
Abstract
Growth hormone (GH) transgenic fish are at a critical step for possible approval for commercialization. Since this hormone is related to salinity tolerance in fish, our main goal was to verify whether the osmoregulatory capacity of the stenohaline zebrafish (Danio rerio) would be modified by GH-transgenesis. For this, we transferred GH-transgenic zebrafish (T) from freshwater to 11 ppt salinity and analyzed survival as well as relative changes in gene expression. Results show an increased mortality in T versus non-transgenic (NT) fish, suggesting an impaired mechanism of osmotic acclimation in T. The salinity effect on expression of genes related to osmoregulation, the somatotropic axis and energy metabolism was evaluated in gills and liver of T and NT. Genes coding for Na+, K+-ATPase, H+-ATPase, plasma carbonic anhydrase and cytosolic carbonic anhydrase were up-regulated in gills of transgenics in freshwater. The growth hormone receptor gene was down-regulated in gills and liver of both NT and T exposed to 11 ppt salinity, while insulin-like growth factor-1 was down-regulated in liver of NT and in gills of T exposed to 11 ppt salinity. In transgenics, all osmoregulation-related genes and the citrate synthase gene were down-regulated in gills of fish exposed to 11 ppt salinity, while lactate dehydrogenase expression was up-regulated in liver. Na+, K+-ATPase activity was higher in gills of T exposed to 11 ppt salinity as well as the whole body content of Na+. Increased ATP content was observed in gills of both NT and T exposed to 11 ppt salinity, being statistically higher in T than NT. Taking altogether, these findings support the hypothesis that GH-transgenesis increases Na+ import capacity and energetic demand, promoting an unfavorable osmotic and energetic physiological status and making this transgenic fish intolerant of hyperosmotic environments.
Similar content being viewed by others


Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
Advances in biotechnological methods, especially fish transgenesis, are potentially important for increasing production and economic efficiency in aquaculture. Transgenic techniques may provide novel aquaculture methods. A variety of transgenes have been tested to improve production traits, such as growth, maturation, resistance to diseases and freezing (Ayoola and Idowu 2008). Since growth is the most important trait for aquaculture, transgenic fish with improved growth have been produced through gene transfer. Enhanced growth using the growth hormone (GH) gene has been obtained in several fish species, such as rainbow trout (Pitkänen et al. 1999), Atlantic salmon (Du et al. 1992), Nile tilapia (Rahman et al. 1998), coho salmon (Devlin et al. 1994; Devlin et al. 1999), Arctic charr (Pitkänen et al. 1999), and mud loach (Nam et al. 2001), among others.
Additionally to its central role in regulating growth, GH is involved in other physiological processes, such as appetite and food intake (Guan et al. 2008; Higgs et al. 2009), metabolism (Leggatt et al. 2003; Guan et al. 2008) and osmoregulation (for reviews see Sakamoto and McCormick 2006; Hallerman et al. 2007). In fact, several studies carried out with salmonids and other euryhaline teleosts have shown that an increase in salinity tolerance is achieved through a proliferation of chloride cells and stimulation of Na+, K+-ATPase activity in gills by the GH/IGF axis (Bolton et al. 1987; Sakamoto and Hirano 1993; McCormick 1996; Mancera and McCormick 1998).
Insulin-like growth factor -1 (IGF1) is the mediator of most GH effects and is involved with osmoregulatory actions of growth hormone (Reinecke 2010). GH transgenesis in fish has increased IGF1 expression in liver and various extrahepatic sites of adult GH-overexpressing tilapia (Eppler et al. 2010). GH treatment also enhanced tilapia Oreochromis niloticus survival in seawater (Xu et al. 1997) and its hypoosmoregulatory ability following hypophysectomy (Sakamoto et al. 1997). Likewise, GH treatment increased salinity tolerance in coho salmon Oncorhynchus kisutch (Shrimpton et al. 1995), rainbow trout Oncorhynchus mykiss (Sangiao-Alvarellos et al. 2005), and the euryhaline killifish Fundulus heteroclitus (Mancera and McCormick 1999).
However, an earlier zebrafish study suggested that GH gene is functionally divergent between zebrafish (a stenohaline fish) and other euryhaline teleosts (Hoshijima and Hirose 2007). In fact, fishes have seawater genetic heritance and transgenesis could alter their gene expression profile and provide the protein machinery needed to change the salt tolerance capacity. An observation supporting this hypothesis is the presence of the NKCC cotransporter gene in the zebrafish genome (Na+, K+, 2Cl− cotransporter—GenBank accession number NM001002080), which is involved mainly in Na+ excretion mechanisms in euryhaline fishes. Accordingly, altering fish growth through GH transgenesis could imply a modification in fish osmoregulatory ability, as well as its tolerance to increasing water salinity. This could prove a useful trait for practical aquaculture, as supplies of clean freshwater are limited, but clean brackish or saltwaters are less limited.
Osmoregulation in fishes occurs mainly in the gills and involves several hormones and enzymes, which can also affect the energetic metabolism. In freshwater fish gills, a decrease in relative permeability can occur as a strategy to reduce ion loss and/or activate mechanisms involved in ion uptake from the water. In this context, Na+, K+-ATPase activity at the basolateral membrane of the gill epithelium is a major component involved in NaCl uptake, since it generates the electrochemical gradient needed for other ion-transporting mechanisms. Regarding Na+ uptake, these mechanisms also include the apical Na+/H+ exchanger (Parks et al. 2008), Na+ channel (ENaC) and the proton pump (H+-ATPase). The proton pump is responsible for proton (H+) secretion, thereby generating a local electrical gradient that promotes Na+ influx across the apical membrane through a Na+ channel (Evans et al. 1999). Likewise, Na+ can exchange with H+ using the Na+/H+ transporter. The intracellular generation of H+ necessary for the proton apical extrusion via H+-ATPase or Na+/H+ exchanger is derived from CO2 hydration catalyzed by cytosolic carbonic anhydrase (CA2). In turn, the CO2 input involves the activity of plasma carbonic anhydrase (CA1) (Breton 2001; Lindskog 1997). In zebrafish, the osmoregulatory mechanism has been investigated through gene expression analysis as Na+, K+-ATPase alpha1a subunit, carbonic anhydrase-1 and 2 (Craig et al. 2007), Na+/H+ exchanger 3b and H+-ATPase (Yan et al. 2007). All these genes encode proteins involved with Na+ uptake. However, zebrafish gills can show a high degree of phenotypic plasticity and gene transcription might be differentially regulated depending on the environmental situation in which fish are acclimated (Craig et al. 2007; Yan et al. 2007).
