
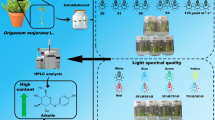
Abstract
Urtica dioica L. (Urticaceae), popularly known as nettle, is a medicinal plant used by the textile, food and pharmaceutical industry. The aim of the current study is to evaluate the influence of different LED wavelengths and light intensity on the growth, phytochemical content and antioxidant activity of Urtica dioica grown in vitro. Nodal segments were taken to MS culture medium under 26, 51, 69, 94 and 130 μmol m−2 s−1, and the following light spectra: blue, red, white, combinations of red and blue (1red/1blue, 2.5red/1blue and 1red/2.5blue) and cool white fluorescent lamps. Leaf area, photosynthetic pigments, total phenolics, total flavonoids and antioxidant activity were evaluated 40 days after culture implementation. Plantlets grown under 94 μmol m−2 s−1 showed better growth and dry weight production outcomes. Phenolic compound and flavonoid production, as well as antioxidant activity were intensified at 130 μmol m−2 s−1. Plantlets grown under 1red/2.5blue produced 5.53 times more phenolic compounds and 8.63 times more flavonoids than plants under fluorescent lights. And dry weight accumulation was favored by wavelength of 2.5red/1blue. The antioxidant activity was influenced by the light intensity, being directly proportional to the increase in light intensity. Increased red light rate induced plantlet etiolation.
Key message
Different spectra and light intensity—altered growth and photosynthetic pigments, total phenolics, total flavonoids and antioxidant activity in plantlet of Nettle.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Urtica dioica L. (Urticaceae), popularly known as nettle, is a medicinal plant rich in vitamins, iron, calcium, sodium and fatty acids (Rutto et al. 2013). It is used by the textile and food industries besides being common in popular medicine to treat arthritis, rheumatism, muscle paralysis and diabetes mellitus (Upton 2013). The species has attracted researchers’ attention for its anticancer and antioxidant activity (Fattahi et al. 2018; Ghaima et al. 2013; Gülçin et al. 2004; Mohammadi et al. 2016). Scions production based on conventional methods requires large spaces (Di Virgilio et al. 2015); on the other hand, micropropagation is a tool adopted to help the vegetative propagation of donor plants that present superior genetic characteristics. It demands smaller spaces for production at large scale throughout the year, as well as allows assessing environmental influence on plant growth and secondary metabolism (Amaral-Baroli et al. 2016; Gonçalves and Romano 2013). In addition, micropropagation methods can contribute to conserve the germplasm bank and propagation of rare and endangered medicinal plants, produce plants free of diseases, hold better scope and future for the production of important plant-based phytopharmaceuticals and offer a lucrative alternative approach to conventional methods in producing controlled amounts of biochemical(Gupta et al. 2020; Hsie et al. 2019b).
The quality (wavelength) and intensity (irradicance) of light are environmental factors that must be controlled, since they affect photosynthesis, photomorphogenesis and the secondary metabolism of plants in vitro (Alvarenga et al. 2015; Andrade et al. 2017; Carvalho et al. 2020; Hsie et al. 2019a; Lazzarini et al. 2018; Silva et al. 2017). Moreover, the microclimate created inside containers can cause physiological and morphological disorders in plants, such as malformed roots, low chlorophyll concentration, reduced survival at acclimation and high hyperhydricity (Isah 2015). According to Kumar and Palni (2003), plantlets of Rosa damascena Mill. and Rhynchostylis retusa (L.) Bl. developed under photosynthetically active radiation (PAR) showed better survival and growth following transfer to pots compared to cool fluorescent light due to higher chlorophyll content and possibly better organization of the photosynthetic apparatus. Photon flux is a factor of paramount importance for explant growth response and plantlet quality in vitro. The responses of explants to light intensity depend on the plant genotype, for example, low light intensity (27 µmol m−2 s−1) significantly influenced the growth and development of Achillea millefolium in vitro (Alvarenga et al. 2015). Hsie et al. (2019a) used 5 different light intensities of cool white fluorescent lamps, namely: 20, 54, 78, 88 and 110 μmol m−2 s−1, and found that the lowest intensities (20 and 54 μmol m −2 s−1) generated the highest total dry weight in Lippia rotundifolia. Lazzarini et al. (2018) adopted light intensities of 26, 51, 69, 94, or 130 μmol m−2 s−1 of fluorescent lamps and recorded higher total dry weight under increased light intensity, mainly under 94 μmol m−2 s−1, for both the apical and nodal explants of Lippia gracilis. Nodal segments of Plectranthus amboinicus were grown under the following light intensities: 26, 51, 69, 94 and 130 μmol m−2 s−1 of cool-white fluorescent lamps and showed that intermediate light intensity (69 μmol m−2 s−1) increased plantlets’ dry weight (Silva et al. 2017).
Overall, the culture in vitro uses cool fluorescent lamps (white light) in tissue culture growth chamber (Gupta and Jatothu 2013). The combination of light-emitting diodes (LEDs) at different wavelengths would be an alternative to improve plant growth and active compound production in vitro. Responses to different wavelengths also affect growth and metabolite compounds in vitro (Batista et al. 2018). Alvarenga et al. (2015) observed differences in the number of compounds in the essential oil of Achillea millefolium grown under different wavelenghts. Hsie et al. (2019a) assessed Lippia rotundifolia and observed better growth parameters and total chlorophyll content at spectra combination 2.5Red:1Blue. However, the most significant leaf and total dry weight gain was recorded for Lippia gracilis subjected to monochromatic red light wavelength (Lazzarini et al. 2018), which also led to longer shoot length and higher dry weight in plantlets, as well as to carvacrol content higher than that provided by fluorescent lamps (Silva et al. 2017).
Therefore, an efficient and specific micropropagation protocol must be developed for each species. Evaluating parameters that influence growth, such as irradiance and wavelength available to plantlets, is a way to find the best conditions for the expression of plant cell totipotence. These parameters allow plants to produce different compounds and amounts of secondary metabolites of interest (Alvarenga et al. 2015).
The literature lacks information on environmental conditions favorable to the micropropagation of Urtica dioica and on the quantification of secondary metabolites of species grown in vitro. Thus, the aim of the present study was to evaluate the effect of light intensity and wavelength (quality of light) on growth, photosynthetic pigment accumulation and antioxidant activity of micropropagated U. dioica plantlets.
Materials and methods
Culture in vitro
Donor plants of nodal segments were collected in the Medicinal Plants Garden at Federal University of Lavras (UFLA, Lavras, Minas Gerais, Brazil). The voucher specimen was deposited in the PAMG Herbarium of the Agricultural Research Company of Minas Gerais (EPAMIG) under registration number 58,577. Explants (nodal segments) were disinfected in running water added with commercial detergent (30 min). This procedure was followed by explants’ immersion in 70% alcohol (30 s) and in bleach (1.25% active chlorine) for 15 min under constant stirring; subsequently, they were washed in autoclaved distilled water (3 times). Next, nodal segments with approximately 1 cm in length were inoculated in test tubes filled with 12.5 mL of Murashige and Skoog (1962) medium (MS), without growth regulator and supplemented with 30 g L−1 sucrose, 5.5 g L−1 agar and pH adjusted to 5.7 ± 0.1, before autoclaving (for 20 min, at 121 °C). The tubes were kept in growth room under cool-white fluorescent light (42 μmol−2 s−1) at 26 °C ± 1 °C and 16-h photoperiod. Plantlets were subcultured every 40 days until the plant material needed for experiment set up was ready.
