
Abstract
Apical meristems of multiple shoots produced from axenic seedlings of Kentucky bluegrass (Poa pratensis L.) were used for Agrobacterium tumefaciens-mediated transformation. Transformation parameters were optimized for concentration of bacterial cells, duration of infection, and vacuum infiltration. The highest transformation frequency (1.42%) was obtained by infection with Agrobacterium suspension of OD600 = 0.6 for 5 min, under a negative pressure of 0.5 × 105 Pa. After co-cultivation, the herbicide-resistant plants were rooted and transplanted into flowerpots. Transgenic plants were confirmed by polymerase chain reaction (PCR) assay and Southern blot analysis. Using this transformation system, the betA gene encoding choline dehydrogenase and mutant als gene encoding the enzyme acetolactate synthase were introduced into three Kentucky bluegrass cultivars.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Kentucky bluegrass (Poa pratensis L.) is a typical cool-season perennial turf and fodder grass with a beautiful color and cold, shade, and shear tolerance. However, it is sensitive to high salinity and drought (McDonnell and Conger 1984) and reproduces by facultative apomixes (Akerberg 1979; Bashaw 1980; Grazi et al. 1961; Hanna and Bashaw 1987). The rate of facultative apomixes in some breeds of Kentucky bluegrass is as high as 98% or more (Bashaw 1980). To date, the genetic improvement of Kentucky bluegrass by conventional breeding methods has been faced with many problems, such as the difficulty of plant regeneration, high frequency of somatic variance, intense genotype-dependence, and a long transformation period. Genetic transformation is an alternative method that can help to overcome these problems and improve certain traits that would not have been possible with conventional breeding.
At present, genetic modification of Kentucky bluegrass is still a relatively inefficient procedure and cannot be performed at large scales. Most of the genetic transformation of Kentucky bluegrass has been achieved by direct DNA transfer techniques, such as biolistic bombardment (Ke et al. 1996; Ma et al. 1999; Ha et al. 2001; Meyer et al. 2000; Gao et al. 2006; Xin et al. 2006) and electroporation (Griffin and Zemetra 1999). In recent years, the preferred method for genetic transformation has been an Agrobacterium-mediated transformation system because of the lower transgenic copy number, fewer rearrangements, and stable gene expression involved (Wang and Ge 2006; Gao et al. 2008). So far, only a few studies on successful Agrobacterium-mediated transformations of Kentucky bluegrass have been reported. Chai et al. (2003) transformed embryogenic calli of Kentucky bluegrass with an Agrobacterium strain and obtained four transgenic lines. She et al. (2005) obtained insect-resistant Kentucky bluegrass plants following transfer of the Bt gene from Bacillus thuringiensis. Later, She et al. (2006) transferred a glucose oxidase-encoding gene to P. pratensis L. via an Agrobacterium-mediated transformation of embryogenic calli.
An efficient regeneration system is critical for developing high numbers of transgenic plants. Previous regeneration systems relied on callus induction from an immature inflorescence (van der Valk et al. 1989), a mature seed (McDonnell and Conger 1984; Boyd and Dale 1986; Pieper and Smith 1988; van der Valk et al. 1989; Griffin and Dibble 1995; Nielsen and Knudsen 1993; Chai et al. 2003; She et al. 2003), a creeping stem, a shoot-tip meristem, hypocotyl, a coleoptile, a leaf, a stem section (Ke and Lee 1996), or a protoplast (Nielsen et al. 1993). However, most genotypes of Kentucky bluegrass cannot easily be made to produce callus tissue or the callus tissues will not easily differentiate into plantlets, thus, limiting the development of genetic transformations using callus tissue as a receptor. Due to the lack of an efficient transformation system, the development of genetic engineering of Kentucky bluegrass has been slow. Thus, the establishment of a high-efficiency, rapid, and genotype-independent transformation system will speed the development of genetic improvement of this species.
In this study, apical meristems of axenic seeding shoot tips were used for the transformation of Kentucky bluegrass (P. pratensis L.). Multiple shoots of Kentucky bluegrass were developed from seedlings as previously described by Hu et al. (2006) and used as apical meristem donors. Various parameters of the transformation system were evaluated, including the concentration of Agrobacterium and the duration of infection and vacuum infiltration. Using this transformation system, the betA gene from Escherichia coli was introduced into three Kentucky bluegrass cultivars. This gene encodes the choline dehydrogenation enzyme and catalyzes the synthesis of betaine in plants growing in adverse environmental conditions, such as high salt, aridity, and low temperature (Sakamoto and Murata 2001).
Materials and methods
Plant materials and production of multiple-shoot clumps
The P. pratensis L. cultivars used in the experiments were Award, NuGlade, and Rugby, the seeds of which were kindly provided by Dr. L. B. Han of the Turf Grass Institute of Beijing Forestry University.
