
Abstract
The characterisation of the small ruminant populations in developing countries will play a major role in the maintenance of the genetic resources as the basis for future improvement in livestock production. The present study aimed at morphological characterisation of the two main breeds of sheep in Ghana by assessing variation within and between breed populations using principal component and discriminant analyses. The two breeds were the Sahel and the Djallonke sheep of both sexes and of two groups namely, young (1 year old, consisting of 74 animals) and mature sheep (≥2 years old, comprising 219 animals). The analysis of variance revealed significant (P < 0.05) differences in the morphological traits of the Sahel and the Djallonke sheep breeds with higher values recorded for the former. Sexual dimorphism was in favour of male animals in all the morphological traits examined. Mature animals also had comparative advantage over the young. Two principal components were extracted to discern the structure of the two genetic groups. The most discriminating traits between the two sheep breeds were rump height, height at withers, neck girth and pin-bone width. Mahalanobis distance between the two genetic groups was 5.723 (P < 0.0001). The developed discriminant functions clearly discriminated and classified the Sahel and the Djallonke sheep into their breeds of origin, thus yielding 100, 93.4 and 90.4 % accurate classification for the rams, ewes and the overall sheep population, respectively. The present approach would greatly help in establishing management and conservation policies for the sustainable production of the two Ghanaian sheep breeds.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Domestic animal diversity is a term that has been used to mean the genetic differences among and within breeds of species used for food and agriculture (Cardellino and Boyazoglu 2009). Sheep rearing is one of the most important means of livelihood and food security for majority of the rural populace, especially in developing countries (Amadou et al. 2012). There are two main sheep breeds in Ghana namely, the West African Dwarf (Djallonke) or the Djallonke sheep and the West African Long-Legged (Sahel) or the Sahelian sheep, although hybrids from two or more different breeds also exist. A bulk of the sheep is confined to the rural areas where almost every family owns a few animals. The small size and early maturity of sheep gives them several distinct economic advantages in smallholder situations. In animal production systems, the value of a species increases in relation to its adaptation, capacity to make socioeconomic contributions, capacity to fill market opportunities and potential for increasing productivity. These are buttressed by the fact that livestock are expected to balance a range of policy objectives among which are supporting rural development and the alleviation of hunger and poverty, meeting the increasing demand for livestock products and responding to changing consumer requirements, ensuring food safety and minimising the threat posed by animal diseases, and maintaining biodiversity and environmental integrity (FAO 2011a). There is a considerable potential for increased sheep production, if proper management is employed. At present, many countries are losing their genetic resources which may have lasting effects on food security and sustainable development especially in the light of global warming changes (Hoffman 2010). The rationale behind conservation of farm animal diversity is that humankind may need to keep this specific genetic biodiversity to face future (unknown) challenges such as changes in demand for livestock products, spread of new diseases, reducing environmental impact and climate change (Thornton 2010; FAO 2011a, b). Analyses of morphometric variables that are easy to measure make it possible to explore areas such as the structure of breeds, the degree of variability between various populations, the harmony of morphological models and the definition of morphological models for a given breed. Hence, it is important to accurately analyse the morphological variables that enable us to distinguish between breeds, as well as explore the use of various discrimination methods to assess the potential of each of the variables under study (Rodero et al. 2011). Measurement of linear body parameters has been used to estimate necessary size in sheep. The use of quantitative information in livestock breeding programmes has become more sophisticated over time. This allows breeders to make faster progress in a chosen set of traits. Phenotypic information was initially used in mass selection, whereby individuals with better trait values were chosen to be parents of the next generation (Carneiro et al. 2010). This model has worked remarkably well and has allowed much progress in genetic merits.
