
Abstract
In this paper, studies on photoautotrophic micropropagation (PM) and inoculant cultivation of arbuscular mycorrhizal fungi (AMF) were reviewed. The existing issues, which include the lack of a bio-method for improving the environmental stress resistance of plantlets in vitro or ex vitro and the mass production of AMF inoculants in commercialization for PM and inoculants cultivation, were discussed. Based on current knowledge and our analysis, we suggested that the integration of PM system and mycorrhization in vitro (here called mycorrhization-PM cultivation system) is highly feasible. Furthermore, combining mycorrhization with PM system in vitro was shown to be an efficient method for alleviating the injuries of plantlets in vitro and ex vitro caused by environmental stresses and for mass producing AMF inoculants through applying different cultivation strategies. Cultivation strategies for the mass production of mycorrhizal plants and AMF inoculants using mycorrhization-PM cultivation system were discussed.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
With the growing world population and the deterioration of the environment, more and more high-quality seedlings will be needed worldwide for agriculture, forestry, horticulture, and eco-restoration. It was estimated that about 20–40 billion plants are required per year for re-afforestation alone (Kozai and Kubota 2001). Micropropagation has established its role as a way to produce large quantities of uniform high-value horticultural crops such as ornaments, fruits, vegetables, plantation crops and spices (Vestberg and Estaún 1994; Vestberg et al. 2002). It has been reported that about 50% of flori-horticultural plants were produced by the micropropagation technique (Sahay and Varma 2000). Conventionally, micropropagated plantlets in vitro are photoheterotrophic or photomixotrophic since sugar is added to the growth media as a source of carbon (Murashige and Skoog 1962). Explants have to be cultured in small containers (triangular flask, test tube, etc.) with little gas exchange to avoid contamination, which results in inner stress microenvironments with high relative humidity, low photosynthetic photon flux density (PPFD), harmful gases accumulation (e.g., ethylene) and depletion of CO2 concentration. Thus, some physiological disorders occur in the plantlets, and the photosynthetic organs are not able to function properly. In addition, plantlets grow heterotrophically and develop with high water content, which implies a great risk of desiccation at acclimatization (Kubota and Kozai 1992) for abnormal anatomical features (e.g., somatic function). In some commercial micropropagation systems of flowers and other ornamental plants, losses during the acclimatization stage were up to 10–40% or higher (Varma and Schūepp 1995).
In the late 1980s, Kozai and co-workers created a novel micropropagation method called sugar-free micropropagation or photoautotrophic micropropagation (PM) (Kozai and Iwanami 1988; Kozai et al. 1988; Fujiwara et al. 1988). PM is characterized by a controllable culture microenvironment in which plantlets are grown photoautotrophically with enriched CO2 concentration, the optimal amount of air exchange and PPFD. Furthermore, plantlets could be cultured in large containers on porous and solid supporting materials (perlite, vermiculite, etc.) without sugar. In a PM system, the physiological status of plantlets (e.g., photosynthetic capacity) was substantially strengthened. Many advantages of PM have been well studied and documented in previous studies with respect to its biological and engineering aspects (Kozai and Kubota 2005), including (1) promotion of growth and photosynthesis, (2) high survival percentage/smooth transition to ex vitro environment, (3) elimination of morphological and physiological disorders, (4) infrequent loss of plantlets due to contamination, (5) flexibility in the design of the vessels and control systems, and (6) simplification of the micropropagation procedure (the acclimatization stage necessary in conventional micropropagation can often be eliminated or shortened). Because of these advantages, PM is believed to be a commercial alternative to conventional propagation for high quality transplants (Kozai 1991b; Xiao et al. 2000; Kozai and Kubota 2001; Xiao and Kozai 2004; Kozai and Kubota 2005; Kozai et al. 2006).