Shoot apices were excised from seedlings germinated from mature seeds of Kentucky bluegrass and used as explants in this experiment. Multiple-shoot clumps were induced from the explants and used as the receptors of Agrobacterium-mediated transformation. Multiple-shoot clumps were produced using the method described by Hu et al. (2006). Seeds of P. pratensis L. were surface-sterilized by immersion in 70% (v/v) ethanol for 1 min, followed by treatment with HgCl2 (0.2%, w/v) for 10–15 min. The seeds were then rinsed five times with sterile water and germinated on moistened filter paper in glass vessels (5 cm in diameter, 8 cm in height) at 25°C in darkness. Vessels were sealed with a polypropylene plastic film-sandwiched Millipore membrane. Ten to fifteen days later, shoot apices with 3–5-mm hypocotyls were excised from seedlings and cultured on initial culture medium, consisting of Murashige and Skoog (MS) medium (Murashige and Skoog 1962) with 0.2 mg L−1 2,4-dichlorophenoxyacetic acid (2,4-D) and 2 mg L−1 6-benzyladenine (BA), in 9-cm Petri dishes for the production of meristematic cell clumps and small calluses. Light and temperature conditions were maintained at 24 ± 2°C under a 14-h photoperiod provided by cool-white fluorescent tubes at a photon flux density of 10–20 μmol m−2 s−1. Twenty days later, the meristematic cell clumps and small calluses were separated from the initial culture medium and transferred to 30 mL of induction medium (MS medium + 2 mg L−1 BA) for the formation of multiple-shoot clumps. Multiple-shoot clumps were then divided into pieces (2–5 shoots each piece) for proliferation in vitro for 15 days on subculture medium (MS medium + 0.07 mg L−1 2,4-D + 2 mg L−1 BA).
All media were suspended with 3% sucrose and 6.5 g L−1 agar, and pH-adjusted to 5.8–6.0, except where otherwise specified.
Bacterial strains and plasmids
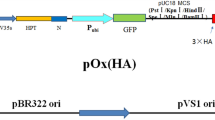
Agrobacterium tumefaciens strain LBA4404 containing vector pCAMBIA1300-P35SbetATnos-als was used in this experiment. The T-DNA regions of the vectors are shown in Fig. 1. The plasmid pCAMBIA1300-P35SbetATnos-als, constructed in our laboratory, consists of the cauliflower mosaic virus 35S promoter driving the target betA gene encoding choline dehydrogenase and the same promoter driving the als gene for herbicide resistance, which acts as the selectable marker.

The T-DNA region of plasmid vector pCAMBIA1300-P35SbetATnos-als used for grass transformation. The betA gene is under the control of the 35S promoter; the als gene, controlled by the same promoter, was used as the selectable marker. LB: left border; RB: right border
A single clone of A. tumefaciens was placed in YEP medium (yeast extract 10 g L−1, peptone 10 g L−1, sodium chloride 5 g L−1, pH 7.0), supplemented with 50 mg L−1 rifamycin and 50 mg L−1 kanamycin, cultured overnight at 28°C, with shaking to mid-log phase. The bacterial cells were then collected by centrifugation at 4,000 rpm for 10 min and then re-suspended in liquid culture medium (MS medium + 2 mg L−1 BA without agar) at a range of concentrations (OD600 = 0.2, 0.4, 0.6, 0.8, 1.0, and 1.2) for use in the following experiments. A 100-μmol L−1 measure of acetosyringone (AS) was added to the bacterial suspensions before infection.
Genetic transformation of shoots and regeneration of plantlets
Vigorous multiple-shoot clumps were cultured on subculture medium for 8–12 days, cut into pieces, and the leaves of the shoots peeled off to expose the shoot-tip meristems. The naked shoot tips were then immersed in the Agrobacterium suspension for some time (0–9 min), supplemented with a negative pressure (0–0.9 × 105 Pa), then lightly blotted dry with filter paper and put onto co-culture medium (MS medium + 0.07 mg L−1 2,4-D + 2 mg L−1 BA, pH 5.2) for 2–4 days. This was followed by 10 days culture on restrain medium (MS medium + 2 mg L−1 BA + 100 mg L−1 cefotaxime) to inhibit the growth of the Agrobacterium. The shoots were then transferred to screen medium (MS medium + 2 mg L−1 BA + chlorsulfuron) and the surviving chlorsulfuron-resistant shoots were selected at 15-day intervals for 45 days. The surviving chlorsulfuron-resistant buds selected were transferred onto subculture medium for recovery. All of the above processes were carried out in darkness. Larger shoots were removed and transferred onto rooting medium (1/2 MS medium + 5 g L−1 agar) after recovery for 10 days. Then, 8–15 days later, the glass vessels were put under sunlight for 1–2 days, after which the roots had attained 2–5 cm in length. Following culture for a further 1–2 days, the plastic films were removed and the plantlets with vigorous roots were finally transplanted into pots containing 6–8 cm of vermiculite over soil. After transplantation, the potted plantlets were irrigated with a solution of 1/2-strength MS inorganic salts every 5 days and with tap water every 2 days.
