
Abstract
Despite their similar morphology, banana and maize shoot tips responded strikingly different with respect to the in vitro formation of homogeneous multiple shoot clusters. While up to 50 small shoots per maize explant could be induced within 1 month, zero to one additional shoot formed starting from a banana shoot tip. Subsequently, banana shoot tips were subjected to different combinations of five cytokinins (0–100 μM) and five auxins (0–5 μM). The cytokinins thidiazuron and benzylaminopurine stimulated multiplication to a higher extent compared to zeatin, kinetin and isopentenyl adenine. The addition of indoleacetic acid, naphthalene acetic acid or indolebutyric acid to cytokinin containing medium did not affect the in vitro response. In contrast, 2,4-dichlorophenoxyacetic acid (1 and 5 μM) and a higher concentration of picloram (5 μM) had a detrimental effect on shoot formation and resulted in explant death and globule development. When small (0.1 cm) shoot tips were grown on cytokinin medium without an auxin source, the average number of shoots was generally two to three times lower compared to bigger (0.5 cm) shoot tips. Based on our experience in maize and this large-scale study with banana shoot tips, we conclude that banana is extremely recalcitrant towards adventitious shoot formation. This recalcitrance could not be overcome by any of the 173 different plant growth regulator combinations tested. In vitro multiplication of banana thus appears solely restricted to axillary shoot formation.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Bananas and plantains (Musa species) are among the five major food commodities in the developing world. Since most banana cultivars produce seedless fruits, they are vegetatively multiplied both in vivo as well as in vitro. Starting from individual shoot tips, vegetative propagation in vitro is routinely stimulated on semi-solid MS (Murashige and Skoog 1962) medium supplemented with 10 μM benzylaminopurine (BAP) and 1 μM indoleacetic acid (IAA) (standard banana multiplication medium p5) (Vuylsteke and De Langhe 1985; Van den Houwe et al. 1995). Subsequent culture cycles of multiple shoots on a medium with 10 times higher BAP concentration and continuous selection for clusters of small (less than 3 mm) shoots induce a gradual increase in the ratio of meristematic versus more differentiated corm and leaf tissue present at the top layers of the culture (Dhed’a et al. 1991). The amount of meristematic tissue per surface unit determines the quality of these so-called multiple meristem cultures (MMC) and their suitability for further applications. MMC can be successfully cryopreserved for the safe storage of Musa germplasm (Panis et al. 2005). In addition, ‘scalps’ (1–3 mm top layer of the MMC) are a good source for embryogenic callus, which in turn provide embryogenic cell suspensions (Schoofs 1999; Strosse et al. 2006). While research is ongoing on genetic transformation of MMC directly (Remy et al. personal communication), embryogenic cell suspensions remain the preferred target material for generation of transgenic plants because of their high transformation efficiency and lower chance for chimaerism (Sági et al. 1995; Schoofs et al. 1999).
A prerequisite for genetic engineering is the development of a highly efficient and reproducible in vitro regeneration system permitting the production of non-chimaeric plants from transformed cells. This is not straightforward in monocots, since totipotent tissues are far more limited compared to the less recalcitrant dicots (Vasil 1990; Benson 2000). A review from 2005 (Sticklen and Oraby 2005), highlights the general availability of apical meristem regeneration systems to stably transform cereal crops including maize, oat, sorghum, barley, millet and wheat. The morphogenetic plasticity of cereal meristem cells was first demonstrated in maize (Zhong et al. 1992). Variation in the cytokinin: auxin (in this case BAP: 2,4-D) ratio of the culture medium resulted in the production of multiple shoots from maize shoot tips and/or somatic embryos. Adventitious shoot formation was shown to play a key role in the development of uniformly transformed plants from these tissues (Zhang et al. 1999).
The origin of multiple shoot cultures in banana, however, remains unclear. Based on their late initiation and unusual position Barker and Steward (1962) excluded the formation of vegetative buds in leaf axils and suggested the adventitious origin of lateral buds in Musa. Fisher (1978) compared the development of lateral buds in Musa with closely related monocots. He concluded that the leaf-opposed position of vegetative buds is a generic feature of Musa, being both morphologically and developmentally unique within the order of Zingiberales. Based on their determined position and development from immature and meristematic cells, Fisher (1978) proposed that vegetative buds in Musa are not adventitious in origin although they are initiated later than the typical axillary buds. In later publications on multiple shoot formation in banana, a debate or confusion exists between the axillary versus adventitious origin of shoots (Vuylsteke and De Langhe 1985; Banerjee et al. 1986; Cronauer and Krikorian 1987; Okole and Schulz 1996; Zaffari et al. 2000; Li et al. 2006).
The present study was performed to gain knowledge on the nature of multiple shoots in banana. For this purpose, we first applied the in vitro method developed for maize by Zhong et al. (1992) on maize and banana shoot tips. Subsequently, we investigated the influence of explant size and a broad range of plant growth regulators (173 combinations of five cytokinins and five auxins, each at various concentrations) on the development of banana shoot tips. The elucidation of the origin (axillary vs. adventitious) of multiple shoots in banana is important in view of their potential applications towards genetic engineering (i.e. direct transformation of multiple meristem cultures or indirect via production of transformation competent embryogenic cell suspensions) but also in view of the occurrence of somaclonal variation after micropropagation.