Arbuscular mycorrhizal fungi (AMF) are multi-functional microorganisms with high biodiversity that can form symbioses with more than 90% of plant species by colonizing root tissues and soil (Newsham et al. 1995; Smith and Read 1997). They promote plant fitness by improving physiological parameters, the uptake of water and nutrients, disease resistance, etc. In ecosystems, AMF play an important role in determining plant biodiversity, ecosystem variability and productivity (van der Heijden et al. 1998a, b), and soil ecological stability. Lovato et al. (1996) regarded the various functions performed by AMF as biofertilizers, bioprotectors, and bioregulators. AMF exist in almost all kinds of soil ecosystems. Therefore, it is believed that there are efficient isolates at specific geographic sites for local mycorrhizal plant species with positive inoculation efficiency. AMF are thus regarded as the most promising beneficial microorganisms that can be produced and applied commercially in the practice of plant production. However, few case studies with a large input quantity of inoculants have been conducted in the field, with the exception of some specific cases, e.g., mine restoration (Saito and Marumoto 2002). Taking into consideration the economy and operability, most previous studies focused on developing methods for AMF application in transplant systems due to the advantages of low inoculant dosage and early symbiosis formation ahead of field growth, features that are helpful for the post-transplantation establishment of plantlets. Mycorrhization treatment of plantlets is currently considered an effective method for enhancing post-transplantation establishment.
Plantlets are first grown in vitro under aseptic conditions devoid of AMF and other rhizosphere microorganisms; manufactured substrates also often lack AMF at the acclimatization stage. Therefore, there is almost no chance for arbuscular mycorrhizal formation prior to transplantation without artificial introduction. Thus, it has been suggested that plantlets could be better protected against the biotic and abiotic stresses that occur in greenhouses or in the field by associating the plantlets with AMF at early stage (Vestberg et al. 2004). Furthermore, by introducing AMF, it would be possible to reduce fertilizer and pesticide use and to grow the plants in a more sustainable way (Cordier et al. 2000). However, commercial application of AMF in transplant systems has not been used extensively due to an inadequate supply of AMF inoculants resulting from a lack of efficient cultivation methods. In addition, the lagging AMF introduction time at the transplantation stage in conventional micropropagation often lead to a reduced symbiotic efficiency caused by C competition between the symbionts.
Many researchers have suggested that mycorrhization is beneficial for the establishment and health of propagated plantlets post-transplantation (Nowak 1998; Sahay and Varma 2000; Rai 2001; Nowak and Shulaev 2003). In addition, numerous data have indicated that the PM system is well developed by in vitro environmental control techniques (Aitken-Christie et al. 1995; Kozai et al. 1997; Zobayed et al. 2001; Xiao and Kozai 2004). However, no study has been published dealing with the integration of PM and mycorrhization for micropropagation in vitro. Compared with AMF inoculation at transplantation, there are more benefits for micropropagated plantlets when AMF is introduced in vitro into PM systems for overcoming stresses in vitro and ex vitro. For example, C competition between the symbionts will occur at the rooting stage, not the transplantation stage. Therefore, the increased photosynthetic capacity of plantlets in PM systems will compensate for C deficiency caused by C competition between the host plant and AMF. In addition, the integration of PM and mycorrhization in vitro will avoid coincidental C competition and transplantation shock. In short, mycorrhization in vitro does markedly enhance the growth and development of plantlets more than conventional inoculation with the aid of appropriate environmental controls. Theoretically, the combination of PM systems and mycorrhization (referred to as a mycorrhization-PM cultivation system) provides a pathway both for mass producing AMF inoculants with the aid of proper environmental controls and for promoting survival and health of the mycorrhizal plantlets by staving off injuries caused by biotic and abiotic stresses. This paper mainly summarizes how research has progressed and the existing problems in the commercial application in PM systems and AMF inoculants cultivation; it also highlights the feasibility and cultivation strategies of the mycorrhization-PM cultivation system, with two objectives in mind: AMF inoculants cultivation and mycorrhizal transplants production on a large scale.
PM systems: a commercialized method for transplant production
Environmental control is essential for promoting the photosynthetic growth and development of in vitro plantlets (Kozai et al. 1997). The establishment and optimization of the culture microenvironment in vitro is needed for commercial application of PM systems. Researchers in several countries, particularly Japan, have been working on PM systems. Today, PM systems have shifted from experimental study to commercial application after successful developments in culture systems and control devices. Currently, three kinds of commercialized PM systems (integrated chamber, large culture vessel and cleanliness room) and associated control devices have been developed. Generally, inner irradiance, relative humidity and CO2 concentration are controlled in all the above-mentioned PM systems, either automatically or manually.