LED lamp wavelengths
Nodal segments (from the 1st and 2nd leaf pairs—counting from shoot apex to root) of pre-established U. dioica plantlets were standardized at approximately 1 cm. Next, they were inoculated in test tubes filled with 12.5 mL of Murashige and Skoog (1962) medium (MS), without growth regulator and supplemented with 30 g L−1 sucrose, 5.5 g L−1 agar and pH adjusted to 5.7 ± 0.1, before autoclaving (for 20 min, at 121 °C). The study followed a completely randomized experimental design with 7 treatments and 4 repetitions—5 tubes per repetition, one explant per tube. The tubes were kept in growth room at 26 °C ± 1 °C under 16-h photoperiod and different light emitting diodes (LEDs TECNAL© Piracicaba, Brazil): monochromatic (red and blue), white LED and combinations of red and blue (1red/1blue, 2.5red/1blue and 1red/2.5blue). The cool-white fluorescent light (Osram©, Brazil) at 42 μmol m−2 s−1 light intensity was used as the control treatment. The proportions between the red and the blue led are derived from the number of diodes present in a lamp, being 1red/1blue (6 red diodes and 6 blue diodes), 2.5red/1blue (6 red diodes and 6 blue diodes) and 1red/2.5blue (6 red diodes and 6 blue diodes). Color LED light was measured with portable spectrometer SPECTRA PEN Z850 (Qubit Systems-Kingston, Ontario-Canada).
Light intensity
Nodal segments (from the 1st and 2nd leaf pairs—counting from shoot apex to root) of pre-established U. dioica plantlets were standardized at approximately 1 cm. Next, nodal segments were inoculated in test tube filled with 12.5 mL of Murashige and Skoog (1962) medium (MS), without growth regulator and supplemented with 30 g L−1 sucrose, 5.5 g L−1 agar and pH adjusted to 5.7 ± 0.1, before autoclaving (for 20 min, at 121 °C). The study followed a completely randomized experimental design with 5 treatments and 4 repetitions—5 tubes per repetition, one explant per tube. The tubes were kept in growth room at 26 °C ± 1 °C and 16-h photoperiod under different LED light intensities (26, 51, 69, 94 and 130 μmol−2 s−1). These intensities were provided by the White LEDs (TECNAL© Piracicaba, Brazil). Photosynthetic flux photon density was measured in QSO-S PRO CHECK + PAR PHOTON FLUX SENSOR (DECAGON DEVICES; Pullman, Washington, USA).
Growth analyses
The length (cm) of plantlets (PL) and of the largest root (LRL); leaf (LDW), stem (SDW), root (RDW), shoot (ShDW = LDW + SDW) dry weight (mg), ratio of ShDW/RDW and total dry weight (TDW; mg) were assessed. In all experiments, each treatment was represented by 20 plantlets. Plantlet parts were packed in kraft paper bags, dehydrated in forced air circulation oven, at 37 ºC, for 72 h, and weighed on precision scale. Five plantlets per treatment were used to evaluate leaf area (cm2) in the WinFOLIA™ software and EPSON PERFECTION V700 PHOTO scanner.
Photosynthetic pigment analyses
In total, 50 mg of fresh leaf tissue from plants in each treatment were collected for photosynthetic pigment analysis, which was carried out in dark room only lit by green light. The second pair of leaves from each plantlet was used in the experiment. Leaves were placed in Falcon tubes and wrapped in aluminum foil; next, 10 mL of dimethyl sulfoxide (DMSO) were added to the tubes, which were taken to the oven at 65 °C, for 24 h. Subsequently, 3 mL of supernatant were placed in quartz cuvettes for absorbance reading at wavelengths of 480 nm (carotenoids), 649 nm (chlorophyll a) and 665 nm (chlorophyll b) (Hiscox and Israelstam 1979). Readings were performed in microplate reader TECAN INFINITY M200 PRO operated with data processing system I-control® version 3.37. Chlorophyll and carotenoid concentrations were calculated based on equations proposed by Wellburn (1994):
Results were expressed in milligrams of pigment per gram of fresh leaf tissue (mg g−1). All treatments were evaluated in triplicate;
Phytochemical content and antioxidant activity analyses
Plant extracts were prepared with powdered dry leaves. The aliquot of 15 mg of powder was weighed in the microtubes. The volume of 2 mL of MeOH: H2O (1: 1) was added to the tubes, which were shaken and subjected to sonication extraction for 15 min at room temperature; this procedure was followed by centrifugation at 10,000 RPM (10 min). Subsequently, the supernatant was collected and stored in freezer (− 20 °C) until the time to carry out the analyses. The molecular absorption readings in ultraviolet light were measured in microplate reader TECAN INFINITY M200 PRO operated with data processing system I-control® version 3.37. All treatments were evaluated in triplicate.
Total phenolic compounds (TPC)
The amount of total phenolic compounds was measured based on the Folin–Ciocalteau method, with modifications (Singleton and Rossi 1965). Aliquots of 50 µL of extract, 130 µL of Folin–Ciocalteau ethanolic solution (10%) and 130 µL of Na2CO3 (7.0%) were added to 2-ml microtubes. Samples were incubated at room temperature for 2 h, in the dark. Subsequently, 300 µL were added to 96-well microplates; absorbance was read at 760 nm. The calibration curve consisted of gallic acid standard (Sigma-Aldrich®, ≥ 98%) at the range 0.0078 to 0.25 mg mL−1 to generate the equation y = 6.2873x + 0.9239 (R2 = 0.9986). Results were expressed in milligrams of gallic acid equivalent per gram of leaf dry weight (mgGAE g LDW−1).
Total flavonoids (TF)
The amount of flavonoids was calculated based on the methodology by Atanassova et al. (2011). The aliquot of 100 μL of sample was added to 100 μL of distilled water and to 50 μL of NaNO2 in 2-mL microtubes, which were stirred. After 5-min stirring, the aliquot of 50 μL of AlCl3 was added to the tubes. After 6 min, 100 μL of NaOH (1 M) was also added to them. Then, 200 μL of samples were pipetted into 96-well microplates; absorbance was measured at 510 nm. The control comprised 100 μL of sample and 300 μL of distilled water. White samples were composed of 100 μL of sample, 100 μL of NaOH (1 M) and 200 μL of distilled water. Calibration curve resulted from quercetin standard (Sigma-Aldrich®, ≥ 95%) diluted at the range 0.0625–1.0 mg mL−1 (y = 3.72x + 0.2596, R2 = 0.9901). Flavonoid accumulation was expressed in milligrams equivalent of quercetin per gram of leaf dry weight (mgQE g LDW−1).
Total antioxidant capacity (TAC)
Total antioxidant capacity was evaluated through the molybdate reduction methodology (Prieto et al. 1999), with modifications. The aliquot of 50 μL of extract was added to 1 mL of the TAC reagent solution (monobasic sodium phosphate (28 mM), ammonium molybdate (4 mM) and sulfuric acid (0.6 M), at equal rates, in 2-mL microtubes, which were incubated at 95 ºC (90 min) and left to cool at room temperature. Then, 200 μL of samples were pipetted in 96-well microplates; absorbance was measured at 695 nm. Calibration curve resulted from ascorbic acid standard (Sigma-Aldrich®, ≥ 99.7%) at concentrations of 0.002–0.5 mg mL−1 (y = 6.80x + 0.0716, R2 = 0.9946). Results were expressed in milligrams of ascorbic acid equivalents per gram of leaf dry weight (mgAAE g LDW−1).