Material and methods
In vitro response of maize and banana shoot tips
The maize inbred line B73 was used to reproduce the production of multiple in vitro shoot cultures from shoot tips according to Zhong et al. (1992). Mature seeds (obtained from the Cereal Research Non-Profit Company, Szeged, Hungary) were surface-sterilised in 70% ethanol, washed once with sterile water and soaked for 30 min in 2.6% sodium hypochlorite containing 0.1% Tween 20. The surface-sterilised seeds were rinsed three times with sterile distilled water prior to germination on MS based medium in glass tubes in the dark at 27°C. Seedlings that reached a length of 3–5 cm within 1 week after inoculation were used for dissection of shoot tips (Fig. 1a).

Morphological appearance of maize (a, c, e, g) and banana (b, d, f, h) shoot cultures; a, b: rooted in vitro plant, bar = 3.9 mm; c, d: explants with enlargement at the level of the shoot apical meristem (white arrow), bar = 2.7 mm; e, f: 0.5 cm explant consisting of the apical dome covered by three to five leaf primordia and a few millimetres of corm tissue beneath the apical dome, bar = 770 μm; g, h: small explant with the apical dome (black arrow) only partly covered by one leaf primordium and only a minimal amount of corm tissue beneath the apical dome, bar = 300 μm
Vegetatively propagated banana (cv. ‘Williams’, AAA group, ITC.0365) shoots were obtained from the International Transit Centre at K.U.Leuven (van den Houwe et al. 1995). Since the initial explants need to be as uniform as possible, shoot tips were not isolated directly from these multiple shoots, but from individually grown plants. For this, vegetatively propagated shoots were separated in individual shoots and grown in glass tubes during three monthly culture cycles on semi-solid MS basal medium supplemented with 10 mg/l ascorbic acid, 30 g/l sucrose, 0.5 g/l active charcoal and solidified with 3 g/l Gelrite® (MSac). Cultures were stored at 27°C under continuous light of 50 μE m−2 s−1. At the onset of a subculture period, roots were removed and leaves cut back to 0.5 cm above the apical meristem. Individual rooted plants of ‘Williams’ that reached a length of about 10 cm at the end of the third subculture period on MSac were used for the isolation of shoot tips (Fig. 1b).
Shoot tips were prepared from maize seedlings and rooted banana plants by removing roots, cutting off leaves at 0.5 cm above the apical meristem followed by gradual removal of leaf sheaths closely encircling each other (‘pseudo stem’ tissue). The bigger explant consisted of the apical dome, fully covered by three to five leaf primordia and 2–3 mm of young corm tissue (true stem, directly beneath the apical dome (0.5 cm explant, Fig. 1e, f). Smaller explants were obtained by further dissection until the apical dome became visible and was partly covered by one leaf primordium (0.1 cm explant, Fig. 1g, h).
In maize as well as in banana, 12 bigger shoot tips (0.5 cm explants, see above) were cultured on corn shoot multiplication (CSM) medium according to Zhong et al. (1992). This CSM medium consisted of MS salts and vitamins, 30 g/l sucrose, 500 mg/l casein hydrolysate, 10 μM BAP and was solidified with 7 g/l Plantagar®. The pH of the culture medium was adjusted to 6.2 prior to autoclaving for 20 min at 120°C. In each Petri dish, three explants were placed horizontally and partly buried in the medium. Four weeks after inoculation, leaf primordia were cut back in six of the 12 explants. In addition, six 0.5 cm shoot tips and six 0.1 cm shoot tips dissected from maize as well as banana plants were grown on basal CSM medium supplemented with 10 μM BAP and 2.5 μM 2,4-dichlorophenoxyacetic acid (2,4-D). All cultures were grown for 3 months at 27°C in the dark. At 4-week intervals, cultures were transferred to fresh medium including fresh plant growth regulators (PGR). The in vitro response (i.e., explant death, outgrowth or multiplication of shoots, formation of globules) was observed every 2 weeks. The entire experiment was performed twice.
Large-scale study on the development of banana shoot tips subjected to various plant growth regulator combinations
The effect of type and concentration of cytokinins
To eliminate the influence of the in vitro history on the development of banana shoot tips, ‘Williams’ plants went through a 9-month cycle in the greenhouse prior to re-initiation in vitro. From each of the greenhouse plants a cube of 1–2 cm3 containing the apical meristem was excised. These explants were surface-sterilised by dipping in 70% ethanol for 10 s and then stirring for 20 min in 2% sodium hypochlorite solution containing 0.1% Tween 20. Subsequently, the plant material was rinsed three times in sterile distilled water for 10 min. After surface-sterilisation, shoot tips of 0.3–0.5 cm in length were aseptically dissected. Multiple shoot cultures were maintained on p5 medium consisting of MS basal salts and vitamins supplemented with 10 μM BAP, 1 μM IAA, 10 mg/l ascorbic acid and 30 g/l sucrose. This banana propagation medium p5 was solidified with 3 g/l Gelrite®. Plant material was grown at 27°C under continuous light of 50 μE m−2 s−1. Two months after re-initiation in vitro, 200 individual plants were isolated from the multiple shoot clusters and cultured for three subsequent cycles on MSac. Small shoot tips (0.1 cm sized, see above) were dissected from ‘Williams’ rooted in vitro plants and used as initial explants to determine the influence of five cytokinins [BAP, kinetin, isopentenyladenine (2iP), zeatin, and thidiazuron (TDZ)] each at three concentrations (1, 10 and 100 μM) added to the basal CSM medium. Inoculation of 0.1 cm sized shoot tips on CSM medium devoid of plant growth regulators served as a control. Per treatment, six shoot tips were equally divided over two Petri dishes and cultured during six monthly cycles at 27°C in the dark. At the end of each culture cycle, plant material was observed for morphological characteristics such as explant death, blackening, number and length of shoots, presence of roots, formation of corm tissue, callus and globules. Without additional manipulation, structures were transferred to fresh culture medium with also PGR refreshed at 4-weekly intervals. The entire experiment was performed twice. For each treatment, the number of shoots per ‘initial’ and per ‘surviving’ explants was counted. ‘Initial’ explants are the total of non-surviving plus surviving explants, i.e. forming single and multiples shoots. This distinction was made to draw attention to the toxicity of treatments. Hence, the average number of shoots per surviving explant is higher than the average number of shoots per initial explant whenever there is toxicity. Standard errors for calculated average number of shoots are presented for surviving explants.