The earliest PM system was the large culture vessel, which used forced ventilation to import pure CO2 or air-enriched CO2. Culture vessels range in volume from about several liters to 180 l (Kozai 1991a; Xiao and Kozai 2004; Liu et al. 2006; Guan 2007). Two practical models of culture vessels, with holes sealed with filters (Kozai 1991a) or with a top-seal film (Tanaka et al. 1988a, b), were developed as experimental tools used for in vitro culture in large culture vessels. Previous data had shown that these culture vessels enhanced the in vitro growth of many plant species, including coffee (Nguyen et al. 1999), anthurium, syngonium, agapanthus (Tanaka et al. 1996), cymbidium (Tanaka et al. 1999), spathiphyllum (Teixeira da Silva et al. 2006) and eucalyptus (Nhut et al. 2002) under photoautrophic conditions. A cleanliness room is another kind of PM system, with a high cleanliness classification in addition to general environmental controls. It uses an enlarged culture vessel that allows more plantlets to be grown in open or air-permeable small vessels with the assistance of physical and chemical pasteurization. Cleanliness technology was introduced into the PM system to guarantee a reduced incidence of bio-contamination (Liu and Yang 2006). The cleanliness control system reported by Liu and Yang (2006) performed at up to ISO7 grade cleanliness classification, according to ISO 14644-1:1999 (E), which made it possible to culture plantlets’ exposure to inner air with a low incidence of contamination caused by microbes brought inside by mixed air and operators. Before the cleanliness room technique was introduced, Kozai and Kubota (2001) developed a scale-up micropropagation system using an aseptic culture room. In this system, no one is permitted to enter the culture room to handle the trays with plants. Thus, both tray transportation and environmental control in the culture room must be automated. Compared to this culture system, the cleanliness room is a more feasible system in developing countries because of the low cost. The integrated chamber, a movable and more complicated product, is a kind of newly developed culture chamber in China that combines culture systems with environmental control devices.
Control precision of CO2 concentration in the above-mentioned PM systems reaches 50 μmol mol−1. CO2 enrichment for the integrated chamber and the large culture vessel can be achieved by releasing pure or air-enriched CO2 gas using a forced ventilation system. However, for the large culture vessels, inner CO2 enrichment can also be achieved by releasing pure or air-enriched CO2 gas into a culture room using forced ventilation. The inner CO2 concentration of the vessel is then balanced through permeable filters sealed on the vents in the vessel. In addition, relative humidity in the culture system (vessel, room and chamber) can be controlled automatically using a water-absorbent device and humidifier. In order to decrease the frequency of renewing the absorbent materials in the water-absorbent device, a membrane-cover technique was developed to keep water in the substratum (Guan 2007). The higher quality of plantlets and the economic advantages of commercial applications of PM systems over conventional methods have been demonstrated for calla lily (Zantedeschia elliottiana) and China fir (Cunninghamia lanceolata) plantlet production, using the large culture vessels (120 l) (Xiao and Kozai 2004). PM systems are currently gradually being applied commercially in China for the production of high value plants.
Inoculants mass production: a limitation of commercial application of AMF
Although it is well known that AMF have great application potential in agriculture, horticulture and eco-environmental protection, insufficient inoculants are cultured by the methods currently available. In the past, a few cultural methods, such as pot culture, hydroponic culture, and aeroponic culture, as well as in vitro culture systems, were developed as research tools with little consideration for large quantity inoculant production. No commercial method for AMF inoculant production has been developed because it is difficult to meet the strict symbiotic conditions and aseptic environment maintenance required for AMF cultivation due to their obligatory biotrophic nature.