Sensitivity of Kentucky bluegrass to chlorsulfuron
The herbicide chlorsulfuron, containing 25% active ingredient, was produced by the Shenyang Pesticide Factory. After culture on subculture medium for 10 days, the non-transgenic single shoots of the three breeds of Kentucky bluegrass were transferred to MS medium containing chlorsulfuron at various concentrations (0, 1, 2, 3, 4, 5, 6, or 7 mg L−1) and cultured for three successive 15-day intervals. About 100 single shoots were set for each herbicide treatment and replicated three times. After 45 days, the surviving shoots were counted and the sensitivity of each breed to chlorsulfuron recorded.
Molecular analysis of transformants
Genomic DNA was isolated from the leaves of the larger transformed, transplanted plantlets, following cetyltrimethyl ammonium bromide (CTAB) protocol. A polymerase chain reaction (PCR) analysis was carried out in a 25-μl reaction mixture containing DNA (50 ng), 250 μM of each dNTP, 0.5 μM of each primer, 1 U AmpliTaq DNA polymerase, 2.0 mM MgCl2, and 1× AmpliTaq DNA polymerase buffer (Takara, Dalian, China). Primers used to amplify the betA gene were: primer (1) 5′ CTA CCC GTC TGA AAG ATC C 3′ and primer (2) 5′ CCC ATT TGC CAC AAA ATA TCC 3′. The reaction was initially denatured at 95°C for 5 min, then amplified for 35 cycles of 1 min at 95°C, 1 min at 58.4°C, and 1 min at 72°C, followed by a final extension of 7 min at 72°C. PCR products for the betA gene were 900 base pairs (bp) in length. Primers used to amplify the als gene were: primer (1) GAG GAC ACG CTG AAA TCA CC and primer (2) GCA TCA GGG TTA GCA ACA G. The amplification products were 1,700 bp in length. The reaction procedure was as follows: denaturing at 95°C for 5 min, then amplification for 35 cycles of 1 min at 95°C, 1 min at 56°C, and 1.5 min at 72°C, followed by a final extension of 7 min at 72°C.
Genomic DNA of some PCR-positive plantlets was digested with EcoR I (with only one recognition site in the T-DNA region), transferred from the gel to a nylon membrane using the vacuum method, and hybridized with a DIG-labeled betA (900-bp) probe. Hybridization was performed according to the manufacturer’s instructions for the DIG High Prime DNA Labeling and Detection Starter Kit I (Boehringer).
Experimental design and data analysis
Three replications were performed for each treatment in the optimization of the transformation parameters. The survival rate (%) and transformation frequency (%) of Kentucky bluegrass were defined as follows: survival rate (%) = the number of shoots survived/the number of shoot tips evaluated; transformation frequency (%) = the number of PCR-positive plants/the number of shoot tips evaluated. The data were analyzed using SPSS version 16.0. Analysis of variance (one-way ANOVA) was used to test the statistical significance, and the significance of differences among means was tested using Duncan’s (1955) multiple range test at a significance of P = 0.05.
Results
Production of transformed plantlets
Multiple shoots of Kentucky bluegrass cultivars, Award, NuGlade, and Midnight, were produced from shoot apical meristems (Fig. 2a). After culture in darkness for 8–12 days, multiple-shoot clumps were divided into small clumps, each with 2–3 shoots (Fig. 2b), and the leaves of the shoots were peeled off to expose the shoot-tip meristems. The naked shoot tips were then immersed in a bacterial suspension containing 100 μmol L−1 AS under negative pressure. After co-culture on medium for 2–4 days (Fig. 2c), the shoots were transferred onto restrain medium containing 100 mg L−1 cefotaxime. Ten to fifteen days later, most of the shoots had grown and formed shoot clumps (Fig. 2d). After transfer to selected medium containing 6 mg L−1 chlorsulfuron, the majority of the infected shoot clumps grew slowly and the emergence of secondary buds stopped. The chlorsulfuron-resistant shoots were then transferred to fresh selected medium for a third selection (Fig. 2e). Single shoots 2–3 cm in length were cut from multiple-shoot clumps and induced to root on rooting medium. A rooting frequency of 95–99% was obtained. Vigorous plantlets were transplanted into pots (Fig. 2f).