The effect of explant size and cytokinins in combination with the auxin 2,4-D
The bigger banana shoot tips (0.5 cm explants, see above) of ‘Williams’ were subjected to 105 plant growth regulator combinations (BAP, kinetin, 2iP, zeatin and TDZ, each at a concentration of 0, 1, 5, 10 and 50 μM in combination with 2,4-D at a concentration of 0, 0.1, 0.5, 1 and 5 μM) supplemented to the basal CSM medium. The influence of the different cytokinins in combination with various concentrations of 2,4-D was also explored for smaller banana shoot tips (0.1 cm explants, see above), however, with a 10 μM cytokinin concentration only. Shoot tips of different sizes grown on CSM medium devoid of plant growth regulators served as a control. For the bigger as well as the smaller shoot tips, six identically treated explants were divided over two Petri dishes and cultured during six months at 27°C in the dark. Culture conditions, monthly refreshment of culture medium (PGR inclusive), observation of morphological characteristics, calculation of the average number of shoots per ‘initial’ and per ‘surviving’ explant and corresponding standard errors was done as described above. The experiment with the bigger shoot tips was performed twice, while the experiment with the smaller shoot tips as initial explant was performed only once.
The effect of type and concentration of auxins in combination with the cytokinin TDZ
Bigger (0.5 cm sized, see above) ‘Williams’ shoot tips were subjected to the following plant growth regulator combinations in the basal CSM medium: IAA, naphthalene acetic acid (NAA), indolebutyric acid (IBA), picloram (Pic) and 2,4-D each at a concentration of 0, 0.1, 0.5, 1 and 5 μM, respectively, in combination with TDZ at a concentration of 0, 1, 5 and 10 μM, respectively. Inoculation of 0.5 cm sized shoot tips on CSM medium devoid of plant growth regulators served as control. Per treatment six shoot tips were equally divided over two Petri dishes and cultured during six months in the dark. Structures were transferred to fresh culture medium with also PGR being refreshed at 4-weekly intervals. Observations were executed as described above. The entire experiment was performed twice.
Results
In vitro response of maize versus banana shoot tips
Maize and banana explants revealed a similar morphology (Fig. 1). In maize as well as banana, the position of the shoot apical meristem can be deduced from the distinct enlargement just above the junction of corm and leaf tissue (Fig. 1c, d). Noticeable was the more conical shape of the maize apical dome compared to the more flattened apical dome in banana (Fig. 1g vs. h).
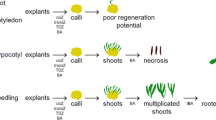
Two weeks after initiation of 0.5 cm maize shoot tip cultures on CSM medium containing 10 μM BAP, 80% of developing cultures showed an increase in volume at about 1 cm above the base of the outer leaf sheaths. After removal of the outer leaves in 4-week old cultures, clusters of up to 50 small (<3 mm) almost transparent shoots became visible (Fig. 2a–c. These multiple shoot clusters were located at the end of an elongated internode. Further culture of individual shoot clusters resulted in excessive multiplication. We estimate that a few 100 shoots formed on each initial explant at the end of the second culture cycle. The addition of 2,4-D resulted in about 30% more shoots formed per initial explant. Explant death was not encountered for any of the 0.5 cm sized maize shoot tips.

Shoot cultures of maize (a, b, c) and banana (d, e, f) one month after inoculation of 0.5 cm sized shoot tips on corn shoot multiplication (CSM) medium (a, d, e) before and (b, c, f) after removal of outer leaves; (a) bar = 0.5 cm; (b) bar = 1.6 mm; (c) bar = 160 μm; (d), (e) bar = 2.5 mm; (f) bar = 830 μm
The same protocol with also 0.5 cm sized banana shoot tips resulted after 1 month in the outgrowth of only single shoots (40% of initial explants, Fig. 2d) or in minimal multiplication (one secondary shoot for 60% of initial explants, Fig. 2e, f). The shoot multiplication factor was constant and limited to 2–3 during subsequent monthly culture cycles. Secondary shoots always formed at the base of the initial explant (Fig. 2e, f). The addition of 2,4-D to the CSM medium did affect the reaction pattern of cultured banana shoot tips. Ten percent of explants blackened and died within 3 months following inoculation. The remaining explants grew into a single shoot, or showed minimal shoot multiplication with the formation of ‘globules’ at their basal part. Each of these in vitro responses appeared at a similar frequency among the surviving explants. The so-called ‘globules’ are yellowish, round or kidney shaped three-layered structures that are not regenerative as such, but are sometimes closely associated with the formation of somatic embryos in banana (Strosse et al. 2006).
The inoculation of small explants (0.1 cm shoot tips) gave similar in vitro responses in maize as well as banana. However, the ensuing shoots were smaller and the response was delayed with approximately two months.
Large-scale study on the development of banana shoot tips subjected to various plant growth regulator combinations
The effect of type and concentration of cytokinins
Three months after treatment of 0.1 cm sized ‘Williams’ shoot tips with different cytokinins at various concentrations, the following developmental patterns could be distinguished: (i) lethal blackening (Fig. 3a) or lack of response (explants with pale white colour, not illustrated) both resulting in explant death, (ii) outgrowth of single shoots (Fig. 3b) and (iii) multiplication of initial explants resulting in a cluster of predominantly equally sized shoots (more than 3 mm in length Fig. 3c) or clusters of small shoots (less than 3 mm in length) spirally arranged at the base of the original shoot tip (Fig. 3d, e).