Indigenous livestock breeds of Africa are well adapted to the local environment even though their productivity is generally lower when compared to other parts of the world. Attempts by breeders and farmers to improve the performance of the indigenous African breeds involve the introduction of exotic animals and crossbreeding practices, which is gradually leading to the erosion and complete masking of important survival traits, such as disease resistance associated with indigenous livestock as well as the extinction of certain breeds (Gizaw et al. 2011). The characterisation of African small ruminant populations will play a major role in the maintenance of these autochthonous genetic resources as the basis for future improvement at both the production and the genetic levels. This can be partly achieved through the analyses of morphological traits to assess variation within and between populations using classical multivariate analyses such as principal components and discriminant analysis, which have been shown to be suitable in assessing variation within and can discriminate different population types when all measured morphological variables are considered simultaneously (Yakubu et al. 2010a; Legaz et al. 2011). The principal component technique can reduce the information contained in the original complex of variables by eliminating redundant information due to correlation among them (Cerqueira et al. 2011; Yakubu et al. 2011a). Discriminant function analysis is a statistical technique that allows new individuals to be assigned to previously established or defined groups. The analysis is based on a set of data from n individuals for which p quantitative variables (independent variables) have been measured as a profile for each of them. On the other hand, an additional qualitative variable (dependent variable), with two or more categories and defined by other means, groups each individual in a category (Rodero et al. 2011). This produces an n × (p + 1) table in which each case has a profile and is assigned to one group. From this table, a discriminant model is obtained to compare to the profile of new individuals. The morphological characterisation of African animal genetic resources is currently receiving increased attention (Traoré et al. 2008; Yakubu and Akinyemi 2010; Yakubu and Ibrahim 2011).
Therefore, this study aimed at distinguishing between the two main local sheep breeds in Northern Ghana using principal component and discriminant analyses. The information obtained may be useful in designing appropriate management and conservation strategies for the autochthonous breeds in Ghana and the sub-Saharan Africa.
Materials and methods
Management of experimental sheep
A total of 293 sheep (230 Djallonke and 63 Sahel) of both sexes (36 males and 257 females) were used for the study. The animals were divided into two groups namely, young (1 year old, consisting of 74 animals) and mature sheep (≥2 years old, comprising 219 animals). The sheep were managed semi-intensively. They were housed in properly constructed pens throughout the night and sometimes during the day when there was the need to restrict their movement. Feed and water were provided for the sheep ad libitum throughout the year. Conventional disease and pest control regimes were practised.
Data collection
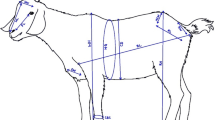
The variables measured included live weight (LW) (in kilogram) and linear body measurements (in centimetre) namely, height at withers (HW), rump height (RH), body length (BL), heart girth (HG), neck girth (NG), chest depth (CD) and pin-bone width (PBW). The anatomical reference points (Hamito 2009) were as follows:
-
HW: The distance from the surface of a platform on which an animal stands, to the withers of the animal.
-
RH: This is the distance from the surface of a platform to the rump.
-
BL: The distance from the base of the tail to the base of the neck.
-
HG: The circumference around the chest just behind the front legs and withers.
-
NG: This is the circumference around the neck near the withers.
-
CD: The distance from the backbone at the shoulder to the brisket between the front legs
-
PBW: It is the distance between the outer edges of the major hip bones on the right and left sides.
Statistical analysis
Data were analysed using the general linear model procedure of Statistical Package for Social Sciences (SPSS 2010) to test the fixed effects of breed, sex, age and their interactions. Means were separated using least significant difference and tested at 95 % confidence interval. The general model employed was the following:
-
Y ijk = individual observation
-
μ = overall mean
-
B i = fixed effect of ith breed (i = Djallonke, Sahel)
-
S j = fixed effect of jth sex (j = male, female)
-
A k = fixed effect of kth age (k = 1 year old, ≥2 years old)
-
(BS) ij = interaction effect of ith breed and jth sex
-
(BA) ik = interaction effect of ith breed and kth age
-
(SA) jk = interaction effect of jth sex and kth age
-
(BSA) ijk = interaction effect of breed, sex and age
-
e ijk = random error associated with each record (normally, independently and identically distributed with zero mean and constant variance)
The PRINCOMP procedure was carried out to investigate the core structure of the sheep traits in order to check whether the traits could be reduced to uncorrelated dimensions (principal components (PCs)). In the PC analysis, Kaiser–Meyer–Olkin (KMO) measures of sampling adequacy, Bartlett's test of sphericity (tests the null hypothesis that the original correlation matrix is an identity matrix) and communality were computed to test the validity of the principal component factor analysis of the data set. Cumulative proportion of variance criterion was employed in determining the number of factors to extract. The varimax criterion of the orthogonal rotation method was employed in the rotation of the factor matrix to enhance the interpretability of the factor analysis. The overall reliability of the PC analysis was tested using Chronbach's alpha. For more effective discrimination and classification of the sheep into Djallonke and Sahel, traits' reduction was obtained using PROC STEPDISC of Statistical Analysis System (SAS 2003). The CANDISC procedure was used to generate the univariate test of the selected traits, canonical functions and the plots of breed membership. The PROC DISCRIM was employed to obtain the Mahalanobis distances and linear discriminant functions. The ability of these functions to identify Djallonke and Sahel sheep (both sex inclusive) was indicated as the percentage of individuals correctly classified from the samples that generated the functions. The honesty (reliability testing) of the functions was validated using split-sample validation (cross-validation). The proportion of individuals correctly reallocated is taken as a measurement of the integrity of that group. The number of misclassified individuals indicates the degree of intermingling between the populations.