Agrobacterium-mediated transformation of Poa pratensis L. multiple shoots and regeneration of transformed plantlets: a multiple shoot clumps were produced from Kentucky bluegrass seedling; b shoot tips for infection; c shoot tips with Agrobacterium dots in the basis after being co-cultured for 3 days; d shoot clumps after a 15-day culture on induction medium with cefotaxime; e shoots selected on medium containing chlorsulfuron for 15 days; f transgenic plantlets in pots
Sensitivity of Kentucky bluegrass to chlorsulfuron and selection of herbicide-resistant plantlets
After culture on subculture medium for 10 days, the non-transgenic single shoots of the three cultivars of Kentucky bluegrass were transferred to the MS medium with chlorsulfuron (0–7 mg L−1). The effects of the herbicide chlorsulfuron were evaluated to determine the appropriate selection dose. The growth rate of the shoots clearly declined in the presence of chlorsulfuron; the leaves of some plantlets turned yellow and some of the plantlets turned entirely yellow and died. When the chlorsulfuron concentration was 6 mg L−1, the survival rate of the three cultivars of Kentucky bluegrass (Award, NuGlade, Rugby) was between 7 and 9%, and most of the non-transformed cells and plantlets died as a result. Therefore, 6 mg L−1 chlorsulfuron was used as the selective concentration in testing the transformation rates of Kentucky bluegrass (Fig. 3).

Effects of chlorsulfuron concentration on the survival rate of shoots The survival rate of shoots was defined as follows: survival rate (%) = the number of shoots survived/the number of shoot tips evaluated. About 100 single shoots were set for each herbicide treatment and repeated three times
Molecular analysis of transformants
Genomic DNA isolated from the surviving transformed and non-transformed plantlets were used as PCR templates. PCR assay showed that the specific bands of the betA and als genes were amplified in some of the putative transformed shoots, but not in the non-transformed plantlets (Fig. 4a, b). Southern hybridization was used to confirm the stable integration of the foreign gene. Genomic DNA of PCR-positive plants was digested with EcoRI and the DIG-labeled betA gene probe was used to make a hybrid. Bands of the betA gene were observed in the transgenic plants (Fig. 4c), indicating that the betA gene had been integrated into the genomes of the transgenic plants.

Polymerase chain reaction (PCR) and Southern blot analysis of transgenic plants: a PCR analysis of transgenic plants showing amplification of a 900-bp fragment (betA), 1: DL2000 Marker, 2: plasmid, 3: non-transformed plant (Award), 4: H2O; 5, 6, 7, 8, 9: transgenic plants; b PCR analysis of transgenic plants showing amplification of a 1,700-bp fragment (als), 1, 2, 3, 4: transgenic plants, 5: non-transformed plant (Award), 6: plasmid, 7: DL2000 Marker; c Southern blot analysis of transgenic plants (betA), 1: λDNA/Hind III Marker, 2: plasmid DNA, 3: non-transformed plant (Award), 4, 5, 6, 7: transgenic plants
Optimization of the genetic transformation
Agrobacterium suspension concentration
In order to analyze the effects of the concentration of Agrobacterium suspension on the percentage of chlorsulfuron-resistant shoots, small shoots were immersed in a range of concentrations (OD600 = 0.2, 0.4, 0.6, 0.8, 1.0, and 1.2) of Agrobacterium suspension containing 100 μmol L−1 AS for 5 min at a negative pressure of 0.5 × 105 Pa. The results showed that the survival rate and transformation frequency of the shoots at first increased with increasing concentration of bacterial cells, and then decreased at higher bacterial cell densities. The highest percentage of PCR-positive plants (1.42%) was obtained when OD600 was 0.6 (Table 1).
The density of Agrobacterium in the inoculation medium is one of the critical factors in the shoot-tip transformation system (Li et al. 2007). The Agrobacterium density has often been set to between 0.5 and 1.0 OD600 in previous Agrobacterium-mediated transformations of the embryogenic calli of grasses (Hu 2005; Zhao et al. 2005; Gao et al. 2008; Ge et al. 2007; Wang et al. 2007; Wu et al. 2006; Kong and Zhao 2008; Qian et al. 2006). Dutt et al. (2007) co-cultured shoot-tip explants of Vitis vinifera L. with Agrobacterium cells at a density of OD600 = 0.8. Gu et al. (2008) obtained the best transformation frequency (3.2%) of winter jujube (Zizyphus jujuba Mill.) using Agrobacterium inoculum at OD600 = 0.8. Our study suggested that Agrobacterium density OD600 = 0.6 was the most effective density for the transformation of Kentucky bluegrass. No PCR-positive plants were obtained when the inoculation density of Agrobacterium was higher than OD600 = 1.0 or lower than OD600 = 0.4. The likely explanation for this is that, when the Agrobacterium density was too low (OD600 = 0.2), few Agrobacterium cells were absorbed by the tissue surface and few entered the shoot-tip cells, whilst when the OD600 was above 1.0, most shoots became necrotic due to the flourishing growth of Agrobacterium, and the resistant shoot percentage (7.05%) decreased significantly.