Pot culture is the most common method for experimental study and inoculant production, but it is difficult to avoid bio-contamination and to implement efficient environmental regulations during culture. Only by modifying the physical and chemical properties of culture substrates can regulate the host-AMF relationship. Methods of hydroponic and aeroponic culture are difficult to master for common producers, and the quality of AMF products is hard to control. Some other methods, such as in vitro aseptic culture and organ culture, need complex technology with little or no commercial inoculant output. In the mid-nineties, two kinds of in vitro culture systems, tripartite (strawberry plant–mycorrhizal carrot root) (Elmeskaoui et al. 1995) and bipartite (strawberry-AMF) (Cassells et al. 1996), were developed for mycorrhization of micropropagated plantlets. For these systems, studies were performed on water uptake and stomatal conductance (Hernández-Sebastià et al. 1999, 2000), the effects of light and CO2 on the growth and photosynthesis of mycorrhizal plants (Louche-Tessandier et al. 1999), and the use of mycorrhizal vitroplants in the micropropagation industry (Mark et al. 1997). In particular, Voets et al. (2005) recently developed an autotrophic culture system for the in vitro mycorrhization of potato plantlets with shoots grown exposed to air conditions. In summary, AMF have been successfully inoculated to several crops, even in vitro (Pons et al. 1983; Cassells et al. 1996; Duffy et al. 1999; Voets et al. 2005), but these methods are not suitable for practical use for AMF inoculant mass production. A practicable method for AMF introduction in vitro is urgently needed.
Mycorrhization-PM cultivation systems for mycorrhizal plantlets and inoculants production: feasibility analysis and verification
Based on current knowledge, the key issues that need to be solved for AMF commercial production are (1) well-kept axenic condition and (2) optimal environment control. In the 1980s and 1990s, numerous experiments showed that mycorrhizal micropropagated plantlets had better growth, physiological parameters and survival rates than their non-mycorrhizal counterparts (Salamanca et al. 1992; Azcón-Aguilar et al. 1996; Azcón-Aguilar and Barea 1997; Subhan et al. 1998). More involved references were given by Rai (2001). However, most studies on mycorrhization were conducted ex vitro due to the lack of feasible in vitro culture systems. PM successfully ensures that the physiological functions of plantlet organs (e.g., leaves and roots) develop normally before transplantation. However, the biotic and abiotic stresses to which the plant is exposed after transplantation still cause some problems with plantlet survival ex vitro. In addition, water and nutrient depletion may occur during the rooting stage of PM cultivation. Transplants have difficulty overcoming post-transplantation stresses since they are devoid of natural allies such as AMF. There is great potential for introducing AMF into PM systems for the purpose of protecting transplants against in vitro and ex vitro stresses. Mycorrhization of conventional propagation is not possible in vitro for agar media with a high concentration of salt (particularly P) and sugar. Mycorrhization in vitro of micropropagated plantlets became feasible when the PM system was developed based on four aspects. First, PM systems provide suitable conditions for AMF introduction and colonization, e.g., a largely aseptic growth space, compatible host plant, and low mineral nutrients (especially P). Second, controllable environmental conditions (lighting, CO2 concentration, air current rate, temperature, etc.) make it possible to mediate interactions between AMF and host plantlets. Third, no complicated operations were added because AMF can be inoculated in combination with rooting stage operation, and rapid rooting can ensure formation of mycorrhizal roots. PM systems have been commercially applied in transplant production. Based on the above-mentioned feasibility analysis, arbuscular mycorrhizas can be well established in PM systems with plantlets at the rooting stage. The main advantages of the mycorrhization-PM cultivation system are as follows. First, AMF introduction is conducted earlier than previous methods, at the rooting stage. Thus, the critical carbon-competition between AMF and host plantlets will not occur at the transplant stage (and thus will not coincide with transplant shock). Second, new emergent roots are more suitable for AMF colonization than old ones, which means that the symbiotic relationship can be easily established.