Free radical scavenging activity (DPPH)
The 2,2-diphenyl-1-picrylhydrazyl (DPPH) radical capture method was used to determine the free radical scavenging activity based on the methodology by Brand-Williams et al. (1995), with modifications, by using a 92.8º GL ethanolic solution of DPPH (60 μg mL−1) kept away from light and refrigeration. The original extractive solution (7.5 g mL−1) and sequential dilution of each sample in MeOH:H2O 1:1 up to 1/128 were used in the experiment. The aliquot of 250 µL of the DPPH solution was added to 50 µL of samples in 96-well microplates. The plates were incubated (60 min) in the dark, at room temperature; absorbance was read at 517 nm. DPPH radical elimination rate was calculated through the following formula: %AA = (A0 – A1)/A0 × 100; wherein, A0: absorbance of the DPPH ethanolic solution; A1: absorbance of the DPPH solution + extract. The free radical scavenging activity was expressed as IC50 (μg mL−1), which corresponds to the concentration of extract capable of causing 50% DPPH radicals’ inhibition.
Oxygen radical absorbance capacity (ORAC)
Oxygen Radical Absorbance Capacity followed the methodology by Ou et al. (2001), with modifications. The aliquot of 25 μL of sample was added to 150 μL of fluorescein (70 mM)—prepared in phosphate buffer (75 mM and pH 7.4)—in 96-well black microplates. The microplate was pre-incubated (10 minu) at 37 °C; next, 30 μL of AAPH-12 mM (2,2′-Azobis (2-amidinopropane) dihydrochloride) was added to it. Trolox standard, at concentrations ranging from 0.25 to 0.015625 mg mL−1 was used as control. Results were calculated based on the regression equation between Trolox concentration and AUC (area under the curve) (AUC sample-AUC white). Values were expressed in mg of Trolox equivalent per gram of leaf dry weight (mgTE g LDW−1).
Statistical analysis
Collected data were subjected to analysis of variance. The means recorded for the treatments were compared through Scott-Knott test at 5% probability level in the R Development Core Team software (2012). The software Statistica® version 13.5 (StatSoft; Tulsa, OK, USA) was used to perform the Principal Component Analysis (PCA).
Results and discussion
LED lamps’ wavelength: growth analyses
Wavelength significantly influenced (P < 0.05) the growth of U. dioica plantlets in vitro (Table 1 and Fig. 1b). Overall, the best growth results were observed for plantlets grown under LED (2.5R:1B): higher root growth (2.09 cm), number of leaves (9.90), number of nodal segments (4.95) and the highest dry weights (Table 1, Fig. 2). Fluorescent light (control) led to total dry weight gain of 14.34 mg plantlet−1; it was 24% lower than in the culture subjected to wavelength of 2.5R:1B. On the other hand, U. dioica plantlets did not develop well under the red and blue monochromatic lights. Monochromatic red light led to longer shoot (12.03 cm) and internode lengths characteristic of etiolated plantlets (Fig. 1b and Table 1). Red light also led to the shortest root length (0.66 cm) (Table 1) and the lowest values of parameters such as leaf (1.95 mg plant−1), stem (2.86 mg plant−1), root (0.69 mg plant−1), shoot (4.81 mg plant−1) and total dry weight (5.50 mg plant−1) (Fig. 2). There was directly proportional association between increased red light proportion and shoot growth induction.

Urtica dioica plantlets grown in vitro under a light intensities of 26, 51, 69, 94 and 130 μmol m−2 s−1 at b different wavelenghts, after 40 days

Growth of U. dioica grown (in vitro) in MS medium, for 40 days, under different light wavelengths (red, blue, 2.5R:1B, 1R:2.5B, 1R:1B, white LED, fluorescent); leaf (LDW), stem (SDW), root (RDW), shoot dry weight (ShDW = LDW + SDW), total (TDW), ratio ShDW/RDW. (Color figure online)
Plectranthus amboinicus also presented longer shoot length under monochromatic red led (Silva et al. 2017) and Lippia rotundifolia shoot elongation stimulated by the red spectrum (Hsie et al. 2019a), similarly to Ajuga multiflora (Jeong and Sivanesan 2018) and Vaccinium corymbosum (Hung et al. 2016). Stem elongation under red light can be an endogenous hormonal response associated with gibberellin in plantlets. According to Kamiya and Garcia-Martinez (1999) and Manivannan et al. (2015), the red light accounts for the activation of genes linked to gibberellin production.
Monochromatic blue light and higher proportions of blue (1R:2.5B) generated shorter plantlet length (3.14 and 4.05 cm, respectively) and lower dry weight. Dry weight gain depends on plant genotype. Hsie et al. (2019a) assessed Lippia rotundifolia and Andrade et al. (2017) evaluated Hyptis suaveolens; they found that the combination 2.5R:1B led to the best dry weight gain. Species Lippia gracilis (Lazzarini et al. 2019) and Plectranthus amboinicus (Silva et al. 2017) showed the highest dry weight under monochromatic red light. However, monochromatic blue light led to the highest dry weight accumulation in Achillea millefolium (Alvarenga et al. 2015). Batista et al. (2018) and Massa et al. (2008) reported that the wavelenght of LED lamps leads to higher growth gain in every species, even among cultivars. More physiological studies are needed to help better understanding why different wavelengths made several species respond differently.
LED lamps’ wavelength: photosynthetic pigment analyses
The photosynthetic apparatus of Urtica dioica was evaluated based on photosynthetic pigment levels; this outcome evidenced its sensitivity to variations in spectral light (Table 2). Wavelenght had impact on plantlets’ physiological response, which reflected the photosynthetic pigment levels. Blue and white wavelengths did not differ from that of the fluorescent light (control) when it comes to chlorophyll a concentration in plantlets. According to Kwon et al. (2015), blue light wavelength in White LED and fluorescent light is important for chlorophyll synthesis, as well as for chloroplast development and for photomorphogenesis. Cioć et al. (2018) reported the highest amount of photosynthetic pigment under fluorescent light. Blue light regulates the activation of enzymes and increases the expression of genes involved in the chlorophyll biosynthesis pathway (Fan et al. 2013; Ruyters 1984). Senger (1982) reported that one of the roles played by the blue wavelength lies on chloroplast and chlorophyll formation.
With respect to the current study, the large amount of energy observed in blue light wavelength induced plants to displace the photoassimilates for the biosynthesis of secondary metabolites responsible for protecting of photosynthetic apparatus (carotenoids 0.18 mg g−1 FW). Plants with morphological and physiological plasticity adjust to lighting conditions (Taiz and Zeiger 2017; Yang et al. 2018); they can use their photoassimilates for plant growth and development (Park and Runkle 2017).
Monochromatic blue light, combinations “1R:2.5B, 2.5R:1B” and “1R:1B”, and fluorescent light allowed the higher chlorophyll a/b ratio values (4.36, 3.99, 3.51, 3.94 and 3.85, respectively). Based on these results, they enabled greater electron transport capacity and allowed more efficient photosynthesis (Zheng and Van Labeke 2017). According to results in the present study, photoassimilates are likely produced in plantlets subjected to monochromatic blue, White LED and fluorescent light. They focused on chlorophyll and carotenoid production, rather than on plantlet growth or on the conversion of phenolic compounds and flavonoids (Table 2). The product of plant photosynthesis under 1R:2.5B seemed to head towards the production of phenolic compounds and flavonoids (Table 2). Plants subjected to 2.5R:1B recorded the largest dry weight production rates (Fig. 2).