Different developmental patterns distinguished three months after inoculation of banana shoot tips on basal corn shoot multiplication (CSM) medium supplemented with (a) 100 μM 2iP, explant death, bar = 300 μm; (b) 1 μM BAP, outgrowth of single shoot, bar = 5 mm; (c) 10 μM kinetin, cluster of equally sized shoots more than 3 mm in length, bar = 5 mm; (d and e) cluster of tiny (less than 3 mm in length) shoots formed at the base of the inoculated shoot tip; (d) 100 μM BAP, bar = 1 mm, (e) 10 μM TDZ, bar = 1.6 mm; (f) 1 μM TDZ and 0.5 μM 2,4-D, cluster of globules developing at the base of the inoculated shoot tip, bar = 1.6 mm
When shoot tips were cultured on basal CSM medium devoid of plant growth regulators all explants grew into elongated single shoots (Fig. 4a, b). Except for TDZ, all three reaction patterns were observed for all cytokinins. However, the frequency of each reaction pattern depended on the applied cytokinin concentration. The highest concentration (100 μM) of cytokinins was lethal for at least part of the treated explants. The average number of shoots per initial explant was lower than one when treated with 100 μM 2iP as a consequence of explant death in the majority of inoculated shoot tips. The toxicity of tested cytokinins could be rated as follows (from high to low toxicity): 2iP (92%) > kinetin (50%) > zeatin (33%) > TDZ (17%) > BAP (8%). For multiplication, TDZ showed the strongest and kinetin the weakest activity. The weaker activity of kinetin, 2iP and zeatin (compared to BAP and TDZ) towards induction of multiple shoots was reflected in the lower average number of shoots per initial and surviving explant. In general, the average number of shoots obtained was less than five. Ten micromolar was the optimal concentration to combine multiple shoot formation with low toxicity when kinetin, 2iP or zeatin was used. Except for TDZ, a cytokinin concentration of 1 μM resulted predominantly in outgrowth of single shoots. The highest number of explants developing into multiple shoots was observed with BAP (up to 92%) and TDZ (up to 100%). In contrast to BAP being the most effective at its highest concentration (100 μM), TDZ should be applied at lower concentrations (1–10 μM).

(a) In vitro response and (b) average number of shoots formed per initial (white bars) and per surviving (white + grey bars) explants three months after inoculation of 0.1 cm sized shoot tips on basal corn shoot multiplication (CSM) medium without plant growth regulators (no PGR) or supplemented with BAP, kinetin (kine), 2iP, zeatin (zea) and TDZ at a concentration of 1, 10 and 100 μM, respectively. Standard error bars are presented for average number of shoots per surviving explant. Data are derived from two independent experiments with six shoot tips cultured per treatment
The average number of shoots induced by 100 μM BAP (4.9 per initial and 5.4 per surviving explant, respectively) was lower than those treated with 1 μM TDZ (7.8 per initial as well as surviving explant). Clusters of small shoots (less than 3 mm in length) formed at the base of the original explant (Fig. 3d, e) were almost exclusively observed after treatment with either 100 μM BAP or 10 μM TDZ. Compared to other cytokinins tested and especially when applied at lower concentrations (1–10 μM), TDZ was thus the most effective in stimulating multiple shoot formation.
The effect of explant size and cytokinins in combination with the auxin 2,4-D
As expected, small shoot tips that were inoculated on basal CSM medium devoid of plant growth regulators grew into single shoots. Bigger shoot tips resulted on average in 1.5 shoots (Table 1). Bigger shoot tips (0.5 cm in length) cultured on auxin-free but cytokinin (1–50 μM) containing medium followed the same trend as the previous experiment (data not shown). A positive correlation between cytokinin concentration and average number of shoots formed per explant was generally obtained. The combination of BAP with 0.1 μM 2,4-D resulted in a higher proliferation rate compared with other cytokinins. At 2,4-D concentrations of 0.5 μM and higher, shoots were rarely observed. While lethal blackening was only sporadically detected when bigger shoot tips were cultured on cytokinin medium in absence or with 0.1 μM 2,4-D, concentrations of 2,4-D equal or higher than 0.5 μM resulted in explant death for one-third to half of the inoculated shoot tips. In explants that were not killed by 2,4-D, 0.5 μM of this auxin triggered the formation of globules (Fig. 3f), with almost no shoots generated (Table 1). Higher 2,4-D concentrations (1 μM and 5 μM) were still associated with the formation of globules but completely suppressed shoot development.
In contrast to previous results, treatment of 0.1 cm sized shoot tips with cytokinins at 10 μM did not stimulate multiplication. Only the combination of BAP (10 μM) with 0.1 μM 2,4-D resulted in an average of nearly four shoots per surviving 0.1 cm shoot tip (Table 1). In the presence of 0.1 μM 2,4-D, the influence of shoot tip size on average number of shoots formed was highly variable. According to the cytokinin treatment applied, 1.9–7 times more shoots were formed by 0.5 cm compared to 0.1 cm shoot tips (Table 1). However, when bigger instead of smaller shoot tips were grown on cytokinin medium without 2,4-D, the average number of shoots obtained in three months old cultures was overall two to three times higher (and in case of TDZ even eight times). Nevertheless, an ‘explosion’ of shoots (as noted for maize shoot tips) was never triggered by any of the applied plant growth regulator combinations in banana.