Duration of infection and co-culture
The effects of infection duration were also investigated in an Agrobacterium suspension of OD600 = 0.6 containing 100 μmol L−1 AS. When the infection duration was about 1–3 min, few shoots survived after selection. A significantly increased frequency of transformation (1.42%) was obtained when shoot-tip explants were incubated for 5 min. Following infection for 9 min, the Agrobacterium grew luxuriantly and the majority of the shoots necrotized, resulting in a decline in the percentage of resistant shoots (Table 2).
Previous studies have reported that the transformation frequency in woody plants was influenced with explants inoculation with Agrobacterium for 1–30 min (Costa et al. 2002; Yu et al. 2002; Chen et al. 2006; Kong and Zhao 2008; Wu et al. 2006; Hu 2005). The duration of the exposure interval to Agrobacterium cells is a critical step in the process of Agrobacterium-mediated transformation, since absorption of Agrobacterium, and transference and integration of T-DNA are all completed during this period. Thus, if the duration of co-cultivation was too short, transference of T-DNA would not be completed. If the exposure was too long, the eugenic Agrobacterium caused damage to the plant cells and inhibited regeneration (Hiei et al. 1994; Smith and Hood 1995). The results in our study suggested that 5 min was the most effective infection period for Kentucky bluegrass when the Agrobacterium density was OD600 = 0.6, the concentration of AS was 100 μmol L−1, and the negative pressure was 0.5 × 105 Pa.
Infection under negative pressure
The percentage of chlorsulfuron-resistant shoots and the transformation frequency of Award under a negative pressure of 0, 0.3 × 105, 0.5 × 105, 0.7 × 105, and 0.9 × 105 Pa for 5 min for shoot tips infected with Agrobacterium suspension of OD600 = 0.6 containing 100 μmol L−1 AS is shown in Table 3. The highest transformation percentage was obtained under a negative pressure of 0.5 × 105 Pa, being 1.42% compared to 0.69% without negative pressure. When the negative pressure was raised, the shoot meristem and the Agrobacterium could be damaged by either higher negative pressure or longer negative pressure duration, leading to a decrease in resistant percentage.
Vacuum infiltration has been widely used in Agrobacterium-mediated transformation of plants. Quan et al. (2003) found that the transformation efficiency of embryogenic calli of elite maize inbred lines Ye 515 and Ye 502 increased from 6.3 and 4.5% to 8.7 and 7.8%, respectively, following vacuum infiltration at 0.5 × 105 Pa for 5 min. Yang et al. (2005) reported that the application of a negative pressure during transformation enhanced the transformation frequency of sugar beet. Gu et al. (2008) obtained a 1.63-fold increase (5.2%) in the transformation frequency of winter jujube (Z. jujuba Mill.) with a negative pressure of 0.5 × 105 Pa in comparison to that obtained at atmospheric pressure. Vacuum infiltration has also been used in Arabidopsis thaliana (Bechtold and Pelletier 1998). It has been suggested that a negative-pressure environment created by a vacuum pump helps the infiltration of Agrobacterium cells and the transference of foreign genes into plant cells. Negative pressure would reduce the spaces between plant cells, aiding the entry of suspended Agrobacterium cells into plant cells and subsequent T-DNA transference.
Discussion
Genetic improvement of Kentucky bluegrass (P. pratensis L.) via transformation methods depends mainly on developing a favorable receptor system. Establishment of an efficient in vitro regeneration system plays an important role in the transgenic transformation. However, some problems have persisted in the genetic transformation of Kentucky bluegrass, such as the difficulty in promoting plant regeneration, the high frequency of somatic variance, the intense genotype-dependence, and the long transformation period required, all of which hinder the genetic improvement of Kentucky bluegrass (van der Valk et al. 1989; Griffin and Dibble 1995; Ke and Lee 1996; She et al. 2003). In our work, the shoot-tip meristem was used as the target tissue to overcome the genotype obstacles, the reluctance to form calluses, and to differentiate buds from calluses. We established a regeneration system using multiple-shoot clumps of Kentucky bluegrass and obtained a regeneration frequency of over 70% for three different cultivars (Hu et al. 2006). Furthermore, transgenic plants produced from the shoot-tip meristem retained their original characteristics due to the avoidance of subculture callus production.
In this study, we have optimized the transformation conditions and established an efficient genetic transformation system. In this system, shoot tips were lacerated and the cells were sensitive to Agrobacterium infection. In addition, the wounded surface of the shoot tips produced a large amount of phenolic compounds, which were useful for the genetic transformation of the plant cells. Using this system, we introduced the betA gene (Landfald and Strøm 1986) into Kentucky bluegrass and created breeding stock with drought and salt tolerance potential that will be tested in our future work.