Plantlets grown under LEDs at red proportions of R; 1R:1B; 2.5R:1B and 1R:2.5B recorded lower chlorophyll values (Table 2). Red wavelength reduces chlorophyll biosynthesis and acts in aminolevulinic-5 acid decrease—it is a precursor in chlorophyll production (Zheng and Van Labeke 2017). The use of monochromatic red light in the present study led to low concentration of photosynthetic pigments that account for defective photosynthesis and for low dry weight production (Fig. 2). Light wavelength acts in carotenoid biosynthesis regulation, which is a species-specific response (Zhang et al. 2015). Treatments with red light wavelength led to lower production of carotenoids (Table 2). Carotenoid accumulation is related to gibberellin levels in plants (Mohanty et al. 2016), since they have geranylgeranyl diphosphate (GGPP) as common precursor in their biosynthetic route (Ruiz-Sola and Rodríguez-Concepción 2012). This low carotenoid production can be related to increased plant length, which suggests that the assessed plant used its photoassimilates to produce gibberellins. Gibberellin biosynthesis genes are activated when phytochromes receive red light; this process controls plantlets’ stem lengthening (Kamiya and Garcı́a-Martı́nez 1999).
Although previous results have confirmed the physiological and morphological effects of light quality, responses change depending on plant species; one cannot determine certain effects of light quality. Accordingly, it is necessary to apply proper light (LED system) in order to meet different purposes, such as shoot and root growth promotion or inhibition, bulbs formation and growth, flowering control, among others.
LED lamps’ wavelength: phytochemical content and antioxidant activity analyses
Natural antioxidants found in vegetables can slow down the process of many chronic diseases, prevent cancer onset and increase food shelf life (Oskoueian et al. 2011). It is possible determining the antioxidant capacity of medicinal plant extracts through different methods. Methods such as DPPH (2,2-diphenyl-1-picrylhydrazyl), Total antioxidant capacity (TAC) and Oxygen Radical Absorbance Capacity (ORAC) were used in the present experiment. All these three methods carry out absorbance reading in spectrophotometer (Ojha et al. 2018). DPPH is a free radical that has one unpaired electron in one of its Nitrogen atoms (purple color). This electron is paired with one electron of hydrogen due to the contact with an antioxidant substance and forms a yellowish chemical complex (Oliveira 2015). The total antioxidant capacity (TAC) assay is based on Mo6+ to Mo5+ reduction by antioxidant compounds in plantlets’ culture—the solution turns greenish (Khan et al. 2012). The Oxygen Radical Absorbance Capacity (ORAC) assay oxidizes fluorescein after the addition of peroxyl radicals. Antioxidant compounds in plants protect fluorescein by blocking the oxidation reaction (Ou et al. 2001).
There was significant difference (P < 0.05) in the concentration of phenolic compounds, flavonoids and antioxidant activity of Urtica dioica grown under different light spectra in vitro (Table 3). Wavelength of 1R:2.5B led to increased total phenolic compound and flavonoid content; it was 5.52 and 15.82 fold higher than that of plants under control conditions (fluorescent light). Antioxidant activity tests (DPPH, CAT and ORAC) also showed the highest concentrations under wavelength 1R:2.5B (Table 3), the lowest content was recorded for plants under fluorescente light. Thus, it can be inferred that phenolic compounds and flavonoids account for the species' antioxidant activity. Jing et al. (2018) reported that the induced biosynthesis of these compounds makes plants more resistant to diseases and environmental stresses. Phenolic compounds quercetin, rutin, isoquercitrin and 5-0-caffeoylquinic acid are the main compounds participating in the antioxidant activity of U. dioica (Jan et al. 2017; Orčić et al. 2014). They have anti-inflammatory and antioxidant activity (Ao et al. 2008; Rogerio et al. 2007).
Plants respond to the environment in different ways, they allocate their photoassimilates to the biosynthesis of elements necessary for their survival and adapt to different light wavelengths. Lobiuc et al. (2017) observed the wavelength effect on the production of phenolic and flavonoid compounds in Ocimum basilicum—2R:1B produced the highest concentrations. Therefore, light wavelength affects the biosynthesis of phenolic and flavonoid compounds in plants differently, depending on the species and on growing conditions (Cioć et al. 2018; Huché-Thélier et al. 2016; Lobiuc et al. 2017; Pedroso et al. 2017; Taulavuori et al. 2018; Zhang et al. 2018).
Light intensity: growth analyses
Plantlet length and number of leaves and nodal segments were not affected by different light intensities. Plantlet length ranged from 7.61 to 8.41 cm, number of leaves, from 6.9 to 7.9 and number of nodal segments, from 3.45 to 3.95 (data not shown). Plantlets under 26, 51 and 69 μmol m−2 s−1 showed thin and whitish stems in comparison to treatments based on 94 and 130 μmol m−2 s−1 (Fig. 1a). However, total leaf area recorded for the assessed plantlet was greater at intensities of 94 and 130 μmol m−2 s−1 with 11.38–13.15 cm2, respectively; these intensities led to longer root length. Leaf area is directly related to light energy interception; it leads to greater photon conversion into photoassimilates and, consequently, to higher dry weight (Sanquetta et al. 2014), which corroborates results in the current study. Light conditions affect hormonal balance and auxin distribution in plants, as well as promote physiological changes, such as the production of longer roots (Halliday et al. 2009). Yang et al. (2018) reported light influence on auxin production in Glicine max; they observed that the endogenous concentration of this phytohormone is regulated by light intensity and quality.
Different light intensities have significantly influenced (P < 0.05) leaf (LDW), stem (SDW), root (RDW), shoot (ShDW) and total (TDW) dry weight production (Fig. 3). Higher stem and root dry weight gain was observed under 69 μmol m−2 s−1. The intensity of 94 μmol m−2 s−1 was better than that in all other treatments when it comes to leaf (14.45 mg plant−1), shoot (20.05 mg plant−1) and total dry weight (22.59 mg plant−1) results. Lazzarini et al. (2018) reported that Lippia gracilis also recorded higher leaf, shoot, root and total weight due to increased light intensity (94 μmol m−2 s−1). The lowest dry weights were observed in plantlets grown at 26 μmol m−2 s−1 (Fig. 3). Overall, plants grown in low light intensity environments present lower biomass values due to low photoassimilate production caused by inefficient photosynthesis (Taiz and Zeiger 2017). Similar results were found by Silva et al. (2017), who assessed the effect of different light intensities on Plectranthus amboinicus culture in vitro; plantlets grown under 26 μmol m−2 s−1 showed the lowest dry weight production values. However, Achillea millefolium showed the highest values for parameters analyzed under the lowest light intensity, 27 μmol m−2 s−1 (Alvarenga et al. 2015). Light intensities of 20 and 54 μmol m−2 s−1 promoted higher growth in Lippia rotundifolia, as well as photosynthetic pigment content and dry weight production (Hsie et al. 2019a). Andrade et al. (2017) showed that both the apical and nodal explants of Hyptis suaveolens presented better growth under photon flux density of 47 and 69 μmol m−2 s−1.

Growth of U. dioica grown (in vitro) in MS medium for 40 days under different light intensities (26, 51, 69, 94 and 130 μmol m−2 s−1); leaf (LDW), stem (SDW), root (RDW), shoot dry weight (ShDW = LDW + SDW), total (TDW), ratio ShDW/RDW
Different responses in dry weight production in vitro under different light intensities depend on genotype. Each species has its minimum and maximum photon flux density limit in order to growth. High active radiation on plantlets can damage the photosystems (PSI and PSII) and produce reactive oxygen species that, in their turn, impair plant growth. Plants growing in unfavorable environments such as proper light intensity can increase the production of reactive oxygen species and damage proteins and lipids, among others, as well as impair biomass gain (Gill and Tuteja 2010).