The mycorrhization-PM cultivation system can improve the fitness of plantlets in vitro and ex vitro through two pathways. First, PM improves the growth and development of plantlets by shoot regulation, i.e., increasing photosynthetic capacity, leaf expansion and photosynthate utilization, while mycorrhizal fungi will enhance the growth and development of plantlets by modifying root morphology and strengthening their uptake function. Mycorrhization in vitro can mediate photosynthetic physiology and CO2 utilization by increasing the carbon sink strength in order to form intraradical and extraradical fungal tissue. Most importantly, symbiotic formation in vitro can take advantage of the interaction benefits between AMF and controllable environmental factors to improve the growth and development of plantlets synergistically. Guan (2007) first demonstrated mycorrhization in vitro with pre-rooting plantlets of the Chinese flowering crabapple (Malus pruniolia var ringo) in a PM system (a 180 l large vessel with a control device and forced air ventilation importing controllably pure CO2) with 1500 ± 50 μmol mol−1 CO2 concentration. The results showed that mycorrhizal symbiosis was well established, and the growth and photosynthetic parameters of the plantlets, such as stomatal resistances, stomatal conductance, chlorophyll content and CO2 fixation, and photosynthesis rate, were promoted by mycorrhizal colonization.
The mycorrhization-PM cultivation system provides a novel method to produce AMF inoculants. Our recent studies with sweet potato plantlets indicated that 40–60% of root colonization was observed, and considerable spores in substrate (vermiculite) could be obtained after 4 months cultivation in the mycorrhization-PM cultivation system with 1500 ± 50 μmol mol−1 CO2 concentration (unpublished data). The mycorrhization-PM cultivation system can be established in vitro as early as the rooting stage only because arbuscular mycorrhizas fail to establish without root organs due to their obligatory biotrophic nature. Taking the biological properties of mycorrhizal fungi into consideration, optimal environmental factors are important for the growth, development and functions of AMF. Therefore, the advantages of PM with controllable environmental conditions (lighting, temperature and substrate properties, etc.) can be collectively utilized to regulate the growth and development of AMF, which is indispensable for AMF commercial cultivation.
Mycorrhization-PM cultivation systems for mycorrhizal plantlets and inoculants production: cultivation strategies
Taking the biological properties of AMF and plantlets into consideration, there are three kinds of factors (biological, environmental and management) that directly influence the performance of mycorrhization-PM cultivation systems (Table 1). Therefore, optimal culture strategies should be developed for mycorrhizal plantlet production and AMF inoculant cultivation, respectively. Through proper culture strategies, the maximal biological benefits of integration between PM and mycorrhization will be achieved at a low cost. In the case of plantlet production, the optimal culture strategy is to facilitate the formation of mutualistic symbiosis, which will assist the fitness of plantlets in vitro and ex vitro. For AMF inoculant cultivation, the optimal culture strategy is to increase the growth and development of fungal partners for more fungal propagules. To summarize, cultivation strategies for mycorrhization-PM cultivation systems are necessarily developed to purposely regulate mycorrhizal status for more products, i.e., high-quality plantlets and inoculants, separately. It is well documented that environmental factors can determine the symbiotic status of arbuscular mycorrhizas, leading to shifts in parasitism, mutualism and commensalism (Johnson et al. 1997). The physiological status of mycorrhizal plantlets in turn affects the carbon distribution between plantlets and fungi, which directly determines the number of fungal propagules. In short, mutual symbiosis is needed to promote the quality of plantlets, while parasitic symbiosis may improve AMF inoculants production. Proper cultivation strategies are important for the two symbionts to function well in mycorrhization-PM cultivation system in order to obtain the desired products.