The initial discriminant function model for separating the two breeds of sheep incorporated seven traits (HG, NG, CD, HW, RH, BL and PBW):
Where z i is the discriminant score for the ith individual and e i is the contribution (magnitude) of the ith trait on the discriminant function.
Results
Morphological traits
The analysis of variance of live weight and linear body measurements of Djallonke and Sahel sheep are presented in Tables 1 and 2. All morphological traits were significantly (P < 0.05) influenced by breed, with higher values recorded for Sahel sheep compared to their Djallonke counterparts. The body dimensions of male sheep were also higher (P < 0.05) than those of the females. Similarly, mature animals had higher (P < 0.05) mean values for all body measurements than young animals. Breed × sex, breed × age, sex × age and breed × sex × age interaction effects were significant for most of the traits investigated.
Principal component analysis
KMO (0.910), Bartlett's test of sphericity (chi-square = 3,076.55, P < 0.001), communality (0.828–0.934) and Cronbach's alpha (0.822) provided support for the validity and reliability of the factor analysis of the data set. The first two PCs explained 87.191 % of all variation between the traits measured (Table 3). While PC1 was characterised by LW, HW, RH, BL, HG, NG and CD, PC2 had its loading for PBW only.
Discriminant analysis
Through the stepwise procedure, only four morphometric traits (RH, HW, NG and PBW) were significant in discriminating between the two breeds of sheep in each sample except in the rams where NG was not significant (Table 4). Interestingly, HG was an important discriminator only for rams. Using the standardised discriminant function coefficients which show the multivariate (joint) discriminating power of all the morphometric traits, the derived discriminant functions for the rams, ewes and the overall sheep, respectively, are
where z is the discriminant score for individual sheep.
Clear morphological differences existed between Sahel and Djallonke because plots of breed membership using the canonical variables (Figs. 1, 2 and 3) indicated that there were two distinct populations. The Mahalanobis distance between the Djallonke and Sahel sheep was 5.723 (Table 5). While 100 % of the rams were classified into the appropriate breeds of origin, 93.4 % of the ewes were correctly assigned (Table 6). Overall, 90.8 % of the sheep (69.8 % of Sahel and 96.5 % of Djallonke) were correctly classified into their source genetic groups.

Plot of breed membership for rams

Plot of breed membership for ewes

Plot of breed membership for the ‘overall’ sheep
The Fisher's linear discriminant functions that were developed and used for the classifications are as follows:
The critical values for the rams, ewes and the overall sheep are 98.60, 35.56 and 28.25, respectively, so that an observation is classified as Sahel if its z* score is greater than the associated critical value; otherwise, it is classified into Djallonke.
Discussion
Genetic variation is vital for the populations to adapt to varying environments and to respond to artificial selection; therefore, any conservation and development scheme should start from assessing the state of variation in the population (Toro et al. 2011). Production of the autochthonous Ghanaian sheep breeds is directly associated with the concept of sustainable agriculture. The difference between the two sheep breeds may have genetic underpinning (Yakubu and Ibrahim 2011), which is often facilitated when measurements are restricted to phenotypically pure animals (Yakubu et al. 2010b). The differentials obtained in the morphological traits of the sexes could be attributed to sexual dimorphism (Yakubu and Akinyemi 2010; Carneiro et al. 2010). Festa-Bianchet et al. (1996) reported that most dimorphism developed post-weaning because of faster mass gain by males during the age of 1–2 years. They also suggested that males might have a longer season of mass gain each year throughout their lives, while females divert annual resources into reproduction, rather than body mass. The superiority of mature animals over the young may be due to the effect of age as an important factor influencing the conformation traits of animals. The significant interaction effects indicate the separate rankings of the breeds under the two sexes and ages investigated.