The effect of type and concentration of auxins in combination with the cytokinin TDZ
The tested auxins can be grouped into two classes according to the developmental patterns triggered. IAA, NAA or IBA in combination with TDZ resulted at each of the tested concentrations in outgrowth and multiplication of banana shoot tips. In contrast, the in vitro response triggered by Pic and 2,4-D was highly concentration dependent. Like in the previous experiment (exploring the effect of different cytokinins in combination with 2,4-D as external auxin source), outgrowth and multiplication were the developmental patterns associated with 2,4-D concentrations lower than 0.5 μM. 2,4-D concentrations higher than 0.5 μM mainly resulted in explant death and formation of globules. The effect of Pic was similar to but weaker compared to 2,4-D since formation of globules was only encountered at 5 μM Pic in the absence of TDZ. Auxins could be placed in the same two categories when considering lethal blackening, only occurring at higher concentrations of Pic and 2,4-D.
Based on the average number of shoots formed, a similar division of the tested auxins into the two categories could be made. Culture of shoot tips on TDZ-medium supplemented with IAA, NAA or IBA resulted 3 months after initiation in values of average number of shoots per explant that hardly varied with the concentration at which plant growth regulators were supplied. Average numbers of shoots per shoot tip noted for respectively, IAA, NAA and IBA ranged from 25 to 45, 15 to 35 and 25 to 40, respectively, (Fig. 5). Raising the concentration of IAA, NAA or IBA in the culture medium caused only a slight decrease in number of induced shoots. The limited influence of changing TDZ concentration confirmed the previously encountered strong action of TDZ already evoked at 1 μM. In accordance with previous observations, it should be noted that length of shoots was often drastically reduced (i.e., from 1 cm sized shoots down to shoots smaller than 0.3 cm) at higher TDZ concentrations.

Average number of shoots per initial (white bars) and per surviving explant (white + grey bars) three months after inoculation of 0.5 cm sized shoot tips on basal corn shoot multiplication (CSM) medium supplemented with IAA, NAA, IBA, Picloram (Pic) and 2,4-D at a concentration of 0, 0.1, 0.5, 1 and 5 μM in combination with TDZ at 0, 1, 5 and 10 μM. Standard error bars are presented for average number of shoots per surviving explant. Data are derived from two independent experiments with six shoot tips inoculated per treatment
In contrast to IAA, NAA and IBA, the concentration effect of the auxins Pic and 2,4-D was highly reflected in the average number of shoots calculated for different shoot tip treatments (Fig. 5). While outgrowth of shoots and multiplication was drastically suppressed with both Pic and 2,4-D, the stronger action of 2,4-D could be deduced from the lower amount or total absence of shoots especially when 2,4-D was supplied at the higher concentrations tested (1 and 5 μM). Although, some shoots formed after Pic treatment, these shoots were curled and watery, two morphological aspects that were never observed with IAA, NAA or IBA.
Discussion
Using shoot tips as initial explants, Zhong et al. (1992) succeeded in the development of a highly efficient regeneration system for maize resulting in mixed (axillary as well as adventitious) multiple shoot cultures. These results were successfully reproduced in the present study. This protocol was also reported to be successful in other cereals such as oat (Zhang et al. 1996), sorghum (Zhong et al. 1998), barley (Zhang et al. 1999), millet (Devi et al. 2000) and wheat (Ahmad et al. 2002). First, we compared the in vitro response of banana with maize, which served as a control. Although strikingly similar in morphological aspects, the in vitro behaviour of maize and banana shoot tips was distinctly different. Besides a remarkably lower and more stable multiplication rate during subsequent culture cycles of banana shoot tips compared to maize, additional differences were blackening of induced explants, absence of elongation of stem tissue and the formation of globules in banana.
Growing banana shoot tips on a broad range of cytokinin-enriched culture media in the absence of an external auxin source indicated that the five cytokinins tested (BAP, kine, 2iP, zea, and TDZ) differed with respect to their strength, but not to their morphogenic effects. Explant death, outgrowth of shoots and multiplication of initial explants were the three types of in vitro responses encountered. While kine, 2iP and zea can be considered as weaker cytokinins, BAP and TDZ stimulated shoot multiplication at a higher rate. Exploring the effect of the same five cytokinins on in vitro seed germination and early seedling morphogenesis in lotus, Nikolic et al. (2006) also ascribed the strongest effect on shoot multiplication to TDZ, followed by BAP, zea, 2iP and kine. In banana shoot tips, a clear concentration effect was observed between the stronger cytokinins. Without a noteworthy negative side effect, BAP exerted its activity especially at higher concentrations (50–100 μM) while TDZ was already active at its lowest concentration (1 μM) but detrimental at concentrations higher than 10 μM. The higher activity towards shoot production of the urea-type cytokinin TDZ compared to adenine-type (BAP and kine) and isoprenoid-type (zea and 2iP) cytokinins, especially at lower concentrations has been reported before (Mok et al. 1982; Huetteman and Preece 1993; Sunagawa et al. 2007). Since we were not able to obtain somatic embryos on any of the banana shoot tips grown on different TDZ containing media, we can not confirm the property of TDZ to substitute both for auxins and cytokinins, as reported for many dicot species (Visser et al. 1992; Murthy et al. 1998).
