, Velasquez I, Scharrón R (1987) Forest openings and the distribution of understory birds in a Puerto Rican rainforest. The Wilson Bulletin 99:22–37
Zimmerman GM, Goetz H, Mielke PW Jr (1985) Use of an improved statistical method for group comparisons to study effects of prairie fire. Ecology 66:606–611
Acknowledgements
We thank Laura May-Collado and Joseph Wunderle for their comments on the manuscript. We thank A.P. Valencia, C. Sanfiorenzo, A. Herrera-Montes, J. Figueroa, S.V. Ramirez, B. Hilje, J. Fonseca and C.A. Rodriguez for their help in the field. We thank C. Milan and the ARBIMON team for their advice with recorders and recording software analysis. We thank P. Rincon and the International Institute of Tropical Forestry (IITF), USDA Forest Service, Puerto Rico for their help with the map. The research was funded by National Science Foundation (BDI 0640143), Junta de Calidad Ambiental de Puerto Rico and Ford Motor Company. Finally, we appreciate the comments and suggestions of two anonymous reviewers and the Editor that helped us improve the manuscript.
Author information
Authors and Affiliations
Corresponding author
Electronic supplementary material
Below is the link to the electronic supplementary material.
Appendix 1
(DOC 74 kb)
Rights and permissions
About this article
Cite this article
Herrera-Montes, M.I., Aide, T.M. Impacts of traffic noise on anuran and bird communities. Urban Ecosyst 14, 415–427 (2011). https://doi.org/10.1007/s11252-011-0158-7
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11252-011-0158-7