The results for size and shape presented here indicate that there are some significant differences between populations. The first PC showed that an animal large for one trait was generally large for all. The second described a subgroup of animals with wide hips. The result of this study is consistent with the reports of earlier workers (Chacon et al. 2011; Cerqueira et al. 2011) where two PCs were retained to explain the variations accounted for by the traits. While the first PC indicates general body size (Pundir et al. 2011; Yakubu et al. 2011b), the second PC represents shape component (Yakubu et al. 2011b).
The RH was the single most important trait that clearly differentiates Sahel from Djallonke sheep breed while HW was a very important discriminator in the ewes and in the overall sheep population. This is laudable because RH and HW represent height and Sahel sheep are generally taller than Djallonke sheep when animals of equal age groups of both breeds are compared and appraised visually or on morphometric basis (Traoré et al. 2008). The present finding on the use of height measurement for discriminating between sheep breeds is consistent with the report of earlier workers (Carneiro et al. 2010; Rodero et al. 2011). The presence of PBW and NG in the functions implies that there is a significant difference between the two breeds in broadness by the neck and pin bone; hence, these breeds can be discriminated on the basis of their anterior and posterior dorso-lateral condition. The traits that caused the marked difference were among those proposed by Yokoo et al. (2010) who suggested the inclusion of linear dimensions in breeding programmes because such traits were easily measured and less affected by environmental variations. A clear distinction between breeds is very necessary to the livestock farmers and livestock product consumers as well as livestock breeders and researchers. The magnitude of such knowledge is well presented in the developed countries where breed names become more widely used as ‘brand’ names for livestock products (Blott et al. 1999). It, therefore, suffice to say that a parsimonious discrimination between sheep breeds may be achieved by using few discriminant traits provided those traits are carefully selected through a statistically appraised procedure. This is buttressed by the reports of Carneiro et al. (2010) in sheep, Yakubu et al. (2011a) in goats and Kefena et al. (2011) in donkeys.
Expectedly, Sahel rams in the present study were higher in height (RH) and larger in size (NG, HG and PBW) than the Djallonke rams. This observation was similar to that of Yakubu et al. (2010a) where the use of rump height, body length, horn length, face length, chest girth, neck circumference and head width as the most discriminant variables separating Djallonke and Red Sokoto goats yielded 100 % correct classification. The considerable (30.2–31.7 %) erroneous classification of Sahel as Djallonke may mean that gene introgression is implicated since some of the breeding stocks were brought to the experimental site (Pong-Tamale Livestock Breeding Station) from elsewhere. Besides, some attempts have been made by the stockmen at the breeding station to crossbreed, which suggests that the genetic resources of these two breeds of sheep might be eroded. Traoré et al. (2008) have attributed the cause of large misclassifications to the manifestation of introgressions across breeds resulting from the actions of most stock breeders who intend to obtain products with bigger conformation. The lower misclassification errors of the Djallonke breed in each sample may be an indication of more uniformity as a result of more genetic homogeneity of this breed than the Sahel. It confirms the assertion that the Djallonke sheep is generally small in body size (Traoré et al. 2008), and so most of them could not have been wrongly categorized as belonging to the large body-sized Sahel breed.
Livestock biodiversity would be the source pool for genes that confer disease resistance, specific product qualities like fatty acid composition or milk composition, resistance to draught and high temperatures and production traits to be combined in newly formed breeds or crossbreds (Cardellino and Boyazoglu 2009). The characterisation of sheep in this study will be helpful to livestock farmers and researchers in preserving the genetic resources of some of the indigenous African sheep breeds, as well as to farmers and dealers in livestock products in the production, processing and marketing of livestock and livestock products. However, whether the variations in these morphological traits are caused by adaptive or non-adaptive sources needs to be further verified by comparing between relative levels of population divergence in quantitative traits and neutral DNA markers.