The best ShDW/RDW ratio result was observed in plantlets grown under 26 μmol m−2 s−1 (Fig. 3). Differences in shoot/root ratio are responses observed in plants exposed to different light intensities (Fernandes et al. 2013). The lowest values were found at intensities of 69 μmol and 130 m−2 s−1; based on this outcome, plantlets grown at such intensities have higher root production and, consequently, they can better absorb water and mineral salts.
Light intensity: photosynthetic pigment analyses
Different light intensities significantly affected (P < 0.05) the concentration of photosynthetic pigments in Urtica dioica plantlets grown in vitro (Table 4). Intensities of 51 and 94 μmol m−2 s−1 provided the highest chlorophyll a content, total chlorophyll and a/b ratio in comparison to chlorophyll b at intensity of 51 μmol m−2 s−1 (Table 4). However, this intensity did not provide the greatest dry weight gain in plantlets grown in vitro. Light intensity (51 μmol m−2 s−1) favored the biosynthesis of carotenoids in plantlets grown in vitro (0.25 mg g−1).
Light intensity is one of the key factors regulating the genes responsible for the biosynthesis of chlorophyll and carotenoids (Zhang et al. 2015). Low light intensity environments influence the photosynthesis process and affect the production of photosynthetic pigments. There is correlation between greater light intensity and reduced concentration of photosynthetic pigments due to photodegradation (Silva et al. 2017). However, Urtica dioica did not show the same behavior likely due to its plasticity, although it depends on genotype. The high adaptability and rusticity of nettles allowed the species to spread worldwide, it is found in shaded environments, such as dense forests, and in degraded areas in full light (Taylor 2009).
Light intensity: phytochemical contents and antioxidant activity analyses
There was significant difference (P < 0.05) between the phenol and total flavonoid levels in U. dioica depending on the treatment, as well as on the assessed antioxidant activity (Table 5). Dry weight gain was higher in plants grown at intensity of 94 μmol m−2 s−1; intensity of 130 μmol m−2 s−1 impaired plantlet growth in vitro (Fig. 3). This outcome may have resulted from the fact that plantlets are often stressed in vessels due to several factors (light, humidity, etc.), which lead to ROS (reactive oxygen species) overproduction and hinder plantlet growth. Gill and Tuteja (2010) reported that plants have antioxidant defense against oxidative stress damages, be them enzymatic (superoxide dismutase, catalase and others enzymes) and non enzymatic (phenolic compounds, etc.) antioxidant defense systems focused on protecting plants from oxidative damage by ROS scavenging. The highest production of phytochemical contents (TPC and TF) and antioxidant activity (TAC, ORAC and DPPH) were observed in plantlets grown at 130 μmol m−2 s−1 (Table 5).
The three antioxidant activity tests evidenced different analysis principles: TAC to evaluate molybdenum reduction, ORAC to assess fluorescein oxidation and DPPH to find the free radicals. Based on DPPH, the lower the value the better the test result; it indicates the amount of extract to be used in the antioxidant activity. The three tests showed the best results at the highest light intensity; this outcome corroborated the assumption that plants develop antioxidants under higher irradiance levels.
Increased light intensity leads to higher photoassimilate production—up to levels harmful to plants -, caused by light stress (Taiz and Zeiger 2017). High light intensity stimulates the production of phenolic compounds in plants; these compounds protect the photosynthetic apparatus (Warren et al. 2003). Light is one of the environmental factors directly linked to the production of secondary metabolites such as phenolic compounds and flavonoids (Jing et al. 2018; Taulavuori et al. 2018). Plantlets grown in vitro often produce less wax on the leaves and, consequently, have less protection. Thus, increased light intensity may have led to light stress because the plant accumulated phenolic compounds and flavonoids to protect chlorophylls. According to Gobbo-Neto and Lopes (2007), there is positive correlation between light intensity and the production of phenolic compounds, such as flavonoids, since these secondary metabolites absorb and dissipate light energy, and reduce damages to the photosynthetic apparatus.
Leaf trichomes help protecting against high intensity, insect attacks and other factors; interestingly, plantlets grown in vitro showed few trichomes in the leaves. According to Pollard and Briggs (1982), Urtica dioica grown in the shade in low light intensity environments presents fewer trichomes than those grown in full sunlight.
Principal component analysis (PCA)
Multivariate statistical analysis techniques like the Principal Component Analysis (PCA) were used to evaluate correlations among growth, phytochemical contents and antioxidant activity in U. dioica plantlets treated under different LED lamp wavelengths and light intensities. General data evaluations showed that growth parameters, phytochemical contents and antioxidant activities differed in plantlets grown under different wavelengths and light intensities. PCA (scores and loadings) based on 12 parameters (growth and chemical compounds) resulted in 2 principal components that accounted for 81.76% and 88.66% of the total variation in the experimental procedure applied to measure light quality and intensity, respectively. Based on these 12 parameters, it was possible grouping the collected data into 3 clusters to assess light quality (Fig. 4) and intensity (Fig. 5).

Score and loadings of the Principal component analyses (PCA) in the correlation matrix built from data recorded for Total phenolic compounds (TPC), total flavonoids (TF), antioxidant activity (TAC, ORAC, DPPH), photosynthetic pigments (total chlorophyll, and carotenoids) and leaf (LDW), stem (SDW), root (RDW), shoot dry weight (ShDW = LDW + SDW), total (TDW) under different light wavelengths (red, blue, 2.5red:1blue, 1red:2.5blue, 1red:1blue, white LED, fluorescent). (Color figure online)

Score and loadings of the Principal component analyses (PCA) in the correlation matrix built from data recorded for Total phenolic compounds (TPC), total flavonoids (TF), antioxidant activity (TAC, ORAC, DPPH), photosynthetic pigments (total chlorophyll, and carotenoids) and leaf (LDW), stem (SDW), root (RDW), shoot dry weight (ShDW = LDW + SDW), total (TDW) under different light intensities (26, 51, 69, 94 and 130 μmol m−2 s−1)
PCA evidenced that U. dioica plantlets under 1R/2.5B showed higher TPC, TF, TAC, ORAC and DPPH content, and plantlets under 2.5R/1B had higher LDW, SDW, ShDW, RDW and TDW. Plants recorded higher TChl and carotenoids under blue, white and fluorescent light (Fig. 4).Therefore, LED lamps with greater red light compared to blue light (2.5R/1B) stimulated the plant's growth, thus enabling dry weight gains in its organs (leaves, shoots and roots). According to Silva et al. (2017), the improved plantlet growth provided by the red spectrum may be associated with the higher sensitivity of the phytochrome to such spectrum. On the other hand, LEDs with a higher proportion of blue light compared to red light (1R/2.5B) activated the production of phenolic compounds and their antioxidant activity, possibly as a result of a stressful environmental condition. Visible blue light has shorter wavelength; consequently, it has greater energy (Taiz and Zeiger 2017).