In recent years, more and more studies have reported the use of multiple-shoot apical meristems as the transformation receptors (Lv et al. 2004; Zimmerman and Scorza 1996), and great progress has been made. Favorable results have been reported in beets (Yang et al. 2005), corn (Zhang et al. 2005; Gould et al. 1991; Sairam et al. 2003), cotton (Lv et al. 2004), Vitis (Dutt et al. 2007), winter jujube (Gu et al. 2008), soybean (Liu et al. 2004), and Vigna (Saini and Jaiwal 2005) etc., indicating great promise for this technique. In this work, multiple-shoot clumps were used as the receptor material and we successfully optimized the various transformation parameters, such as the density of Agrobacterium and duration of infection, and explored the use of vacuum infiltration to increase transformation frequencies. The results indicate that the transformation frequency and process efficiency can be improved. However, the genetic transformation system also has a disadvantage. In this system, transformed cells produced by Agrobacterium-mediated infection only retained a small part of the meristem after co-culture, and divided repeatedly on the selection medium to form new shoot meristems. This resulted in some transgenic chimeric plants in generation T0, in which not every cell of the reproductive organ was formed from transgenic ones. Nevertheless, chimeric transgenic plants are still valuable for breeding.
Abbreviations
- CTAB:
-
Cetyltrimethyl ammonium bromide
- BA:
-
6-benzyladenine
- 2,4-D :
-
2,4-dichlorophenoxyacetic acid
- AS:
-
Acetosyringone
References
Akerberg E (1979) Apomictic and sexual seed formation in Poa pratensis L. Hereditas 25:359–371
Bashaw EC (1980) Apomixis and its application in crop improvement. In: Fehr WR, Hadley HH (eds) Hybridization of crop plants. Madison Press, Wisconsin, pp 455–633
Bechtold N, Pelletier G (1998) In planta Agrobacterium-mediated transformation of adult Arabidopsis thaliana plants by vacuum infiltration. Methods Mol Biol 82:259–266
Boyd LA, Dale PJ (1986) Callus production and plant regeneration from mature embryos of Poa pratensis L. Plant Breed 97:246–254
Chai BF, Liang AH, Wang W, Hu W (2003) Agrobacterium-mediated transformation of Kentucky bluegrass. Acta Bot Sinica 45(8):966–973, in English with Chinese abstract
Chen YQ, Lu LT, Deng W, Yang XY, McAvoy R, Zhao DG, Pei Y, Luo KM, Duan H, Smith W, Thammina C, Zheng XL, Ellis D, Li Y (2006) In vitro regeneration and Agrobacterium-mediated genetic transformation of Euonymus alatus. Plant Cell Rep 25:1043–1051
Costa MGC, Otoni WC, Moore GA (2002) An evaluation of factors affecting the efficiency of Agrobacterium-mediated transformation of Citrus paradise (Macf.) and production of transgenic plants containing carotenoid biosynthetic genes. Plant Cell Rep 21:365–373
Duncan DB (1955) Multiple range and multiple F tests. Biometrics 11:1–42
Dutt M, Li ZT, Dhekney SA, Gray DJ (2007) Transgenic plants from shoot apical meristems of Vitis vinifera L. “Thompson Seedless” via Agrobacterium-mediated transformation. Plant Cell Rep 26:2101–2110
Gao CX, Jiang L, Folling M, Han LB, Nielsen KK (2006) Generation of large numbers of transgenic Kentucky bluegrass (Poa pratensis L.) plants following biolistic gene transfer. Plant Cell Rep 25:19–25
Gao CX, Long DF, Lenk I, Nielsen KK (2008) Comparative analysis of transgenic tall fescue (Festuca arundinacea Schreb.) plants obtained by Agrobacterium-mediated transformation and particle bombardment. Plant Cell Rep 27(10):1601–1609
Ge YX, Cheng XF, Hopkins A, Wang ZY (2007) Generation of transgenic Lolium temulentum plants by Agrobacterium tumefaciens-mediated transformation. Plant Cell Rep 26:783–789
Gould J, Devey M, Hasegawa O, Ulian EC, Peterson G, Smith RH (1991) Transformation of Zea mays L. using Agrobacterium tumefaciens and the shoot apex. Plant Physiol 95:426–434
Grazi F, Umaerus M, Åkerberg E (1961) Observations on the mode of reproduction and the embryology of Poa pratensis. Hereditas 47:489–541
Griffin JD, Dibble MS (1995) High-frequency plant regeneration from seed-derived callus cultures of Kentucky bluegrass (Poa pratensis L.). Plant Cell Rep 14:721–724