Conclusion
The study revealed that the Sahel sheep were larger than their Djallonke counterparts in all the eight body measurements investigated. The structure of the sheep was explained using two principal components. Rump height, height at withers, neck girth and pin-bone width were found as the most discriminating variables to separate the two sheep breeds. Most of the animals were correctly assigned into their breeds of origin. The present information when complemented with DNA microsatellites may help in management and in situ conservation of the Djallonke and Sahel sheep in Ghana.
Abbreviations
- LW:
-
Live weight
- HW:
-
Height at withers
- RH:
-
Rump height
- BL:
-
Body length
- HG:
-
Heart girth
- NG:
-
Neck girth
- CD:
-
Chest depth
- PBW:
-
Pin-bone width
References
Amadou, H., Dossa, L. H., Lompo, D. J., Abdulkadir, A. and Schlecht E. 2012. A comparison between urban livestock production strategies in Burkina Faso, Mali and Nigeria in West Africa, Tropical Animal Health and Production, Available at: doi:10.1007/s11250-012-0118-0
Blott, S. C., Williams, J. L. and Haley, C. S., 1999. Discriminating among cattle breeds using genetic markers, Heredity, 82, 613-619
Cardellino, R. A. and Boyazoglu, J., 2009. Research opportunities in the field of animal genetic resources, Livestock Science, 120, 166–173
Carneiro, H., Louvandini, H., Paiva, S. R., Macedo, F., Mernies, B. and McManus, C., 2010. Morphological characterization of sheep breeds in Brazil, Uruguay and Colombia, Small Ruminant Research, 94, 58-65
Cerqueira, J. O. L., Feas, X., Iglesias, A., Pacheco, L. F. and Araujo, J. P. P., 2011. Morphological traits in Portuguese Bordaleira de Entre Douro e Minho sheep: divergence of the breed, Animal Reproduction and Science, 51, 635-641
Chacon, E., Macedo, F., Velazquez, F., Paiva, S. R., Pineda, E. and McManus, C., 2011. Morphological measurements and body indices for Cuban Creole goats and their crossbreds, Revista Brasileira de Zootecnia, 40, 1671-1679
FAO., 2011a. Draft guidelines on molecular genetic characterization of animal genetic resources, Commission on Genetic Resources for Food and Agriculture, 13th Regular Session, 18-22 July, 2011, Rome. Available at: http://www.fao.org/docrep/meeting/022/am652e.pdf
FAO., 2011b. Draft guidelines on phenotypic characterization of animal genetic resources, Commission on Genetic Resources for Food and Agriculture, 13th Regular Session, 18-22 July, 2011, Rome. Available at http://www.fao.org/docrep/meeting/022/am651e.pdf
Festa-Bianchet, M., Jorgenson, J. T., King, W. J., Smith, K. G. and Wishart, W. D., 1996. The development of sexual dimorphism: Seasonal and lifetime mass changes in bighorn sheep, Canadian Journal of Zoology, 74, 330-342
Gizaw, S., Komen, H., Hanote, O., van Arendonk, J. A. M., Kemp, S., Haile, A., Mwai, O. and Dessie, T., 2011. Characterization and conservation of indigenous sheep genetic resources: A practical framework for developing countries. ILRI Research Report No. 27. Nairobi, Kenya, ILRI
Hamito, D., 2009. Estimation of weight and age of sheep and goats. Ethiopia sheep and goat productivity improvement program (ESGPIP), Technical bulletin No. 23, 11
Hoffman, I., 2010. Climate change and the characterization, breeding and conservation of animal genetic resources. Animal Genetics, 41, 32-46
Kefena, E., Beja-Pereira, A., Han, J. L., Haile, A., Mohammed, Y. K. and Dessie, T., 2011. Eco-geographical structuring and morphological diversities in Ethiopian donkey populations, Livestock Science, 141, 232-241
Legaz, E., Cervantes, I., Perez-Cabal, M. A., de la Fuente, L. F., Martinez, R., Goyache, F. and Gutierrez, J. P., 2011. Multivariate characterisation of morphological traits in Assaf (Assaf.E) sheep, Small Ruminant Research, 100, 122-130