In relation on light intensity, it is possible concluding that U. dioica under lower intensity (51 μmol m−2 s−1) showed higher TChl and carotenoid contents. Pantlets grown under 94 μmol m−2 s−1 light intensity recorded higher LDW, SDW, ShDW, RDW and TDW, whereas higher light intensity (130 μmol m−2 s−1) led to more photosynthetic pigments (total chlorophyll and carotenoids) and to higher TPC, TF and antioxidants activity (TAC,ORAC, DPPH) (Fig. 5). Therefore, in an overview, plantlets grown in a condition of high light intensity (130 μmol m−2 s−1) did not reach the maximum in dry weight gain of leaves, shoots and roots, as observed in plantlets submitted to 94 μmol m−2 s−1. Possibly, plants grown in a condition of high light intensity (130 μmol m−2 s−1) initiated a response to a stressful environmental condition, thus reflecting in the increased production of phenolic compounds and antioxidant activity.
Conclusion
Light intensity and quality have affected the growth, photosynthetic pigment concentrations and antioxidant activity in Urtica dioica plantlets. The intensity of 94 μmol−2 s−1 led to the best light conditions for the micropropagation of this species and for the best leaf dry weight production in vitro. The production of photosynthetic pigments was better at intensity of 51 μmol−2 s−1. The concentration of phenolic compounds, flavonoids and antioxidant activity increases at higher light intensities. The antioxidant activity was directly proportional to light intensity increase; this outcome indicates that the use of 130 μmol−2 s−1 initiated an induction of light stress. Wavelenght of 2.5R:1B has favored growth and dry weight accumulation in vitro. Wavelength of 1R:2.5B led to 5.53-fold increase in the concentration of phenolic compounds and to 8.63-fold increase in flavonoid concentation in comparison to fluorescent lamps. Increase red light proportion induced plantlet etiolation.
Abbreviations
- LED:
-
Light emitting diodes
- R:
-
Red
- B:
-
Blue
- F:
-
Fluorescent
- TPC:
-
Total phenolic compounds
- TF:
-
Total flavonoids
- TAC:
-
Total antioxidant capacity
- ORAC:
-
Oxygen radical absorbance capacity
- DPPH:
-
2,2-Diphenyl-1-picrylhydrazyl
References
Alvarenga ICA, Pacheco FV, Silva ST, Bertolucci SKV, Pinto JEBP (2015) In vitro culture of Achillea millefolium L.: quality and intensity of light on growth and production of volatiles. Plant Cell Tissue Organ Cult 122(2):299–308. https://doi.org/10.1007/s11240-015-0766-7
Amaral-Baroli A, Lago JHG, de Almeida CV, de Almeida M, Scotti MT, Leone GF, Soares MG, Cavalari AA, Sartorelli P (2016) Variability in essential oil composition produced by micropropagated (in vitro), acclimated (ex vitro) and in-field plants of Ocimum basilicum (Lamiaceae). Ind Crop Prod 86:180–185. https://doi.org/10.1016/j.indcrop.2016.03.048
Andrade HB, Braga AF, Bertolucci SKV, Hsie BS, Silva ST, Pinto JEBP (2017) Effect of plant growth regulators, light intensity and LED on growth and volatile compound of Hyptis suaveolens (L.) Poit in vitro plantlets. International Society for Horticultural Science (ISHS), Leuven, pp 277–284
Ao C, Li A, Elzaawely AA, Xuan TD, Tawata S (2008) Evaluation of antioxidant and antibacterial activities of Ficus microcarpa L. fil. extract. Food Control 19(10):940–948. https://doi.org/10.1016/j.foodcont.2007.09.007
Atanassova M, Georgieva S, Ivancheva K (2011) Total phenolic and total flavonoid contents, antioxidant capacity and biological contaminants in medicinal herbs. J Univ Chem Technol Metal 46(1):81–88
Batista DS, Felipe SHS, Silva TD, de Castro KM, Mamedes-Rodrigues TC, Miranda NA, Ríos-Ríos AM, Faria DV, Fortini EA, Chagas K, Torres-Silva G, Xavier A, Arencibia AD, Otoni WC (2018) Light quality in plant tissue culture: does it matter? In Vitro Cell Dev Biol Plant 54(3):195–215. https://doi.org/10.1007/s11627-018-9902-5
Brand-Williams W, Cuvelier ME, Berset C (1995) Use of a free radical method to evaluate antioxidant activity. Food Sci Technol 28(1):25–30. https://doi.org/10.1016/S0023-6438(95)80008-5
Carvalho AAd, Bertolucci SKV, Honorato AdC, Rocha TT, Silva ST, Pinto JEBP (2020) Influence of light spectra and elicitors on growth and ascaridole content using in vitro cultures of Dysphania ambrosioides L. Plant Cell Tissue Organ Cult 143(2):277–290. https://doi.org/10.1007/s11240-020-01892-5
Cioć M, Szewczyk A, Żupnik M, Kalisz A, Pawłowska B (2018) LED lighting affects plant growth, morphogenesis and phytochemical contents of Myrtus communis L. in vitro. Plant Cell Tissue Organ Cult 132(3):433–447. https://doi.org/10.1007/s11240-017-1340-2
de Hsie BS, Bueno AIS, Bertolucci SKV, de Carvalho AA, da Cunha SHB, Martins ER, Pinto JEBP (2019a) Study of the influence of wavelengths and intensities of LEDs on the growth, photosynthetic pigment, and volatile compounds production of Lippia rotundifolia Cham in vitro. J Photochem Photobiol B 198:111577. https://doi.org/10.1016/j.jphotobiol.2019.111577
Di Virgilio N, Papazoglou EG, Jankauskiene Z, Di Lonardo S, Praczyk M, Wielgusz K (2015) The potential of stinging nettle (Urtica dioica L.) as a crop with multiple uses. Ind Crop Prod 68:42–49. https://doi.org/10.1016/j.indcrop.2014.08.012
Fan X, Zang J, Xu Z, Guo S, Jiao X, Liu X, Gao Y (2013) Effects of different light quality on growth, chlorophyll concentration and chlorophyll biosynthesis precursors of non-heading Chinese cabbage (Brassica campestris L.). Acta Physiol Plant 35(9):2721–2726. https://doi.org/10.1007/s11738-013-1304-z
Fattahi S, Ghadami E, Asouri M, Ardekanid AM, Akhavan-Niaki H (2018) Urtica dioica inhibits cell growth and induces apoptosis by targeting Ornithine decarboxylase and Adenosine deaminase as key regulatory enzymes in adenosine and polyamines homeostasis in human breast cancer cell lines. Cell Mol Biol 64(3):97–102. https://doi.org/10.14715/cmb/2018.64.3.16
Fernandes VF, Almeida LBd, Feijó EVRdS, Silva DdC, Oliveira RAd, Mielke MS, Costa LCdB (2013) Light intensity on growth, leaf micromorphology and essential oil production of Ocimum gratissimum. Rev Bras Farmacogn 23:419–424
Ghaima KK, Hashim NM, Ali SA (2013) Antibacterial and antioxidant activities of ethyl acetate extract of nettle (Urtica dioica) and dandelion (Taraxacum officinale). J Appl Pharm Sci 3(5):096–099. https://doi.org/10.7324/JAPS.2013.3518
Gill SS, Tuteja N (2010) Reactive oxygen species and antioxidant machinery in abiotic stress tolerance in crop plants. Plant Physiol Biochem 48(12):909–930. https://doi.org/10.1016/j.plaphy.2010.08.016