Considering the fact that environmental risk of transgenic fish is related to overexpression of the transferred gene, our main goal was to verify whether the osmoregulatory capacity of a stenohaline fish (zebrafish—Danio rerio) would be modified by GH transgenesis. For this purpose, the following aspects were evaluated: (1) influence of increasing salinity on growth rate of NT zebrafish; (2) expression of major genes involved in iono-osmoregulatory regulation of zebrafish [Na+, K+ATPase alpha 1 (NKA), H+-ATPase (HATPAse), plasma carbonic anhydrase (CA1), cytosolic carbonic anhydrase (CA2), Na+/H+ exchange 3b (NHE3b)], in gills of NT and T zebrafish; (3) median lethal salinity (MLS) in non-transgenic (NT) zebrafish; (4) tolerance of GH-transgenic (T) zebrafish to MLS; (5) salinity effect on expression of genes related to osmoregulation, somatotropic axis [GH receptor (GHR) and IGF1] and energy metabolism in gills and liver of NT and T zebrafish; and (6) salinity effects on Na+, K+-ATPase activity, whole body sodium concentration, ATP content in gills of NT and T zebrafish.
Materials and methods
Transgenic and non-transgenic fish
T and NT control zebrafish were obtained from crosses between non-transgenic females and hemizygous transgenic males from lineage F0104, following a protocol previously described (Figueiredo et al. 2007a). The F0104 lineage was produced by the co-injection of transgenes cbA/msGH and cbA/GFP. Both transgenes include the carp (Cyprinus carpio) β-actin promoter and either the marine silverside (Odontesthes argentinensis) GH cDNA (Marins et al. 2002) or the green fluorescent protein (GFP) gene. All our experiments were performed as suggested by the Ethics in Animal Use Committee of the Rio Grande Federal University, where this work was carried out.
Median lethal salinity for non-transgenic zebrafish
Median lethal salinity (MLS) for NT adult zebrafish was determined by transferring groups of adult male fish (30 fish for each group) from salinity 0 (control) to 2.5, 5, 7.5, 10, 12.5 and 15 parts per thousand (ppt) for 96 h. Salinity of 0 ppt (control) represent values lower than 1 ppt. Experimental salinities were prepared by mixing dechlorinated tapwater with filtered seawater and daily monitored with a hand refractometer. Each experimental salinity was tested in triplicate using circular 3-l plastic tanks containing 30 fish in each tank. The experimental media was continuously aerated. Water salinity, pH (7.51 ± 0.34), nitrate concentration (non-detected) and temperature (27 ± 1 °C) were monitored daily. Fish were fed twice a day with commercial ration (ColorBits, Tetra). Mortality was monitored every 24 h. Death criteria adopted were lack of both opercular movement and twitching reflex upon prodding with a needle. MLS and 95 % confidence interval (CI 95 %) were estimated using the Trimmed–Spearman–Karber method described by Hamilton et al. (1977).
Growth rate of non-transgenic zebrafish at different salinities
Growth rates of non-transgenic zebrafish were determined at different salinities. Groups of twenty (5 week-old) fish were exposed to salinities of 0 (control), 5 and 7.5 ppt for 15 days. Experimental salinities were prepared and monitored as described for the MLS test. Each experimental salinity was tested in triplicate using circular 3-l plastic tanks containing 20 fish in each. The experimental media were continuously aerated. Temperature (27 ± 1 °C) and photoperiod (14:10-h light–dark) were fixed. Fish were fed twice a day with commercial ration (ColorBits, Tetra). Food provided corresponded to 5 % of the mean fish mass in the respective tank. Growth rate at each experimental salinity was determined by measuring fish body mass at 7-day intervals. For measurement of growth, fish were anesthetized with tricaine methanesulfonate (0.1 mg/ml), weighed, and returned to the respective tank. Endpoints calculated were relative growth rate (RGR) and instantaneous growth rate (G). RGR was expressed as the percentage in body weight (W) increase after 15 days, while G represented the exponential growth rate. These endpoints were calculated using the equations RGR = (Wfinal − Winitial) × (Winitial)−1 × 100 and G = [ln(Wfinal − Winitial) × 15−1].
Salinity tolerance of non-transgenic and GH-transgenic zebrafish
Salinity tolerance of NT and T zebrafish was tested using the MLS method described above. T zebrafish males from F0104 lineage were used for this test. These fish showed a 2.6-fold higher body mass than the NT fish at the same age, demonstrating that the cβA/msGH transgene produces an active hormone (Figueiredo et al. 2007a). NT and T zebrafish males of same age (adults) were exposed to 11 ppt salinity for 96 h and mortality rate was checked every 12 h. Experiments were performed in triplicate (n = 10 fish per replicate) using the experimental conditions previously described for the growth rate experiment.
Expression of osmoregulation-related genes
Expression of osmoregulation-related genes [Na+, K+ATPase alpha 1 (NKA), H+-ATPase (HATPAse), plasmatic carbonic anhydrase (CA1), cytosolic carbonic anhydrase (CA2) and Na+/H+ exchange 3b (NHE3b)] was determined in gills of adult male control NT and control T zebrafish (n = 6). Control means raised at 0 ppt salinity. Prior to tissue dissection, adult zebrafish were anesthetized with buffered tricaine (MS222, 0.2 mg ml−1). Gills were dissected and washed with deionized water, dried on filter paper, transferred to plastic assay tubes, and frozen (−80 °C) for subsequent analysis of gene expression. Isolated gills were mixed and homogenized in Trizol reagent (Invitrogen, Brazil). Total RNA was purified with RNAse-free DNAse I (Applied Biosystems, Brazil) following the manufacturer’s protocol. The total amount of RNA was determined with a Qubit Fluorometer and a Quant-iT RNA BR Assay Kit (Invitrogen, Brazil). For cDNA synthesis, 5 μg of total RNA were reverse-transcribed using a High-Capacity cDNA Reverse Transcription Kit (Applied Biosystems, Brazil). The obtained cDNA was used as a template for gene amplification using specific primers designed based on specific gene sequences available at GenBank (www.ncbi.nlm.nih.gov). The source sequences and designed primers are shown in Table 1. Primers were designed using Primer Express 2.0 software (Applied Biosystems, Brazil). The total amount of cDNA also was determined with a Qubit Fluorometer and Quant-iT ssDNA Assay Kit (Invitrogen, Brazil). Quantitative PCR was performed with an ABI 7500 System (Applied Biosystems, Brazil) using Platinum SYBR Green qPCR SuperMix-UDG (Invitrogen, Brazil). Quantitative real-time PCRs with cDNA serial dilutions was performed for all primers in order to test reaction efficiency. The thermocycling program used was 40 cycles of 95 °C for 15 s and 60 °C for 30 s, with an initial cycle of 50 °C for 2 min and 95 °C for 2 min cDNA (0.8 μL) was used for Q-PCR reaction in a total volume of 10 μL. Triplicates were run for each sample. Expression levels of target genes were normalized using the expression of a housekeeping gene, elongation factor 1a (EF1a). mRNA abundance of EF1a relative to total RNA did not exhibit any significant difference among experimental groups.