Skoog and Miller (1957) were the first to highlight the importance of the cytokinin: auxin ratio in triggering an in vitro response in cultured explants. In our comparative study with maize and banana shoot tips, the plant growth regulator combination 10 μM BAP/2.5 μM 2,4-D did not stimulate multiple shoot formation in banana as was the case in maize. Therefore, we tested five cytokinins (1–50 μM) in combination with various 2,4-D concentrations (ranging from 0.1–5 μM). Furthermore, the five auxins most frequently used in tissue culture (IAA, NAA, IBA, 2,4-D and Pic, each supplied at concentrations ranging from 0.1 to 5 μM) were tested in combination with TDZ (1–10 μM). Auxins could be grouped in two classes according to their morphogenic effect when supplied in combination with cytokinin(s): IAA, NAA and IBA versus Pic and 2,4-D. In combination with IAA, NAA or IBA, the auxin effect was overruled by the action of the cytokinin TDZ and identical developmental patterns were observed as in absence of an external auxin source. The replacement of IAA, NAA or IBA by Pic or 2,4-D resulted besides explant death, outgrowth of single shoots and shoot multiplication in the formation of globules. In contrast to other auxins tested, the action of Pic and 2,4-D was highly dependent on the concentration used. The weaker action of Pic compared to 2,4-D could be deduced from the higher concentration at which globules developed and the more gradual compared to a drastic reduction of the average number of shoots formed with increasing auxin concentration. As reviewed by George (1993), IAA, NAA and IBA are mainly applied in plant propagation systems for their stimulating effect on shoot morphogenesis, while Pic and 2,4-D are especially associated with callus formation and induction of embryogenesis. Although shoot multiplication was indeed the main in vitro response triggered in banana shoot tips cultured on cytokinin enriched medium supplemented with IAA, NAA or IBA, the stimulating effect of auxins on shoot multiplication (Hu et al. 2006; Landi and Mezzetti 2006) was not confirmed for banana. In banana, the development of embryogenic callus on meristematic tissue is preceded by the appearance of globules at the explant surface (Strosse et al. 2006). Despite the formation of globules on banana shoot tips cultured in the presence of Pic and especially 2,4-D, the formation of somatic embryos was not observed at any of the plant growth regulator combinations tested.
The calculated average number of shoots per initial explant was not only dependent on the plant regulator treatment to which shoot tips were subjected, but also highly determined by the initial explant size. In general, 0.5 cm sized explants (consisting of the apical dome fully covered by three to five leaf primordia and a few millimeters of corm tissue beneath the apical dome) produced two to three times more shoots in three months old cultures than 0.1 cm sized shoot tips (consisting of the apical dome only partly covered by one leaf primordium).
Rather than the determination of the exact number of shoots formed per initial explant subjected to each of the tested plant growth regulator combinations, this study provided knowledge on the origin of secondary shoots formed in banana multiple shoot cultures. Reviewing the literature on morphogenesis in tissue culture and on shoot meristem formation during vegetative development (George 1993; Thorpe 1994; Kerstetter and Hake 1997), we consider in this study, shoots as truly adventitious when they occur at random and thus at an unexpected location, for they are formed by totipotent cells or tissues that have regained meristematic activity. In contrast, axillary shoots arise at specific locations since they are in one way or another associated with leaves or leaf axils. They are originally formed by the shoot apical meristem, growing or proliferating from cells that are still in an immature and/or meristematic state and retained their plasticity to develop into vegetative buds. In order not to generate more confusion by introducing new terms for the origin of banana shoots and still in accordance with our above definition, also the leaf-opposed buds as described by Fisher (1978) are considered by us as axillary in origin.
Whether highly efficient in vitro shoot multiplication is induced in monocots or dicots (Konan et al. 1997; Srivatanakul et al. 2000; Sticklen and Oraby 2005), all systems seem to have in common an initial period of proliferation of axillary meristems characterized by a relatively low multiplication rate of shoots. An ‘explosion’ of adventitious shoots is preceded by the enlargement of the present meristematic domes. However, some genotypes or even species are not able to surmount the initial period of rather slow multiplication and seem to be recalcitrant for adventitious shoot formation under the in vitro conditions applied (Gomes and Canhoto 2003; Monteuuis 2004). Our results indicate that also Musa is extremely recalcitrant towards adventitious shoot formation. None of the 173 different plant growth regulator combinations tested, triggered de novo shoot formation. This implies that the mode of action of all cytokinins solely consisted of reducing the apical dominance of the primary shoot resulting in the outgrowth or proliferation of meristematic tissue already present in the initial banana shoot tip. A first indication supporting this hypothesis was the relatively low and stable multiplication rate noted during subsequent culture cycles of banana shoot tips. Secondly, the calculated average number of shoots per initial explant was generally two to three times higher when the bigger (0.5 cm in length) shoot tips were inoculated instead of the smaller (0.1 cm in length) explants. We already observed that removal of corm tissue involved simultaneous removal of meristematic tissue (i.e. ‘axillary’ shoot- and/or root initials) other than the shoot apical meristem. This is supported by the findings of Okole and Schulz (1996) and Li et al. (2006) studying shoot morphogenesis in banana micro-sections from which the apical meristem was removed. These authors reported that more shoots were formed from explants cut out from regions close to the apical meristem compared to more distal sections. Thirdly, dedifferentiation of tissue triggered by higher concentrations of Pic and 2,4-D was not followed by de novo shoot formation in banana shoot tips. Furthermore, secondary shoots were always spirally arranged at the base of banana explants. While adventitious shoots originate most often at random locations (Zhong et al. 1992), the spiral arrangement of secondary shoots according to a fixed phyllotaxic pattern is a fourth indication towards the outgrowth of bud initials already present in the cultured explant.