Griffin J, Zemetra R (1999) Genetic transformation of Kentucky bluegrass. In: Dibble MS, Gu SZ, Griffin JD (eds) Progress report: transgene expression in Kentucky bluegrass: GUS and BAR. In Agronomy Abstracts (p 98). ASA, Madison, Wisconsin
Gu XF, Meng H, Qi G, Zhang JR (2008) Agrobacterium-mediated transformation of the winter jujube (Zizyphus jujuba Mill.). Plant Cell Tiss Org Cult 94:23–32
Ha CD, Lemaux PG, Cho MJ (2001) Stable transformation of a recalcitrant Kentucky bluegrass (Poa pratensis L.) cultivar using mature seed-derived highly regenerative tissues. In Vitro Cell Dev Biol Plant 37(1):6–11
Hanna WW, Bashaw EC (1987) Apomixis: its identification and use in plant breeding. Crop Sci 27(6):1136–1139
Hiei Y, Ohta S, Komari T, Kumashiro T (1994) Efficient transformation of rice (Oryza saliva L.) mediated by Agrobacterium and sequence analysis of the boundaries of the T-DNA. Plant J 6:271–282
Hu FR (2005) Optimization of Agrobacterium-mediated transformation of tall fescue (Festuca arundinacea Schreb.). Mol Plant Breed 3(3):375–380, in Chinese
Hu XR, Yang AF, Zhang KW, Wang J, Zhang JR (2006) Optimization of in vitro multiple shoot clump induction and plantlet regeneration of Kentucky bluegrass (Poa pratensis). Plant Cell Tissue Organ Cult 84:89–98
Ke SQ, Lee CW (1996) Plant regeneration in Kentucky bluegrass (Poa pratensis L.) via coleoptile tissue cultures. Plant Cell Rep 15:882–887
Ke SQ, Lee CW, Cheng ZM (1996) Genetic transformation of Kentucky bluegrass with rolC gene. HortSci 31:616
Kong Z, Zhao DG (2008) The combination of CHI and AFP genes introduced into ryegrass mediated by Agrobacterium. Mol Plant Breed 6(2):281–285, in Chinese
Landfald B, Strøm AR (1986) Choline-glycine betaine pathway confers a high level of osmotic tolerance in Escherichia coli. J Bacteriol 165(3):849–855
Li ZN, Fang F, Liu GF, Bao MZ (2007) Stable Agrobacterium-mediated genetic transformation of London plane tree (Platanus acerifolia Willd.). Plant Cell Rep 26:641–650
Liu HK, Yang C, Wei ZM (2004) Efficient Agrobacterium tumefaciens-mediated transformation of soybeans using an embryonic tip regeneration system. Planta 219:1042–1049
Lv SL, Yin XY, Zhang KW, Zhang JR (2004) Agrobacterium-mediated transformation of shoot apex of cotton and production of transgenic plants carrying betA gene. High Tech Lett 14:20–25, in Chinese
Ma ZH, Zhang YF, Xu CX, Chen WJ, Yin HH, Kuai BK (1999) Tissue culture and genetic transformation of Kentucky bluegrass (Poa pratensis) via microprojectile bombardment. J Fudan Univ (Natural Science) 38(5):540–544, in Chinese
McDonnell RE, Conger BV (1984) Callus induction and plantlet formation from mature embryo explants of Kentucky bluegrass. Crop Sci 24:573–578
Meyer W, Zhang G, Lu S, Chen S, Chen TA, Funk R (2000) Transformation of Kentucky bluegrass (Poa pratensis L.) with betaine aldehyde dehydrogenase gene for salt and drought tolerance. Annual Meeting Abstracts, American Society of Agronomy, Crop Science Society of America, Soil Science Society of American (ASA/CSSA/SSSA), Minneapolis, Minnesota, November 2000, p 167
Murashige T, Skoog F (1962) A revised medium for rapid growth and bioassays with tobacco tissue cultures. Physiol Plant 15:473–497
Nielsen KA, Knudsen E (1993) Regeneration of green plants from embryogenic suspension cultures of Kentucky bluegrass (Poa pratensis L.). J Plant Physiol 141:589–595
Nielsen KA, Larsen E, Knudsen E (1993) Regeneration of protoplast-derived green plants of Kentucky bluegrass (Poa pratensis L.). Plant Cell Rep 12:537–540
Pieper MA, Smith MAL (1988) A whole plant microculture selection system for Kentucky bluegrass. Crop Sci 28:611–614
Qian HF, Shaukat A, Hong L, Xu H (2006) Establishment of genetic transformation system via Agrobacterium in tall fescue cultivar. J For Res 17(3):238–242
Quan RD, Shang M, Zhang JR (2003) Optimization of transformation conditions in maize inbred line calluses via Agrobacterium tumefaciens. J Plant Physiol Mol Biol 29(3):245–250, in Chinese