Effect of salinity on the expression of genes related to osmoregulation, somatotropic axis and energy metabolism
The effect of median lethal salinity (MLS) on expression of zebrafish genes related to osmoregulation (NKA alpha1, HATPase, CA1, CA2, and NHE3b), somatotropic axis (GHR and IGF1) and energy metabolism [citrate synthase (CS) and lactate dehydrogenase (LDH)] was evaluated in NT and T zebrafish. Adult males (n = 6 for each fish group) were exposed to the MLS (11 ppt) for 24 h. Prior to tissue dissection, adult zebrafish were anesthetized with buffered tricaine methanesulfonate (MS222, 0.2 mg ml−1). Gills were dissected, washed with deionized water, dried on filter paper, transferred to plastic assay tubes, and frozen (−80 °C) for subsequent gene expression analysis, ATP concentration and Na+, K+-ATPase activity. The liver was dissected, washed with deionized water, transferred to plastic assay tubes, and frozen (−80 °C) for analysis of energy metabolism-related gene expression. Gene expression analyses were performed using the same protocol described above. Primers used for energy metabolism-related genes are shown in Table 1.
Gill Na+, K−-ATPase activity
Gill enzyme activity was analyzed in NT and T zebrafish (n = 6 for each group). For the enzyme activity assay, samples were thawed and kept on ice throughout the analysis. Tissue was homogenized in 100 μl of ice-cold buffer solution (300 mM sucrose, 20 mM ethylenediaminetetraacetic acid, 100 mM imidazole, and 0.1 % sodium deoxycholate), and centrifuged at 5,000g for 30 s at 4 °C. Enzyme activity was measured in the supernatant using the method described by Wheatly and Henry (1987) with modifications according to McCormick (1993). Two reaction mixtures were assayed. Reaction mixture A consisted of 20 ml of sample, 50 ml of salt solution A, and 150 ml of working solution A. Reaction mixture B consisted of 20 ml of sample, 50 ml of salt solution B, and 150 ml of working solution B. Salt solution A contained 100 mM NaCl, 10.5 mM MgCl2, 30 mM KCl, and 50 mM imidazole, with pH adjusted to 7.5. In salt solution B, NaCl replaced KCl at the same concentration. Working solution A contained 5 enzymatic units ml−1 lactate dehydrogenase, 5 enzymatic units ml−1 pyruvate kinase, 2.8 mM phosphoenolpyruvate, 3.5 mM adenosine triphosphate (ATP), 0.22 mM nicotinamide adenine dinucleotide phosphate, and 50 mM imidazole, with pH adjusted to 7.5. To obtain working solution B, 1 mM ouabain was added to working solution A. Kinetic assays then were run in duplicate at 25 °C in a temperature-controlled microplate reader (Molecular Devices, Menlo Park, CA, USA), for 10 min. An adenosine diphosphate standard curve (0–20 n moles/10 ml) also was run. Na+, K+-ATPase activity was then calculated by considering the difference in adenosine diphosphate (ADP) production between the two reaction mixtures (A and B). Protein content in the homogenate was measured with Quant-iT™ dsDNA BR Assay Kits (Invitrogen, Brazil) following the manufacturer’s protocol. Enzyme activity was then expressed as μmolesADP mg protein−1 h−1.
Gill ATP concentration
Gills of NT and T zebrafish exposed to median lethal salinity (MLS) were sonicated for 30 min in sterile water and centrifuged at 5,000 rpm for 1 min at 4 °C (n = 6 for each group). Tissue ATP concentration was measured in the supernatant using a luciferin-luciferase ATP Bioluminescent Assay Kit (Sigma-Aldrich, Brazil) following the manufacturer’s protocol. ATP concentration was quantified in a luminometer (Victor 2, Perkin Elmer, Brazil). Protein content in the supernatant was analyzed using the Quant-iT™ dsDNA BR Assay Kit (Invitrogen, Brazil) following the manufacturer’s protocol. Protein content data were used to normalize the measured luminescence (ATP content).
Determination of whole body Na+ concentration
Adult males from T and NT fish (n = 6 for each group) were exposed to the MLS (11 ppt salinity) for 24 h. Prior to analysis, adult zebrafish were anesthetized with buffered tricaine (MS222, 0.2 mg ml−1). For determination of whole-body sodium concentration, fish from freshwater (control) and 11 ppt salinity were anesthetized in tricaine methanesulfonate, rinsed in deionised water, blotted dry, digested in 1.5 ml 65 % HNO3 (Suprapur, Merck) and diluted. Plasma Na+ was analyzed by flame photometry (Corning 480 FlamePhotometer) and tissue sodium concentrations are represented as mmol Na+ per wet weight of tissue (mmol Na+/Kg wet weight).
Statistical analyses
Mortality and growth rate data were compared through analysis of variance (ANOVA) followed by Tukey’s test. Significant differences in gene expression data were assessed using the Relative Expression Software Tool—REST (Pfaffl et al. 2002). ATP concentration and Na+, K+-ATPase activity data for NT and T zebrafish were compared using paired t tests. Normality and homogeneity of variances were previously verified. In all analyses, a significant difference was inferred when P < 0.05.
Results
No mortality was observed in NT zebrafish kept of salinities of 0 (control), 2.5, 5 and 7.5 ppt for 96 h. However, 6.7 (±7.7), 70 (±13.5) and 100 % mortality was observed in those kept in salinities of 10, 12.5, and 15 ppt, respectively. The MLS for NT zebrafish was calculated as 11.6 ppt (CI 95 % = 11.2–12.2). Percentage mortality of NT male zebrafish was lower than that of T male zebrafish over the 96 h of exposure to the MLS. After 96 h of exposure, mortality of NT and T zebrafish was 64 % (±19) and 100 %, respectively (Fig. 1).

Percentage mortality of non-transgenic (control; NT) and GH-transgenic (T) zebrafish exposed to salinity of 11 ppt over 96 h. Data are mean ± SEM (n = 10 for each replicate; n = 3 replicates). Asterisks indicate significant different mean values between NT and T zebrafish (*P < 0.05)
Growth rate of NT zebrafish exposed to 5 ppt salinity was 1.8-fold higher than in those kept in 0 ppt salinity (control). However, growth rate of zebrafish reared in salinity of 7.5 ppt was not significantly different from that of control zebrafish (Table 2).
Comparison of gene expression in NT and T zebrafish (NT × T) under control conditions showed that the NKA, HATPase, CA1 and CA2 genes were up-regulated (2.4, 8.5, 3.2 and 9.3-fold induction, respectively), in T zebrafish, while NHE3b gene expression did not show significant differences between the fish groups (Fig. 2).