Gobbo-Neto L, Lopes NP (2007) Plantas medicinais: fatores de influência no conteúdo de metabólitos secundários. Quim Nova 30:374–381. https://doi.org/10.1590/S0100-40422007000200026
Gonçalves S, Romano A (2013) In vitro culture of lavenders (Lavandula spp.) and the production of secondary metabolites. Biotechnol Adv 31(2):166–174. https://doi.org/10.1016/j.biotechadv.2012.09.006
Gülçin İ, Küfrevioǧlu Öİ, Oktay M, Büyükokuroǧlu ME (2004) Antioxidant, antimicrobial, antiulcer and analgesic activities of nettle (Urtica dioica L.). J Ethnopharm 90(2):205–215. https://doi.org/10.1016/j.jep.2003.09.028
Gupta SD, Jatothu B (2013) Fundamentals and applications of light-emitting diodes (LEDs) in in vitro plant growth and morphogenesis. Plant Biotechnol Rep 7(3):211–220. https://doi.org/10.1007/s11816-013-0277-0
Gupta N, Jain V, Joseph MR, Devi S (2020) A review on micropropagation culture method. Asian J Pharm Res Dev 8(1):86–93. https://doi.org/10.22270/ajprd.v8i1.653
Halliday KJ, Martínez-García JF, Josse E-M (2009) Integration of light and auxin signaling. CSH Perspect Biol. https://doi.org/10.1101/cshperspect.a001586
Hiscox JD, Israelstam GF (1979) A method for the extraction of chlorophyll from leaf tissue without maceration. Can J Bot 57(12):1332–1334. https://doi.org/10.1139/b79-163
Hsie BSd, Bueno AIS, Bertolucci SKV, de Carvalho AA, Martins ER, Pinto JEBP (2019b) Growth regulators induced shoot regeneration and volatile compound production in Lippia rotundifolia Cham., a threatened medicinal plant. Ind Crop Prod 137:401–409. https://doi.org/10.1016/j.indcrop.2019.05.050
Huché-Thélier L, Crespel L, Gourrierec JL, Morel P, Sakr S, Leduc N (2016) Light signaling and plant responses to blue and UV radiations—perspectives for applications in horticulture. Environ Exp Bot 121:22–38. https://doi.org/10.1016/j.envexpbot.2015.06.009
Hung CD, Hong C-H, Kim S-K, Lee K-H, Park J-Y, Nam M-W, Choi D-H, Lee H-I (2016) LED light for in vitro and ex vitro efficient growth of economically important highbush blueberry (Vaccinium corymbosum L.). Acta Physiol Plant 38(6):152. https://doi.org/10.1007/s11738-016-2164-0
Isah T (2015) Adjustments to in vitro culture conditions and associated anomalies in plants. Acta Biol Cracov Bot 57(2):9–28
Jan KN, Zarafshan K, Singh S (2017) Stinging nettle (Urtica dioica L.): a reservoir of nutrition and bioactive components with great functional potential. J Food Meas Charact 11(2):423–433. https://doi.org/10.1007/s11694-016-9410-4
Jeong BR, Sivanesan I (2018) Impact of light quality and sucrose on adventitious shoot regeneration and bioactive compound accumulation in Ajuga multiflora Bunge. Sci Hortic 236:222–228. https://doi.org/10.1016/j.scienta.2018.03.056
Jing X, Wang H, Gong B, Liu S, Wei M, Ai X, Li Y, Shi Q (2018) Secondary and sucrose metabolism regulated by different light quality combinations involved in melon tolerance to powdery mildew. Plant Physiol Biochem 124:77–87. https://doi.org/10.1016/j.plaphy.2017.12.039
Kamiya Y, Garcı́a-Martı́nez JL (1999) Regulation of gibberellin biosynthesis by light. Curr Opin Plant Biol 2(5):398–403. https://doi.org/10.1016/S1369-5266(99)00012-6
Khan RA, Khan MR, Sahreen S, Ahmed M (2012) Assessment of flavonoids contents and in vitro antioxidant activity of Launaea procumbens. Chem Cent J 6(1):43. https://doi.org/10.1186/1752-153x-6-43
Kumar A, Palni LMS (2003) The effect of light source and gelling agent on micropropagation of Rosa damascena Mill. and Rhynchostylis retusa (L.) Bl. J Hortic Sci Biotechnol 78(6):786–792. https://doi.org/10.1080/14620316.2003.11511700
Kwon AR, Cui H-Y, Lee H, Shin H, Kang K-S, Park S-Y (2015) Light quality affects shoot regeneration, cell division, and wood formation in elite clones of Populus euramericana. Acta Physiol Plant 37(3):65. https://doi.org/10.1007/s11738-015-1812-0
Lazzarini LES, Bertolucci SKV, Pacheco FV, dos Santos J, Silva ST, de Carvalho AA, Pinto JEBP (2018) Quality and intensity of light affect Lippia gracilis Schauer plant growth and volatile compounds in vitro. Plant Cell Tissue Organ Cult 135(3):367–379. https://doi.org/10.1007/s11240-018-1470-1
Lazzarini LES, Bertolucci SKV, de Carvalho AA, Santiago AC, Pacheco FV, Yucesan B, Pinto JEBP (2019) Explant type and natural ventilation systems influence growth and content of carvacrol and thymol of Lippia gracilis Schauer. Plant Cell Tissue Organ Cult 137(1):33–43. https://doi.org/10.1007/s11240-018-01548-5
Lobiuc A, Vasilache V, Oroian M, Stoleru T, Burducea M, Pintilie O, Zamfirache M-M (2017) Blue and red LED illumination improves growth and bioactive compounds contents in acyanic and cyanic Ocimum basilicum L. microgreens. Molecules 22(12):2111
Manivannan A, Soundararajan P, Halimah N, Ko CH, Jeong BR (2015) Blue LED light enhances growth, phytochemical contents, and antioxidant enzyme activities of Rehmannia glutinosa cultured in vitro. Hortic Environ Biotechnol 56(1):105–113. https://doi.org/10.1007/s13580-015-0114-1
Massa DG, Kim H-H, Wheeler RM, Mitchell CA (2008) Plant productivity in response to LED lighting. HortScience 43(7):1951–1956. https://doi.org/10.21273/hortsci.43.7.1951
Mohammadi A, Mansoori B, Aghapour M, Baradaran B (2016) Urtica dioica dichloromethane extract induce apoptosis from intrinsic pathway on human prostate cancer cells (PC3). Cell Mol Biol 62(3):78–83
Mohanty B, Lakshmanan M, Lim S-H, Kim JK, Ha S-H, Lee D-Y (2016) Light-specific transcriptional regulation of the accumulation of carotenoids and phenolic compounds in rice leaves. Plant Signal Behav 11(6):e1184808. https://doi.org/10.1080/15592324.2016.1184808
Murashige T, Skoog F (1962) A revised medium for rapid growth and bio assays with tobacco tissue cultures. Physiol Plant 15(3):473–497
Ojha K, Dubey S, Chandrakar J, Minj R, Dehariya R, Dixit A (2018) A review on different methods of determination of antioxidant activity assay of herbal plants. Life Sci Inform Public 4(6):707–730
Oliveira GLS (2015) Determinação da capacidade antioxidante de produtos naturais in vitro pelo método do DPPH": estudo de revisão. Rev Bras Plantas Med 17:36–44. https://doi.org/10.1590/1983-084X/12_165
Orčić D, Francišković M, Bekvalac K, Svirčev E, Beara I, Lesjak M, Mimica-Dukić N (2014) Quantitative determination of plant phenolics in Urtica dioica extracts by high-performance liquid chromatography coupled with tandem mass spectrometric detection. Food Chem 143:48–53. https://doi.org/10.1016/j.foodchem.2013.07.097
Oskoueian E, Abdullah N, Ahmad S, Saad WZ, Omar AR, Ho YW (2011) Bioactive compounds and biological activities of Jatropha curcas L. kernel meal extract. Int J Mol Sci 12(9):5955–5970