Saini R, Jaiwal PK (2005) Transformation of a recalcitrant grain legume, Vigna mungo L. Hepper, using Agrobacterium tumefaciens-mediated gene transfer to shoot apical meristem cultures. Plant Cell Rep 24:164–171
Sairam RV, Parani M, Franklin G, Lifeng Z, Smith B, MacDougall J, Wilber C, Sheikhi H, Kashikar N, Meeker K, Al-Abed D, Berry K, Vierling R, Goldman SL (2003) Shoot meristem: an ideal explant for Zea mays L. transformation. Genome 46:323–329
Sakamoto A, Murata N (2001) The use of bacterial choline oxidase, a glycinebetaine-synthesizing enzyme, to create stress-resistant transgenic plants. Plant Physiol 125(1):180–188
She JM, Zhang BL, Chen ZY, Ni WC (2003) A study on the technique of plant regeneration from mature seed embryo of Kentucky bluegrass in vitro. Acta Agrestia Sin 11(1):58–62, in Chinese
She JM, Liang LF, Zhang BL, He XL, Chen ZY, Ni WC (2005) Acquirement of Bt transgenic plants by Agrobacterium tumefaciens in Kentucky bluegrass (Poa pratensis L.). Jiangsu J Agr Sci 21(2):102–105, in Chinese
She JM, Zhang BL, Liang LF, He XL, Yao S, Chen ZY, Ni WC (2006) Acquirement of transgenic plants with glucose oxidase gene in Kentucky bluegrass (Poa pratensis L.). Jiangsu J Agr Sci 22(3):217–221, in Chinese
Smith RH, Hood EE (1995) Agrobacterium tumefaciens transformation of monocotyledons (review). Crop Sci 35(2):301–309
van der Valk P, Zaal MACM, Creemers-Molenaar J (1989) Somatic embryogenesis and plant regeneration in inflorescence and seed derived callus cultures of Poa pratensis L. (Kentucky bluegrass). Plant Cell Rep 7:644–647
Wang ZY, Ge Y (2006) Recent advances in genetic transformation of forage and turf grasses. In Vitro Cell Dev Biol Plant 42:1–18
Wang YL, Ye XG, Dong F, Qiao WH, Tao LL, Li XL, Xing XU (2007) Agrobacterium-mediated transformation of tall fescue and perennial ryegrass. China Biotech 27(1):22–27, in Chinese
Wu GT, Chen JQ, Hu ZH, Lang CX, Chen XY, Wang FL, Jin W, Xia YW (2006) Production of transgenic tall fescue plants with enhanced stress tolerance by Agrobacterium tumefaciens-mediated transformation. Scientia Agri Sin 38(12):2395–2402, in Chinese
Xin JN, Han LB, Liu J, Han XB (2006) Transformation of Kentucky bluegrasss (Poa pratensis L.) by particle bombardment. China Biotech 26(8):10–14, in Chinese
Yang AF, Duan XG, Gu XF, Gao F, Zhang JR (2005) Efficient transformation of beet (Beta vulgaris) and production of plants with improved salt-tolerance. Plant Cell Tissue Organ Cult 83:259–270
Yu CH, Huang S, Chen CX, Deng ZN, Ling P, Gmitter FG Jr (2002) Factors affecting Agrobacterium-mediated transformation and regeneration of sweet orange and citrange. Plant Cell Tissue Organ Cult 71:147–155
Zhang JR, Li XH, Quan RD, Zhang QW, Shang M, Yang AF, Li GS (2005) Method for creating transgenic receptor system of corn and application. China Patent 00110842.5, 29 October 2003
Zhao JS, Zhi DY, Xue ZY, Xia GM (2005) Research on Festuca arundinacea transformation mediated by Agrobacterium tumefaciens. Acta Genet Sin 32(6):579–585, in Chinese
Zimmerman TW, Scorza R (1996) Genetic transformation through the use of hyperhydric tobacco meristems. Mol Breed 2:73–80
Acknowledgments
We would like to thank Dr. L. B. Han at the Turfgrass Institute of Beijing Forestry University for providing the seeds of Kentucky bluegrass. This research was supported by the National Project for Transgenic Plant Research and Industrialization of China (2008ZX08005-4), the Cotton Improved Variety Project of Shandong Province (2009GG10009010), and the High-Tech Research and Development Program of China (2007AA091701). We would also like to thank International Science Editing for their assistance in the language editing of the manuscript.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Zhang, K., Wang, J., Hu, X. et al. Agrobacterium-mediated transformation of shoot apices of Kentucky bluegrass (Poa pratensis L.) and production of transgenic plants carrying a betA gene. Plant Cell Tiss Organ Cult 102, 135–143 (2010). https://doi.org/10.1007/s11240-010-9713-9
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11240-010-9713-9