Relative expression of osmoregulation-related genes in gills of non-transgenic (control; NT) and GH-transgenic (T) zebrafish kept under control condition (salinity of 0 ppt). The horizontal line represents the relative expression of NT zebrafish. The expression level of each gene was normalized by comparison with the expression of the elongation factor 1 alpha (ef1a) gene. Data are median ± SEM (n = 6). Significant differences (P < 0.05) in gene expression levels are denoted by an asterisk. NKA Na+/K+-ATPase gene; H + -ATPase H+-ATPase gene, CA1 carbonic anhydrase 1 gene, CA2 carbonic anhydrase 2 gene, and NHE3b Na+/H+ exchanger 3b gene
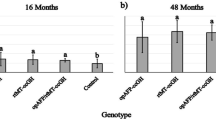
The effect of 11 ppt salinity on expression of zebrafish genes related to the somatotropic axis (GHR and IGF1) was evaluated in NT and T zebrafish. The GHR gene was significantly down-regulated in gills and liver of NT zebrafish exposed to 11 ppt salinity (86 and 79.4 % inhibition, respectively). While IGF1 was down-regulated in liver (89.3 % inhibition) of NT exposed to 11 ppt salinity (Fig. 3a). For T zebrafish, transfer to 11 ppt salinity caused a down-regulation of GHR in gills and liver (67.8 and 88.9 % inhibition, respectively). IGF1 was down-regulated (84.1 % inhibition) in gills of T exposed to 11 ppt salinity (Fig. 3b).

Relative expression of somatotropic axis genes in gills and liver of a non-transgenic (NT) and b GH-transgenic (T) zebrafish exposed to 11 ppt salinity for 24 h. The horizontal line represents the relative expression of fish kept under control condition (salinity of 0 ppt). The expression level of each gene was normalized by comparison with the expression of the elongation factor 1 alpha (ef1a) gene. Data are median ± SEM (n = 6). Significant difference (P < 0.05) between zebrafish kept under control condition (salinity 0; NT0 or T0) and those exposed to 11 ppt salinity (NT11 or T11) is denoted by an asterisk. IGF1 insulin-like growth factor-1 and GHR growth hormone receptor
For NT zebrafish, comparison of gene expression between those kept under control conditions (salinity of 0 ppt) and those exposed to 11 ppt salinity (NT0 × NT11) showed that H+-ATPase, CA1, and CA2 genes were up-regulated (2.2, 6.3 and 4.7-fold induction, respectively) in gills of zebrafish exposed to 11 ppt salinity (Fig. 4a). However, the expression of NKA and NHE3b genes was not significantly different in the gills of the two fish groups. Regarding energy metabolism-related genes, CS gene expression was not significantly different between zebrafish kept under control condition (0 ppt salinity) and those exposed to 11 ppt salinity (NT0 × NT11) for both liver and gills (Fig. 5a). Furthermore, LDH expression did not change in liver of control and salinity-exposed fish. However, the expression of this gene was significantly lower (97 % inhibition) in gills of zebrafish exposed to 11 ppt salinity than in those kept under the control condition (salinity of 0 ppt).

Relative expression of osmoregulation-related genes in gills of a non-transgenic (NT) and b GH-transgenic (T) zebrafish exposed to 11 ppt salinity for 24 h. The horizontal line represents the relative expression of fish kept under control condition (salinity 0). The expression level of each gene was normalized by comparison with the expression of the elongation factor 1 alpha (ef1a) gene. Data are median ± SEM (n = 6). Significant difference (P < 0.05) between zebrafish kept under control condition (salinity 0; NT0 or T0) and those exposed to 11 ppt salinity (NT11 or T11) is denoted by an asterisk. NKA Na+/K+-ATPase gene, H + -ATPase H+-ATPase gene, CA1 carbonic anhydrase 1 gene; CA2 carbonic anhydrase 2 gene, and NHE3b Na+/H+ exchanger 3b gene

Relative expression of energy metabolism-related genes in gills and liver of a non-transgenic (NT) and b GH-transgenic (T) zebrafish exposed to 11 ppt salinity for 24 h. The horizontal line represents the relative expression of fish kept under control conditions (salinity 0). The expression level of each gene was normalized by comparison with the expression of the elongation factor 1 alpha (ef1a) gene. Data are median ± SEM (n = 6). Significant difference (P < 0.05) between zebrafish kept under control condition (salinity 0; NT0 or T0) and those exposed to 11 ppt salinity (NT11 or T11) is denoted by an asterisk. LDH lactate dehydrogenase gene; and CS citrate synthase gene
For T zebrafish, comparison between those kept under control condition (salinity of 0 ppt) and those transferred to 11 ppt salinity (T0 × T11) showed that all osmoregulation-related genes were down-regulated (57, 68, 58, 73 and 88 % inhibition for NKA, HATPase, CA1, CA2 and NHE3b, respectively) in gills of T zebrafish exposed to 11 ppt salinity (Fig. 4b). In contrast, LDH gene expression was up-regulated (5.9-fold) in the liver of T zebrafish exposed to salinity of 11 ppt, whereas CS gene expression was down-regulated (40 % inhibition) in gills of these fish (Fig. 5b).
Na+, K+-ATPase activity was significantly higher in gills of T zebrafish exposed to 11 ppt salinity when compared with other treatments (Fig. 6), and fishes from this group (T zebrafish) also showed the highest whole-body content of Na+ (Fig. 7). However, concentration of Na+ in whole body of NT zebrafish exposed to 11 ppt salinity, was higher than for animals kept under control conditions (T and NT zebrafish at salinity of 0 ppt) (Fig. 7).

Na+, K+-ATPase activity in gills of non-transgenic (NT) and GH-transgenic (T) zebrafish exposed to 11 ppt salinity for 24 h. Data are mean ± SEM (n = 6). Different letters denoting significant difference between zebrafish kept under control condition (salinity 0; NT0 or T0) and those exposed to 11 ppt salinity (NT11 or T11) (P < 0.05)

Whole-body sodium concentration in non-transgenic (NT0) and GH-transgenic (T0) zebrafish exposed to 0 ppt salinity (control) and non-transgenic (NT11) and GH-transgenic (T11) zebrafish exposed to 11 ppt salinity for 24 h (mmol kg−1). Data are mean ± SEM (n = 6–9). Different letters denote significant differences among treatment groups (P < 0.05)
For both NT and T zebrafish, gill ATP content increased when fish were exposed to 11 ppt salinity. However, ATP content was 9.5-fold higher in gills of T zebrafish when compared to control fish kept at salinity of 0 ppt, while it was only threefold higher in gills of NT zebrafish when compared to their respective control (Fig. 8).