Among environmental conditions, lighting, CO2 concentration, temperature, and substrate conditions are the main environmental factors previously studied for AMF cultivation. The mechanism of action was to mediate the C distribution between the two partners of AMF and host plants. In addition, there are important interactions between most factors (e.g., lighting and CO2 concentration). Lighting, CO2 concentration and substrate conditions are also key factors in PM systems, and they have been studied extensively. However, no optimal environmental parameters have been determined with respect to mycorrhization-PM cultivation systems. For lighting and CO2 concentration, previous studies concentrated mainly on the effect of interactions between lighting and CO2 concentration on AMF function and development, as well as on the growth of plantlets in in vitro tripartite systems (Elmeskaoui et al. 1995; Louche-Tessandier et al. 1999; Gavito et al. 2002). For substrate conditions, pH (Clark and Zeto 1996; van Aarle et al. 2002), temperature (Addy et al. 1994, 1998; Wang et al. 2002; Gavito et al. 2003), and the interaction effects of atmospheric CO2 concentration and soil temperature on P uptake by AMF were examined (Gavito et al. 2003). In addition, it is well known that high P levels inhibit the root colonization and function of AMF (Abbott et al. 1984), and the porosity of the substratum has an impact on AMF growth and the function of extraradical hyphae, as previous data have shown that compacted soil prevents hyphae passing through to the far site to absorb nutrients (Li et al. 1997). As far as mycorrhization-PM cultivation systems are concerned, there are few data about interactions among environmental factors, plantlets and AMF; numerous investigations are needed to implement to enrich the results previously obtained under field conditions or for in vitro systems.
Concluding remarks
In practice, the production of high-quality tissue culture-derived plants on a large scale is the initial target of PM. However, as an in vitro system, the originated plants are free of any beneficial microorganisms. Introducing beneficial microorganisms before transplantation into the field has been highly recommended for alleviating stress injuries under field conditions (Nowak 1998; Rai 2001). Combined with mycorrhization, PM becomes a multi-functional technique (called the mycorrhization-PM cultivation system) pursuing two objectives: (1) the enhancement of the fitness of plantlets in vitro and ex vitro and (2) the mass production of AMF inoculants. Through feasibility analysis, it is concluded that mycorrhization-PM cultivation systems are feasible for the mass production of high quality transplants and inoculants of AMF through applying different cultivation strategies (Fig. 1). Two issues in mycorrhization-PM cultivation systems should be addressed in order to achieve those functions: the selection of suitable plant species and AMF species or isolates, and the development of optimal cultivation strategies (all relevant environmental parameters), i.e., optimization of the whole plant–substrate-microorganism system. It is believed that, when the above issues are solved, mycorrhization-PM cultivation systems will perform better in the mass production of transplants and AMF inoculants with regard to both quality and quantity.

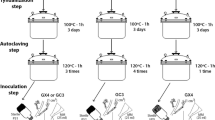
Diagrammatic representation of mycorrhization-PM cultivation system for mycorrhizal transplants and AMF inoculant mass production
Abbreviations
- AMF:
-
Arbuscular mycorrhizal fungi
- C:
-
Carbon
- CO2 :
-
Carbon dioxide
- PM:
-
Photoautotrophic micropropagation
- PPFD:
-
Photosynthetic photon flux density
References
Abbott LK, Robson AD, de Boer G (1984) The effect of phosphorus on the formation of hyphae in soil by vesicular-arbuscular mycorrhizal fungi, Glomus fasciculatum. New Phytol 97:437–446. doi:10.1111/j.1469-8137.1984.tb03609.x
Addy HD, Schaffer GF, de Boer G (1994) Survival of the external mycelium of a VAM fungus in frozen soil over winter. Mycorrhiza 5:1–5. doi:10.1007/BF00204013
Addy HD, Boswell EP, Koide RT (1998) Low temperature acclimation and freezing resistance of extraradical VA mycorrhizal hyphae. Mycol Res 102:582–586. doi:10.1017/S0953756297005376