The determination of the origin of multiple shoots in banana (axillary versus adventitious—associated with predetermined outgrowth of bud initials versus de novo formation of shoots from dedifferentiated tissue, respectively) is important for further applications. When commercial propagation or safe storage is pursued, ‘axillary’ propagation (even when subjected to high concentrations of cytokinin) is advantageous to reduce the risk of somaclonal variation (George 1993). In contrast, triggering adventitious shoot formation in banana could, as also seen in maize (Zhang et al. 1999), significantly improve transformation efficiency as no embryogenic cell suspension would be needed. This is highly desired as high quality banana embryogenic cell suspensions remain hard to establish and maintain (Schoofs et al. 1999). The fact that banana MMCs are less homogeneously consisting of meristematic tissue compared to maize could be explained by the different origin of multiple shoots in both plants. In contrast to cereal crops (Sticklen and Oraby 2005), the shoot apical meristem could not be identified as a sustainable explant for fast and large-scale plant regeneration in banana.
Abbreviations
- 2,4-D:
-
2,4-Dichlorophenoxy acetic acid
- 2iP:
-
2-Isopentenyladenine
- BAP:
-
Benzylaminopurine
- CSM:
-
Corn shoot multiplication
- IAA:
-
Indole acetic acid
- IBA:
-
Indole butyric acid
- ITC:
-
International transit center
- Kine:
-
Kinetin
- MMC:
-
Multiple meristem cultures
- MS:
-
Murashige and Skoog
- MSac :
-
MS medium supplemented with 0.5 g/l active charcoal
- NAA:
-
Naphthalene acetic acid
- Pic:
-
Picloram
- TDZ:
-
Thidiazuron
- Zea:
-
Zeatin
References
Ahmad A, Zhong H, Wang WL, Sticklen MB (2002) Shoot apical meristem: In vitro regeneration and morphogenesis in wheat (Triticum aestivum L.). In Vitro Cell Dev Biol Plant 38:163–167. doi:10.1079/IVP2001267
Banerjee N, Vuylsteke D, De Langhe E (1986) Meristem tip culture of Musa: histomorphological studies of shoot bud proliferation. In: Withers LA, Anderson PG (eds) Plant tissue culture and its agricultural applications. Butterworth Scientific Ltd, London, UK, pp 139–147
Barker WG, Steward FC (1962) Growth and development of the banana plantI: the growing regions of the vegetative shoot. Ann Bot (Lond) 26:389–411
Benson EE (2000) In vitro plant recalcitrance: an introduction. In Vitro Cell Dev Biol Plant 36:141–148. doi:10.1007/s11627-000-0029-z
Cronauer SS, Krikorian AD (1987) Adventitious shoot production from calloid cultures of banana. Plant Cell Rep 6:443–445
Devi P, Zhong H, Sticklen MB (2000) In vitro morphogenesis of pearl millet [Pennisetum glaucum (L.) R.Br.]: efficient production of multiple shoots and inflorescences from shoot apices. Plant Cell Rep 19:546–550. doi:10.1007/s002990050771
Dhed’a D, Dumortier F, Panis B, Vuylsteke D, De Langhe E (1991) Plant regeneration in cell suspension cultures of the cooking banana cv. ‘Bluggoe’ (Musa spp. ABB group). Fruits 46:125–135
Fisher JB (1978) Leaf-opposed buds in Musa: their development and a comparison with allied monocotyledons. Am J Bot 65:784–791. doi:10.2307/2442154
George EF (1993) Plant propagation by tissue culture-Part 1: the technology. Exegetics Ltd, Edington, UK, p 574
Gomes F, Canhoto JM (2003) Micropropagation of Eucalyptus nitens Maiden (Shining Gum). In Vitro Cell Dev Biol Plant 39:316–321. doi:10.1079/IVP2002376
Hu XR, Yang AF, Zhang KW, Wang J, Zhang JR (2006) Optimization of in vitro multiple shoot clump induction and plantlet regeneration of Kentucky bluegrass (Poa pratensis). Plant Cell Tissue Organ Cult 84:89–98. doi:10.1007/s11240-005-9009-7
Huetteman CA, Preece JE (1993) Thidiazuron: a potent cytokinin for woody plant tissue culture. Plant Cell Tissue Organ Cult 33:105–119. doi:10.1007/BF01983223
Kerstetter RA, Hake S (1997) Shoot meristem formation in vegetative development. Plant Cell 9:1001–1010. doi:10.1105/tpc.9.7.1001
Konan NK, Schöpke C, Cárcamo R, Beachy RN, Fauquet C (1997) An efficient mass propagation system for cassava (Manihot esculenta Crantz) based on nodal explants and axillary bud-derived meristems. Plant Cell Rep 16:444–449
Landi L, Mezzetti B (2006) TDZ, auxin and genotype effects on leaf organogenesis in Fragaria. Plant Cell Rep 25:281–288. doi:10.1007/s00299-005-0066-5
Li J, Huang X-L, Wei Y-R, Huang X, Li Z, Li X-J (2006) Histological analysis of direct organogenesis from micro-cross-sections of cultures of the banana. Aust J Bot 54:595–599. doi:10.1071/BT05013
Mok MC, Mok DWS, Armstrong DJ, Shudo K, Isogai Y, Okamoto T (1982) Cytokinin activity of N-phenyl-N′-1, 2, 3-thiadiazol-5-ylurea (thidiazuron). Phytochem 21:1509–1511
Monteuuis O (2004) In vitro micropropagation and rooting of Acacia mangium microshoots from juvenile and mature origins. In Vitro Cell Dev Biol Plant 40:102–107. doi:10.1079/IVP2003493
Murashige T, Skoog F (1962) A revised medium for rapid growth and bioassays with tobacco tissue cultures. Physiol Plant 15:473–497. doi:10.1111/j.1399-3054.1962.tb08052.x