ATP concentration in gills of a non-transgenic (NT) and b GH-transgenic (T) zebrafish exposed to 11 ppt salinity for 24 h. Data are mean ± SEM (n = 6). Significant difference (P < 0.05) between zebrafish kept under control condition (salinity 0; NT0 or T0) and those exposed to 11 ppt salinity (NT11 or T11) is denoted by an asterisk
Discussion
In teleost fishes, growth hormone (GH) has been shown to have a significant involvement in osmoregulation, in addition to its growth-promoting role. In the present study, we evaluated the effect of GH transgenesis on osmoregulation and energy metabolism in zebrafish, a stenohaline freshwater fish. Even though the choice of this species could seem counterintuitive for osmoregulation studies since it copes poorly with increasing salinities, our objective was to verify whether GH transgenesis could change the species’ salinity tolerance.
The growth rate of non-transgenic zebrafish exposed to intermediate salinities was evaluated in order to verify whether its growth rate was affected by this environmental parameter. Results from the growth experiment showed that zebrafish exposed to 5 ppt salinity had higher growth rates than those kept in salinity of 0 ppt (control), while the growth rate of fish reared in 7.5 ppt salinity was not different from that of control zebrafish. This finding clearly indicates that moderate increases in salinity can induce growth in zebrafish. Indeed, in isosmotic condition the considerable costs of osmoregulation are minimized, freeing up energy for growth. In fact, several studies with croaker (Peterson et al. 1999), tilapia (Jonassen et al. 1997) and sea bream (Laiz-Carrión et al. 2005) showed that better growth occurs at intermediate salinities. Moreover, acclimation to conditions of isosmotic salinity resulted in significantly higher GH gene expression in euryhaline species, as silver sea bream (Deane and Woo 2006), black sea bream (Deane and Woo 2005), as well as in stenohaline fish as channel catfish Ictalurus punctatus (Tang et al. 2001; Drennon et al. 2003). While the induction of GH expression during acclimation to conditions of isosmotic salinity has been shown for some species, this phenomenon has yet to be examined in a wider sampling of marine and especially freshwater fishes.
The molecular mechanism of the salinity/GH relationship was evaluated by gene expression analysis. First, we analyzed the expression of osmoregulation-related genes in NT and T zebrafish and the results showed that the excessive release of GH in T zebrafish promotes an up-regulation of the Na+K+ATPase, H+-ATPase, plasma carbonic anhydrase and cytosolic carbonic anhydrase genes, without changes in the expression of the Na+/H+ exchange 3b gene. These findings are in agreement with the observation that GH and IGF1 stimulate gill chloride cells in number and/or size in several fish species (for review, see McCormick 2001). Furthermore, immunocytochemical studies indicate that hormone-induced increases in Na+, K+-ATPase activity take place in chloride cells (Seidelin et al. 1999).
As previously mentioned, two pathways have been proposed for the apical transport of Na+ in zebrafish gill cells: an apical V-type H+-ATPase, which is electrically linked to the Na+ absorption via the epithelial Na+ channel, and an electroneutral exchange of Na+ and H+ via the apical Na+/H+ exchanger (Hirose et al. 2003; Perry et al. 2003; Evans et al. 2005). Thus, the increased expression of the H+-ATPase gene observed in T zebrafish in the present study suggests that GH may also influence the Na+ uptake via the apical Na+ channel in addition to the stimulating the increasing number of chloride cells and consequently NKA expression.
Even though zebrafish gill ionocytes are responsible for Na+ uptake mechanisms, it is possible that GH transgenesis works blocking Na+ import and/or promoting Na+ export under hyperosmotic conditions. To test the hypothesis that a GH-transgenic fish could modify the osmoregulatory mechanism, we first determined the MLS for NT zebrafish and then challenged GH-transgenic zebrafish to this salinity. Mortalities, 96 h post transfer to 11 ppt salinity were 63 and 100 % for NT and T fish, respectively. Despite the GH-transgenic zebrafish seeming to be more sensitive to salinity, this result suggests that both T and NT zebrafish are not adapted to live in hyperosmotic environment. Moreover, it would be expected that at this salinity (11 ppt), the elevation of plasma sodium concentration would increase the energetic cost for ionic and osmoregulation, thereby decreasing the amount of energy available for growth. Even though there is uncertainly on the actual energetic cost for osmotic regulation in fish, the classical approach based on direct measurements of O2 consumption indicates that as much as 20–68 % of the total energy budget is involved in the process of osmoregulation (Bouef and Payan 2001).
To clarify the lower ability of T zebrafish to cope with increasing salinity, the genes coding for the major proteins involved in ionic and osmotic regulation in aquatic animals as well as GHR and IGF1 and energy production (ATP) were investigated. All analyzed osmoregulatory and somatotropic-related genes were down-regulated in transgenic zebrafish exposed to 11 ppt salinity. Following transfer from freshwater to saltwater, the fish must reduce or block this active Na+ entry. In zebrafish gills, H+-ATPase-rich cells (HR) are responsible for Na+-uptake mechanism (For review, see Hwang 2009). Down-regulation of the NHE3b gene could be associated with attempts at reducing Na+ import, since NHE3b is the main exchanger involved with the regulation of internal Na+ balance (Boisen et al. 2003). Another adaptation could be the induction of HR cell death, explaining the parallel decrease of all HR genes seen in T fish.
Thus, GH transgenesis and acute salinity stress down-regulated osmoregulatory genes in gills of GH-transgenic zebrafish but increased Na+, K+-ATPase activity. The Na+, K+-ATPase activity is closely associated with the osmoregulatory capacity of gills in eliminating the excess of ions in a hyperosmotic environment (Marshall 2002). For example, Mancera and McCormick (2000) showed a very rapid and transitory increase in gill Na+, K+-ATPase activity in the first hours after the transfer of euryhaline Fundulus heteroclitus to seawater, which is dependent on translational and transcriptional processes. Therefore, the higher gill Na+, K+-ATPase activity in GH-transgenic zebrafish with increasing salinity observed in the present study suggests that this pump is activated/stimulated by a combination of increasing salinity and GH, since the same significant increase was not observed in NT zebrafish subjected to 11 ppt salinity.
On the other hand, in freshwater fishes Na+, K+-ATPase is responsible for Na+ active uptake and in this study, we showed a significant increase in Na+, K+-ATPase activity in gills of GH-transgenic fish. In order to verify whether this increase in Na+, K+-ATPase activity reflects an higher ability of Na+ import, we measured the whole-body Na+ concentration. T zebrafish as well as NT zebrafish exposed to 11 ppt showed an augment in the Na+ whole-body content when compared to the fishes kept under control condition (salinity of 0 ppt). These results indicate that both NT and T zebrafish at 11 ppt are in an unfavorable physiological situation and that the excess of GH due to transgenesis stimulates Na+, K+-ATPase activity, leading to an increase in Na+ uptake. Thus, no adaptative response was observed as a consequence of GH-transgenesis which could make this fish more tolerant to saltwater environments. Actually, the higher Na+ concentration in T zebrafish at 11 ppt could contribute to the higher mortality observed in this group. T zebrafish mortality also could be associated with other physiological processes that involve GH expression such as ammonia excretion and acid–base balance, but this is unclear to this time. In addition, it is important to keep in mind that saline stress naturally increases energy expenditure, with this fact much more impacting in a T fish with a high energy demand imposed by GH excess.