Ou B, Hampsch-Woodill M, Prior RL (2001) Development and validation of an improved oxygen radical absorbance capacity assay using fluorescein as the fluorescent probe. J Agric Food Chem 49(10):4619–4626. https://doi.org/10.1021/jf010586o
Park Y, Runkle ES (2017) Far-red radiation promotes growth of seedlings by increasing leaf expansion and whole-plant net assimilation. Environ Exp Bot 136:41–49. https://doi.org/10.1016/j.envexpbot.2016.12.013
Pedroso RCN, Branquinho NAA, Hara ACBAM, Costa AC, Silva FG, Pimenta LP, Silva MLA, Cunha WR, Pauletti PM, Januario AH (2017) Impact of light quality on flavonoid production and growth of Hyptis marrubioides seedlings cultivated in vitro. Rev Bras Farmacogn 27:466–470. https://doi.org/10.1016/j.bjp.2016.12.004
Pollard AJ, Briggs D (1982) Genecological studies of Urtica dioica L. New Phytol 92(3):453–470. https://doi.org/10.1111/j.1469-8137.1982.tb03403.x
Prieto P, Pineda M, Aguilar M (1999) Spectrophotometric quantitation of antioxidant capacity through the formation of a phosphomolybdenum complex: specific application to the determination of vitamin E. Anal Biochem 269(2):337–341. https://doi.org/10.1006/abio.1999.4019
R, Development, Core, Team (2012) R: a language and environment for statistical computing. R Foundation for Statistical Computing, Vienna
Rogerio AP, Kanashiro A, Fontanari C, da Silva EVG, Lucisano-Valim YM, Soares EG, Faccioli LH (2007) Anti-inflammatory activity of quercetin and isoquercitrin in experimental murine allergic asthma. J Inflamm Res 56(10):402–408. https://doi.org/10.1007/s00011-007-7005-6
Ruiz-Sola MÁ, Rodríguez-Concepción M (2012) Carotenoid biosynthesis in Arabidopsis: a colorful pathway. Am Soc Plant Biol 10:e0158
Rutto LK, Xu Y, Ramirez E, Brandt M (2013) Mineral properties and dietary value of raw and processed stinging nettle (Urtica dioica L.). Int J Food Sci 2013:857120. https://doi.org/10.1155/2013/857120
Ruyters G (1984) Effects of blue light on enzymes. In: Senger H (ed) Blue light effects in biological systems. Springer, Berlin, pp 283–301
Sanquetta CR, Behling A, Corte AD, Cadori GC, Costa S Jr, Macedo JP (2014) Eficiência de conversão da radiação fotossintética interceptada em fitomassa de mudas de Eucalyptus dunii Maiden em função da densidade de plantas e do ambiente de cultivo. Sci For 42(104):573–580
Senger H (1982) The effect of blue light on plants and microorganisms. Photochem Photobiol 35(6):911–920. https://doi.org/10.1111/j.1751-1097.1982.tb02668.x
Silva ST, Bertolucci SKV, da Cunha SHB, Lazzarini LES, Tavares MC, Pinto JEBP (2017) Effect of light and natural ventilation systems on the growth parameters and carvacrol content in the in vitro cultures of Plectranthus amboinicus (Lour.) Spreng. Plant Cell Tissue Organ Cult. https://doi.org/10.1007/s11240-017-1195-6
Singleton VL, Rossi JA (1965) Colorimetry of total phenolics with phosphomolybdic-phosphotungstic acid reagents. Am J Enol Viticult 16(3):144
Taiz L, Zeiger E (2017) Fisiologia e desenvolvimento vegetal, 6th edn. Artmed Editora, Porto Alegre
Taulavuori K, Pyysalo A, Taulavuori E, Julkunen-Tiitto R (2018) Responses of phenolic acid and flavonoid synthesis to blue and blue-violet light depends on plant species. Environ Exp Bot 150:183–187. https://doi.org/10.1016/j.envexpbot.2018.03.016
Taylor K (2009) Biological flora of the British Isles: Urtica dioica L. J Ecol 97(6):1436–1458. https://doi.org/10.1111/j.1365-2745.2009.01575.x
Upton R (2013) Stinging nettles leaf (Urtica dioica L.): extraordinary vegetable medicine. J Herbal Med 3(1):9–38. https://doi.org/10.1016/j.hermed.2012.11.001
Warren JM, Bassman JH, Fellman JK, Mattinson DS, Eigenbrode S (2003) Ultraviolet-B radiation alters phenolic salicylate and flavonoid composition of Populus trichocarpa leaves. Tree Physiol 23(8):527–535. https://doi.org/10.1093/treephys/23.8.527
Wellburn AR (1994) The spectral determination of chlorophylls a and b, as well as total carotenoids, using various solvents with spectrophotometers of different resolution. J Plant Physiol 144(3):307–313. https://doi.org/10.1016/S0176-1617(11)81192-2
Yang F, Fan Y, Wu X, Cheng Y, Liu Q, Feng L, Chen J, Wang Z, Wang X, Yong T, Liu W, Liu J, Du J, Shu K, Yang W (2018) Auxin-to-Gibberellin ratio as a signal for light intensity and quality in regulating soybean growth and matter partitioning. Front Plant Sci 9:56–56. https://doi.org/10.3389/fpls.2018.00056
Zhang L, Ma G, Yamawaki K, Ikoma Y, Matsumoto H, Yoshioka T, Ohta S, Kato M (2015) Effect of blue LED light intensity on carotenoid accumulation in citrus juice sacs. J Plant Physiol 188:58–63. https://doi.org/10.1016/j.jplph.2015.09.006
Zhang T, Shi Y, Piao F, Sun Z (2018) Effects of different LED sources on the growth and nitrogen metabolism of lettuce. Plant Cell Tissue Organ Cult 134(2):231–240. https://doi.org/10.1007/s11240-018-1415-8
Zheng L, Van Labeke M-C (2017) Long-term effects of red- and blue-light emitting diodes on leaf anatomy and photosynthetic efficiency of three ornamental pot plants. Front Plant Sci. https://doi.org/10.3389/fpls.2017.00917
Acknowledgements
The authors would like to thank the National Council for Scientific and Technological Development (CNPq—Conselho Nacional de Desenvolvimento Científico e Tecnológico), the Coordination for the Improvement of Higher Education Personnel (CAPES—Coordenação de Aperfeiçoamento de Pessoal de Nível Superior) and the Minas Gerais State Research Foundation (FAPEMIG—Fundação de Pesquisa do Estado de Minas Gerais) for financial support (scholarships and research grants).
Author information
Authors and Affiliations
Contributions
The following declarations about authors contributions to the research have been made: concept of the study: JEBPP, SKVB; Intensity and quality light system—design and settings: ADC, JEBPP, SKVB; laboratory research: ADC, CKS, GCS, TO, EAM, JPS; statistical analyses: AAC, ADC; writing of the manuscript ADC, AAC, SKVB, JEBPP.
Corresponding author
Ethics declarations
Conflict of interest
The authors declare no conflict of interests.
Additional information
Communicated by Danny Geelen.
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
About this article

Cite this article
Coelho, A.D., de Souza, C.K., Bertolucci, S.K.V. et al. Wavelength and light intensity enhance growth, phytochemical contents and antioxidant activity in micropropagated plantlets of Urtica dioica L.. Plant Cell Tiss Organ Cult 145, 59–74 (2021). https://doi.org/10.1007/s11240-020-01992-2
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11240-020-01992-2