Pundir, R. K., Singh, P. K., Singh, K. P. and Dangi, P. S., 2011. Factor analysis of biometric traits of Kankrej cows to explain body conformation, Asian-Australian Journal of Animal Science, 24, 449-456
Rodero, E., Gonzalez, A., Luque, M., Herrera, M. and Gutierrez-Estrada, J. C., 2011. Classification of Spanish autochthonous bovine breeds: Morphometric study using classical and heuristic techniques, Livestock Science, doi:10.1016/j.livsci.2011.09.022
SAS., 2003. Statistical Analysis System. SAS Stat. SAS Institute Incorporation, Cary NC 27513, USA
SPSS., 2010. Statistical Package for Social Sciences. SPSS Incorporation, 444 Michigan Avenue, Chicago, IL60611, USA
Thornton, P. K., 2010. Livestock production: recent trends, future prospects, Philosophical Transactions of the Royal Society B: Biological Sciences, 365, 2853-2867
Toro, M. A., Meuwissen, T. H. E., Fernandez, J., Shaat, I. and Maki-Tanila, A., 2011. Assessing the genetic diversity in small farm animal populations, Animal, 5, 1669–1683
Traoré, A., Tamboura, H. H., Kabore, A., Royo, L. J., Fernandez, I., Álvarez, I., Sangare, M., Bouchel, D., Poivey, J. P., Francois, D., Toguyeni, A., Sawadogo, L. and Goyache, F., 2008. Multivariate characterization of morphological traits in Burkina Faso sheep, Small Ruminant Research, 80, 62-67
Yakubu, A. and Akinyemi, M. O., 2010. An evaluation of sexual size dimorphism in Uda sheep using multifactorial discriminant analysis, Acta Agriculturae Scandinavica, Section A-Animal Science, 60, 74-78
Yakubu, A. and Ibrahim, I. A., 2011. Multivariate analysis of morphostructural characteristics in Nigerian indigenous sheep, Italian Journal of Animal Science, 10, 83-86
Yakubu, A., Idahor, K. O., Haruna, H. S., Wheto, M. and Amusan, S. 2010b. Multivariate analysis of phenotypic differentiation in Bunaji and Sokoto Gudali cattle, Acta Argiculturae Slovenica, 96/2, 75-80
Yakubu, A., Salako, A. E., Imumorin, I. G., Ige, A. O. and Akinyemi, M. O., 2010a. Discriminant analysis of morphometric differentiation in the West African Dwarf and Red Sokoto goats, South African Journal of Animal Science, 40, 381-387
Yakubu, A., Salako, A. E. and Imumorin, I. G., 2011a. Comparative multivariate analysis of biometric traits of West African Dwarf and Red Sokoto goats, Tropical Animal Health and Production, 43, 561-566
Yakubu, A., Salako, A. E. and Abdullah, A. R., 2011b. Varimax rotated principal component analysis of the zoometrical traits of Uda sheep, Archivos de Zootecnia, 60, 813-816
Yokoo, M. J., Lobo, R. B., Araujo, F. R. C., Bezerra, L. A. F., Sainz, R. D. and Albuquerque, L. G., 2010. Genetic associations between carcass traits measured by real-time ultrasound and scrotal circumference and growth traits in Nelore cattle, Journal of Animal Science, 88, 52-58
Acknowledgments
The authors would like to thank the staff of the Pong-Tamale Livestock Breeding Station especially the head of the sheep unit, Mr. Lawrence Dartay, and the manager, Mr. Ibrahim Shahadu, for providing the experimental animals and being instrumental in the data gathering process.
Author information
Authors and Affiliations
Corresponding author
Additional information
An erratum to this article is available at http://dx.doi.org/10.1007/s11250-014-0555-z.
Rights and permissions
About this article
Cite this article
Birteeb, P.T., Peters, S.O., Yakubu, A. et al. Multivariate characterisation of the phenotypic traits of Djallonke and Sahel sheep in Northern Ghana. Trop Anim Health Prod 45, 267–274 (2012). https://doi.org/10.1007/s11250-012-0211-4
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11250-012-0211-4