Rosa et al. (2008) reported a significant increase in oxygen consumption and production of reactive oxygen species (ROS) in GH-transgenic zebrafish from the F0104 lineage than in NT zebrafish. A similar response also was reported for GH-transgenic mice, where an increment in GH levels was associated with an increased metabolic demand (Carlson et al. 1999), increasing the mitochondrial ROS production (Muradian et al. 2002; Fridovich 2004). These observations indicate that GH excess has the potential of inducing oxidative stress in fishes and mammals, possibly compromising growth by imposing an additional energetic cost, which is not required when an adequate balance exists between energy available and circulating GH level. Leggatt et al. (2009) showed that a GH transgene significantly influences the metabolic enzyme capacity in coho salmon, suggesting that accelerated growth is not met simply by increasing feed intake, but also by shifting emphasis on energy production through modification of carbohydrate, amino acid, and lipid metabolism pathways.
Reorganization and alterations in intermediate metabolic pathways may occur in an attempt to meet the increases energetic demand associated with salinity acclimation (for review, see Tseng and Hwang 2008). Some fishes such as the rainbow trout Oncorhynchus mykiss seem to alter their metabolic pathways and use lactate as an energetic substrate during acclimation to changing salinities (Sangiao-Alvarellos et al. 2005). Lactate is synthesized by the conversion of pyruvate through the catalytic action of lactate dehydrogenase (LDH), as an end-product of glycolysis. In the present study, expression of the LDH gene was evaluated in both NT and T zebrafish. GH-transgenic zebrafish subjected to elevated salinity of 11 ppt showed increased expression of the LDH gene in the liver, but not in the gills. Tseng et al. (2008) suggested that the stimulation of glycolysis and Kreb’s cycle is associated with the increased Na+, K+-ATPase activity observed in mitochondria-rich cells in fish gills during acute acclimation to seawater. They also suggested that the lactate released by the liver during this process can be considered a secondary metabolite, since both lactate and glucose increased in the plasma of Spartus aurata immediately after transfer to seawater (Sangiao-Alvarellos et al. 2003). The fact that the LDH expression did not change significantly in the liver of NT zebrafish after abrupt transfer from freshwater to brackish water (salinity of 11 ppt) is in agreement with findings reported for S. aurata. In turn, the reduced LDH expression observed in gills of NT zebrafish exposed to brackish water indicates that increasing salinity may inhibit LDH mRNA synthesis in fish that are not being stressed by continuous growth. On the other hand, increased expression of the LDH gene in liver of GH-transgenic zebrafish supports the explanation that these fish are using the anaerobic metabolism to meet the additional energy requirements imposed by the increasing environmental salinity.
Energy metabolism also was analyzed in both NT and T zebrafish through the expression of citrate synthase (CS), which is the first enzyme involved in the Krebs’ cycle. This enzyme catalyzes the condensation reaction of acetyl-CoA molecules and oxaloacetate to form citrate. In T zebrafish, CS mRNA synthesis decreased under saline stress, indicating a possible instant increase of aerobic metabolism in gills, since the largest inhibitor of CS expression is the tissue ATP concentration. In contrast, no alteration in CS gene expression was found in the liver of both NT and T zebrafish. For a better understanding of these findings, ATP concentration was determined in gills of zebrafish kept under control conditions (freshwater) or exposed to saline stress. The increased ATP content observed in gills of T zebrafish subjected to 11 ppt salinity in fact indicate that a higher amount of ATP is being redirected to the gills during the saline stress, which would be inhibiting gill CS mRNA synthesis. Furthermore, increased activity of gill Na+, K+-ATPase, an ATP-dependent enzyme, also was observed in T zebrafish exposed to 11 ppt salinity. It is important to note that a similar response was not observed in the NT zebrafish, since no significant change in CS expression and Na+, K+-ATPase activity was observed in gills of these fish after the abrupt transfer from freshwater to 11 ppt salinity.
Regarding salinity tolerance, results from the present study show that GH-transgenic zebrafish are more sensitive to increased salinity than NT zebrafish. This reduced tolerance to environmental salinity is likely because available energy is being used to support the continuous growth and higher Na+ uptake induced by the GH excess, which leads to increased demand for oxygen to supply the aerobic pathways that support the growth enhancement (Cook et al. 2000; Lee et al. 2003). Another factor that should be considered is the structure of the transgene used for transgenic fish production. Normally, transgenes are designed for inducing over-expression of the gene of interest through promoters that are characterized by strong and constitutive expression, in which regulation is independent of the normal regulation of the target gene. For example, in the present study, a transgene comprised of the carp β-actin promoter driving the expression of the GH gene from the marine silverside was employed. Thus, the GH-transgenic zebrafish are continuously producing this hormone and consequently suffering all its physiological effects. Under this situation, fish should activate the negative feedback system to decrease the amount of circulating GH that is pushing continuous protein synthesis, thus avoiding excessive energy loss (Figueiredo et al. 2007b).
Taking altogether, data reported here study clearly indicate that GH-transgenic zebrafish are continuously facing an energetic imbalance, and GH excess could increase Na+ import, leading to higher mortality of GH-transgenic zebrafish in a hyperosmotic environment. Thus, we can infer that GH excess caused by transgenesis promotes an unfavorable osmotic and energetic physiological status in GH-transgenic zebrafish.