Murthy BNS, Murch SJ, Saxena PK (1998) Thidiazuron: a potent regulator of in vitro plant morphogenesis. In Vitro Cell Dev Biol Plant 34:267–375. doi:10.1007/BF02822732
Nikolic R, Mitic N, Miletic R, Neskovic M (2006) Effects of cytokinins on in vitro seed germination and early seedling morphogenesis in Lotus corniculatus L. J Plant Growth Regul 25:187–194. doi:10.1007/s00344-005-0129-4
Okole BN, Schulz FA (1996) Micro-cross sections of banana and plantains (Musa spp.): morphogenesis and regeneration of callus and shoot buds. Plant Sci 116:185–195. doi:10.1016/0168-9452(96)04381-6
Panis B, Piette B, Swennen R (2005) Droplet vitrification of apical meristems: a cryopreservation protocol applicable to all Musaceae. Plant Sci 168:45–55. doi:10.1016/j.plantsci.2004.07.022
Sági L, Panis B, Remy S, Schoofs H, De Smet K, Swennen R et al (1995) Genetic transformation of banana and plantain (Musa spp.) via particle bombardment. Biotechnology 13:481–485
Schoofs H, Panis B, Strosse H, Mayo Mosqueda A, Lopez Torres J, Roux N et al (1999) Bottlenecks in the generation and maintenance of morphogenic banana cell suspensions and plant regeneration via somatic embryogenesis therefrom. InfoMusa 8:3–7
Skoog F, Miller CO (1957) Chemical regulation of growth and organ formation in plant tissue cultured in vitro. Symp Soc Exp Biol 11:118–131
Srivatanakul M, Park SH, Sanders JR, Salas MG, Smith RH (2000) Multiple shoot regeneration of kenaf (Hibiscus cannabinus L.) from a shoot apex culture system. Plant Cell Rep 19:1165–1170
Sticklen MB, Oraby HF (2005) Shoot apical meristem: a sustainable explant for genetic transformation of cereal crops. In Vitro Cell Dev Biol Plant 41:187–200. doi:10.1079/IVP2004616
Strosse H, Schoofs H, Panis B, Andre E, Reyniers K, Swennen R (2006) Development of embryogenic cell suspensions from shoot meristematic tissue in bananas and plantains (Musa spp.). Plant Sci 170:104–112. doi:10.1016/j.plantsci.2005.08.007
Sunagawa H, Agarie S, Umemoto M, Makishi Y, Nose A (2007) Effect of urea-type cytokinins on the adventitious shoots regeneration from cotyledonary node explant in the common ice plant, Mesembroanthemum crystallinum. Plant Prod Sci 10:47–56. doi:10.1626/pps.10.47
Thorpe TA (1994) Morphogenesis and regeneration. In: Vasil IK, Thorpe T (eds) Plant Cell and Tissue Culture. Kluwer Academic Publishers, Dordrecht, The Netherlands, pp 17–36
van den Houwe I, De Smet K, Tezenas du Montcel H, Swennen R (1995) Variability in storage potential of banana shoot cultures under medium term storage conditions. Plant Cell Tissue Organ Cult 42:269–274. doi:10.1007/BF00029998
Vasil IK (1990) The realities and challenges of plant biotechnology. Biotechnology 6:296–300. doi:10.1038/nbt0490-296
Visser C, Qureshi JA, Gill R, Saxena PK (1992) Morphoregulatory role of thidiazuron. Plant Physiol 99:1704–1707
Vuylsteke D, De Langhe E (1985) Feasibility of in vitro propagation of bananas and plantains. Trop Agric 62:323–328
Zaffari GR, Kerbauy GB, Kraus JE, Romano EC (2000) Hormonal and histological studies related to in vitro banana bud formation. Plant Cell Tissue Organ Cult 63:187–192. doi:10.1023/A:1010611318932
Zhang S, Cho MJ, Koprek T, Yun R, Bregitzer P, Lemaux PG (1999) Genetic transformation of commercial cultivars of oat (Avena sativa L.) and barley (Hordeum vulgare L.) using in vitro shoot meristematic cultures derived from germinated seedlings. Plant Cell Rep 18:959–966. doi:10.1007/s002990050691
Zhang S, Zhong H, Sticklen MB (1996) Production of multiple shoots from shoot apical meristems of oat (Avena sativa L.). J Plant Physiol 148:667–671
Zhong H, Srinivasan C, Sticklen MB (1992) In vitro morphogenesis of corn (Zea mays L.) I: differentiation of multiple shoot clumps and somatic embryos from shoot tips. Planta 187:483–489
Zhong H, Wang WL, Sticklen MB (1998) In vitro morphogenesis of Sorghum bicolour (L.) Moench: efficient plant regeneration from shoot apices. J Plant Physiol 153:719–726
Acknowledgements
This work was possible due to the financial support of DGDC (Directorate-General for Development Cooperation), Belgium through a grant to IITA (International Institute of Tropical Agriculture) and INIBAP (International Network for the Improvement of Banana and Plantain, currently Bioversity International). We thank Dr. Sándor Mórocz at the Cereal Research Non-Profit Company (Szeged, Hungary) for supplying maize seeds. Special thanks also to all colleagues of the Laboratory of Tropical Crop Improvement who contributed through many discussions to an improved knowledge on the origin of multiple shoot cultures in banana.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Strosse, H., Andre, E., Sági, L. et al. Adventitious shoot formation is not inherent to micropropagation of banana as it is in maize. Plant Cell Tiss Organ Cult 95, 321–332 (2008). https://doi.org/10.1007/s11240-008-9446-1
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11240-008-9446-1