References
Ayoola SO, Idowu AA (2008) Biotechnology and species development in aquaculture. Afr J Biotechnol 7(25):4722–4725
Boisen A, Amstrup J, Novak I, Grosell M (2003) Sodium and chloride transport in soft water and hard water acclimated zebrafish (Danio rerio). Biochim Biophys Acta 1618:207–218
Bolton J, Kawauchi H, Hirano T (1987) Osmoregulatory actions of growth hormone in rainbow trout (Salmo gairdneri). J Endocrinol 112:63–68
Bouef G, Payan P (2001) How should salinity in influence fish growth? Comp Biochem Physiol C 130:411–423
Breton S (2001) The cellular physiology of carbonic anhydrases. J Pancreas 2:159–164
Carlson L, Bharadwaj R, Bartlke A (1999) Oxidative stress in hypopituitary dwarf mice and in transgenic mice overexpressing human and bovine GH. Age 22:181–186
Cook JT, McNiven MA, Sutterlin AM (2000) Metabolic rate of pre-smolt growth-enhanced transgenic Atlantic salmon Salmo salar. Aquaculture 188:33–45
Craig PM, Wood CM, McClelland GB (2007) Gill membrane remodeling with soft-water acclimation in zebrafish (Danio rerio). Physiol Genomics 30(1):53–60
Deane EE, Woo NY (2005) Cloning and characterization of sea bream Na+K+-ATPase α and β subunit genes: in vitro effects of hormones on transcriptional and translational expression. Biochem Biophys Res Commun 331:1229–1238
Deane EE, Woo NY (2006) Molecular cloning of growth hormone from silver sea bream: effects of abiotic and biotic stress on transcriptional and translational expression. Biochem Biophys Res Commun 342:1077–1082
Devlin RH, Yesaki TY, Biagi CA, Donaldson EM, Swanson P, Chan WK (1994) Extraordinary growth in salmon. Nature 371:209–210
Devlin RH, Yesaki TY, Donaldson EM, Shao JD, Hew C-L (1999) Production of germline transgenic Pacific salmonids with dramatically increased growth performance. Can J Fish Aquat Sci 52:1376–1384
Drennon K, Moriyama S, Kawauchi H, Small B, Silverstein J, Parhar I, Shepherd B (2003) Development of an enzyme-linked immunosorbent assay for the measurement of plasma growth hormone (GH) levels in channel catfish (Ictalurus punctatus): assessment of environmental salinity and GH secretogogues on plasma GH levels. Gen Comp Endocrinol 133(3):314–322
Du SJ, Gong Z, Fletcher GL, Shears MA, King MJ, Idler DR, Hew CL (1992) Growth enhancement in transgenic Atlantic salmon by the use of an ‘all fish’ chimeric growth hormone construct. Biotechnology 10:176–181
Eppler E, Berishvili G, Mazel P, Caelers A, Hwang G, Maclean N, Reinecke M (2010) Distinct organ-specific up- and down-regulation of IGF-I and IGF-II mRNA in various organs of a GH-overexpressing transgenic Nile tilapia. Transgenic Res 19:231–240
Evans DH, Piermarini PM, Potts W (1999) Ionic transport in the fish gill epithelium. J Exp Zool 283:641–652
Evans DH, Piermarini PM, Choe KP (2005) The multifunctional fish gill: dominant site of gas exchange, osmoregulation, acid–base regulation, and excretion of nitrogenous waste. Physiol Rev 85:97–177
Figueiredo MD, Lanes CF, Almeida DV, Marins LF (2007a) Improving the production of transgenic fish germlines: in vivo evaluation of mosaicism in zebrafish (Danio rerio) using a green fluorescent protein (GFP) and growth hormone cDNA transgene co-injection strategy. Genet Mol Biol 30:31–36
Figueiredo MD, Lanes CF, Almeida DV, Proietti MC, Marins LF (2007b) The effect of GH overexpression on GHR and IGF-I gene regulation in different genotypes of GH-transgenic zebrafish. Comp Biochem Physiol Part D 2:228–233
Fridovich I (2004) Mitochondria: are they the seat of senescence? Aging Cell 3:13–16
Guan B, Hu W, Zhang T, Wang Y, Zhu Z (2008) Metabolism traits of “all-fish” growth hormone transgenic common carp (Cyprinus carpio L.). Aquaculture 284:217–223
Hallerman EM, McLean E, Fleming IA (2007) Effects of growth hormone transgenes on the behavior and welfare of aquacultured fishes: a review identifying research needs. Appl Animal Behav Sci 104:265–294
Hamilton MA, Russo RC, Thurston RV (1977) Trimmed Spearman-Karber method for estimating median lethal concentrations in toxicity bioassays. Environ Sci Technol 11:714–719
Higgs DA, Sutton JN, Kim H, Oakes JD, Smith J, Biagi C, Rowshandeli M, Devlin RH (2009) Influence of dietary concentrations of protein, lipid and carbohydrate on growth, protein and energy utilization, body composition, and plasma titres of growth hormone and insulin-like growth factor-1 in non-transgenic and growth hormone transgenic coho salmon, Oncorhynchus kisutch (Walbaum). Aquaculture 286:127–137
Hirose S, Kaneko T, Naito N, Takei Y (2003) Molecular biology of major components of chloride cells. Comp Biochem Physiol Part B Biochem Mol Biol 136:593–620
Hoshijima K, Hirose S (2007) Expression of endocrine genes in zebrafish larvae in response to environmental salinity. J Endocrinol 193(3):481–491
Hwang PP (2009) Ion uptake and acid secretion in zebrafish (Danio rerio). J Exp Biol 212:1745–1752
Jonassen TM, Pittman K, Imsland AK (1997) Seawater acclimation of tilapia, Oreochromis spilurus spilurus Günter, fry and fingerlings. Aquac Res 28:205–214
Laiz-Carrión R, Sangiao-Alvarellos S, Guzmán JM, Río MP, Soengas JL, Mancera JM (2005) Growth performance of gilthead sea bream Sparus aurata in different osmotic conditions: implications for osmoregulation and energy metabolism. Aquaculture 250:849–861
Lee CG, Devlin RH, Farrell AP (2003) Swimming performance, oxygen consumption and excess post-exercise oxygen consumption in adult transgenic and ocean-ranched coho salmon. J Fish Biol 62:753–766
Leggatt RA, Devlin RH, Farrell AP, Randall DJ (2003) Oxygen uptake of growth hormone transgenic coho salmon during starvation and feeding. J Fish Biol 62:1053–1066
Leggatt RA, Raven PA, Mommsen TP, Sakhrani D, Higgs D, Devlin RH (2009) Growth hormone transgenesis influences carbohydrate, lipid and protein metabolism capacity for energy production in coho salmon (Oncorhynchus kisutch). Comp Biochem Physiol Part B 154:121–133
Lindskog S (1997) Structure and mechanism of carbonic anhydrase. Pharmacol Ther 74:1–20
Mancera JM, McCormick SD (1998) Evidence for growth hormone/insulin-like growth factor I axis regulation of seawater acclimation in the euryhaline teleost Fundulus heteroclitus. Gen Comp Endocrinol 111:103–112
Mancera JM, McCormick SD (1999) Influence of cortisol, growth hormone, insulin-like growth factor I and 3,3,5-triiodo-L-thyronine on hypoosmoregulatory ability in the euryhaline teleost Fundulus heteroclitus. Fish Physiol Biochem 21:25–33
Mancera JM, McCormick SD (2000) Rapid activation of gill Na+, K+-ATPase in the euryhaline teleost Fundulus heteroclitus. J Exp Zool 287:263–274
Marins LF, Iyengar A, Maclean N, Levy JA, Sohm F (2002) Simultaneous overexpression of GH and STAT5b genes inhibits the STAT5 signalling pathway in tilapia (Oreochromis niloticus) embryos. Genet Mol Biol 25:293–298
Marshall WS (2002) Na+, Cl−, Ca2+ and Zn2+ transport by fish gills: retrospective review and prospective synthesis. Transport 283:264–283
McCormick SD (1993) Methods for nonlethal gill biopsy and measurement of Na+, K+-ATPase activity. Can J Fish Aquat Sci 50:656–658
McCormick SD (1996) Effects of growth hormone and insulin-like growth factor I on salinity tolerance and gill Na+ , K+-ATPase in Atlantic salmon (Salmo salar): interaction with cortisol. Gen Comp Endocrinol 101:3–11
McCormick SD (2001) Endocrine control of osmoregulation in teleost fish. Am Zool 41:781–794
Muradian KK, Utko NA, Mozzhukhina TG, Litoshenko AY, Pishel IN, Bezrukov VV, Fraifield VE (2002) Pair-wise linear and 3d nonlinear relationships between the liver antioxidant enzyme activities and the rate of body oxygen consumption in mice. Free Radical Biol Med 33:1736–1739
Nam YK, Noh JK, Cho YS, Cho HJ, Cho KN, Kim CG, Kim DS (2001) Dramatically accelerated growth and extraordinary gigantism of transgenic mud loach Misgurnus mizolepis. Transgenic Res 10:353–362
Parks SK, Tresguerres M, Goss GG (2008) Review of theoretical considerations underlying Na+ uptake mechanisms in freshwater fishes. Comp Biochem Physiol Part C 148:411–418
Perry SF, Shahsavarani A, Georgalis T, Bayaa M, Furimsky M, Thomas SL (2003) Channels, pumps, and exchangers in the gill and kidney of freshwater fishes: their role in ionic and acid–base regulation. J Exp Zool Part A Comp Exp Biol 300:53–62
Peterson MS, Comyns BH, Rakocinski CF, Fulling GL (1999) Does salinity affect somatic growth in early juvenile Atlantic croaker, Micropogonias undulatus (L.)? J Exp Mar Biol Ecol 238:199–207
Pfaffl MW, Horgan GW, Dempfle L (2002) Relative expression software tool (REST©) for group-wise comparison and statistical analysis of relative expression results in real-time PCR. Nucleic Acids Res 30:e36
Pitkänen TI, Krasnov A, Teerijoki H, Mölsä H (1999) Transfer of growth hormone (GH) transgenes into Arctic charr (Salvelinus alpinus L.) I. Growth response to various GH constructs. Genet Anal Biomol Eng 15:91–98
Rahman MA, Mak R, Ayad H, Smith A, Maclean N (1998) Expression of a novel piscine growth hormone gene results in growth enhancement in transgenic tilapia (Oreochromis niloticus). Transg Res 7:357–369
Reinecke M (2010) Influences of the environment on the endocrine and paracrine fish growth hormone–insulin-like growth factor-I system. J Fish Biol 76:1233–1254
Rosa CE, Figueiredo MA, Almeida DV, Marins LF (2008) Metabolic rate and reactive oxygen species production in different genotypes of GH-transgenic zebrafish. Comparative Biochemistry and Physiology—A. Mol Integr Physiol 149:209–214
Sakamoto T, Hirano T (1993) Expression of insulin-like growth factor I gene in osmoregulatory organs during seawater adaptation of the salmonid fish: possible mode of osmoregulatory action of growth hormone. Proc Natl Acad Sci USA 90:1912–1916
Sakamoto T, McCormick SD (2006) Prolactin and growth hormone in fish osmoregulation. Gen Comp Endocrinol 147:24–30
Sakamoto T, Shepherd BS, Madsen SS, Nishioka RS, Siharath K, Richman NH, Bern HA, Grau EG (1997) Osmoregulatory actions of growth hormone and prolactin in an advanced teleost. Gen Comp Endocrinol 106:95–101
Sangiao-Alvarellos S, Laiz-Carrión R, Guzmán JM, Martín Del Río MP, Miguez JM, Mancera JM, Soengas JL (2003) Acclimation of S. aurata to various salinities alters energy metabolism of osmoregulatory and nonosmoregulatory organs. Am J Physiol Reg Integr Comp Physiol 285:897–907
Sangiao-Alvarellos S, Míguez JM, Soengas JL (2005) Actions of growth hormone on carbohydrate metabolism and osmoregulation of rainbow trout (Oncorhynchus mykiss). Gen Comp Endocrinol 141:214–225
Seidelin M, Madsen SS, Byrialsen A, Kristiansen K (1999) Effects of insulin-like growth factor-I and cortisol on Na+ , K+-ATPase expression in osmoregulatory tissues of brown trout (Salmo trutta). Gen Comp Endocrinol 113:331–342
Shrimpton J, Devlin RH, Mclean E, Byatt CJ, Donaldson E, Randall D (1995) Increases in gill cytosolic corticosteroid receptor abundance and saltwater tolerance in juvenile coho salmon (Oncorhynchus kisutch) treated with growth hormone and placental lactogen. Gen Comp Endocrinol 98:1–15
Tang Y, Shepherd BS, Nichols AJ, Dunham R, Chen TT (2001) Influence of environmental salinity on messenger RNA levels of growth hormone, prolactin, and somatolactin in pituitary of the channel catfish (Ictalurus punctatus). Mar Biotechnol 3:205–217
Tseng Y, Hwang P (2008) Some insights into energy metabolism for osmoregulation in fish. Comp Biochem Physiol Part C 148:419–429
Tseng YC, Lee JR, Chang JCH, Kuo CH, Lee SJ, Hwang PP (2008) Regulation of lactate dehydrogenase in tilapia (Oreochromis mossambicus) gills during acclimation to salinity challenge. Zoolog Stud 47:473–480
Wheatly BM, Henry RP (1987) Branchial and antennal gland Na+/K+-dependent ATPase and carbonic anhydrase activity during salinity acclimation of the euryhaline crayfish Pacifastacus leniusculus. J Exp Biol 133:73–86
Xu B, Miao H, Zhang P, Li D (1997) Osmoregulatory actions of growth hormone in juvenile tilapia (Oreochromis niloticus). Fish Physiol Biochem 17:295–301
Yan JJ, Chou MY, Kaneko T, Hwang PP (2007) Gene expression of Na+/H+ exchanger in zebrafish H+ -ATPase-rich cells during acclimation to low-Na+ and acidic environments. Am J Physiol 293:1814–1823
Acknowledgments
The authors would like to thank Maíra Proietti for valuable corrections of the manuscript. This work was supported by Brazilian CNPq (Conselho Nacional de Desenvolvimento Científico e Tecnológico) and CAPES (Coordenação de Aperfeiçoamento de Pessoal de Nível Superior). AB is a research fellow from CNPq (Proc. # 304430/2009-9) and is supported by the International Canada Research Chair Program from the International Development Research Centre (Ottawa, Canada).
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Almeida, D.V., de Martinez Gaspar Martins, C., de Azevedo Figueiredo, M. et al. Growth hormone transgenesis affects osmoregulation and energy metabolism in zebrafish (Danio rerio). Transgenic Res 22, 75–88 (2013). https://doi.org/10.1007/s11248-012-9627-x
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11248-012-9627-x