Aitken-christie J, Kozai T, Smith MAL (1995) Automation and environmental control in plant tissue culture. Kluwer Academic Publishers, Dordrecht
Azcón-Aguilar C, Barea JM (1997) Applying mycorrhiza biotechnology to horticulture-significance and potentials. Sci Hortic (Amsterdam) 68:1–24. doi:10.1016/S0304-4238(96)00954-5
Azcón-Aguilar C, Padilla IG, Encina CL, Azcón R, Barea JM (1996) Arbuscular mycorrhizal inoculation enhances plant growth and changes root system morphology in micropropagated Annona cherimola Mill. Agronomie 16:647–652. doi:10.1051/agro:19961009
Cassells AC, Mark GL, Periappuram C (1996) Establishment of arbuscular mycorrhizal fungi in autotrophic strawberry cultures in vitro. Comparison with inoculation of microplants in vivo. Agronomie 16:625–632. doi:10.1051/agro:19961006
Clark RB, Zeto SK (1996) Mineral acquisition by mycorrhizal maize grown on acid and alkaline soil. Soil Biol Biochem 28:1495–1503. doi:10.1016/S0038-0717(96)00163-0
Cordier C, Lemoine MC, Lemanceau P, Gianinazzi-Pearson V, Gianinazzi S (2000) The beneficial rhizosphere: a necessary strategy for microplant production. Acta Hortic 530:259–268
Duffy EM, Hurley EM, Cassells AC (1999) Weaning performance of potato microplants following bacterization and mycorrhization. Potato Res 42:521–527. doi:10.1007/BF02358168
Elmeskaoui A, Damont JP, Poulin MJ, Piche Y, Desjardins Y (1995) A tripartite culture system for endomycorrhizal inoculation of micropropagated strawberry plantlets in vitro. Mycorrhiza 5:313–319. doi:10.1007/BF00207403
Fujiwara K, Kozai T, Watanbe I (1988) Development of a photoantotrophic tissue culture system for shoot and/or plantlets at rooting stage and acclimatization stages. Acta Hortic 230:153–158
Gavito M, Bruhn D, Jakobsen I (2002) P uptake by arbuscular mycorrhizal hyphae does not increase when the host plant grows under atmospheric CO2 enrichment. New Phytol 154:751–760. doi:10.1046/j.1469-8137.2002.00404.x
Gavito M, Schweiger P, Jakobsen I (2003) P uptake by arbuscular mycorrhizal fungi: effects of soil temperature and atomospheric CO2 enrichment. Glob Change Biol 9:106–116. doi:10.1046/j.1365-2486.2003.00560.x
Guan DP (2007) Influence of arbuscular mycorrhizal fungi and sugar-free culture on physiological effects of malus pruniolia var ringo plantlets in vitro. Doctoral Dissertation of Chinese Academy of Agricultural Sciences, Beijing
Hernández-Sebastiá C, Piche Y, Desjardins Y (1999) Water relations of whole strawberry plantlets in vitro inoculated with Glomus intraradices in a tripartite culture system. Plant Sci 143:81–91. doi:10.1016/S0168-9452(99)00014-X
Hernández-Sebastiá C, Samson G, Bernier PY, Piche Y, Desjardins Y (2000) Glomus intraradices causes differential changes in amino acid and starch concentrations of in vitro strawberry subjected to water stress. New Phytol 148:177–186. doi:10.1046/j.1469-8137.2000.00744.x
Johnson NC, Graham JH, Smith FA (1997) Functioning of mycorrhizal associations along the mutualism-parasitisms continuum. New Phytol 135:575–586. doi:10.1046/j.1469-8137.1997.00729.x
Kozai T (1991a) Micropropagation under photoautotrophic conditions. In: Debergh P, Zimmerman R (eds) Micropropagation Technology and Application. Kluwer Academic Publ, Dordrecht–Boston–London, pp 447–469
Kozai T (1991b) Photoautotrophic micropropagation. In Vitro Cell Dev Biol Plant 27:47–51. doi:10.1007/BF02632127
Kozai T, Iwanami Y (1988) Effects of CO2 enrichment and sucrose concentration under high photon flux on plantlet growth of carnation (Dianthus caryophyllus L.) in tissue culture during the preparation stage. J Jpn Soc Hortic Sci 57:279–288. doi:10.2503/jjshs.57.279
Kozai T, Kubota C (2001) Developing a photoautotrophic micropropagation system for woody plants. J Plant Res 114:525–537. doi:10.1007/PL00014020
Kozai T, Kubota C (2005) Concepts, definitions, ventilation methods, advantages and disadvantages. In: Kozai T, Afreen F, Zobayed SMA (eds) Photoautotrophic (sugar-free medium) micropropagation as a new micropropagation and transplant production system. Springer, Dordrecht, The Netherlands, pp 19–30
Kozai T, Koyama Y, Watanabe I (1988) Multiplication and rooting of potato plantlets in vitro with sugar-free medium under high photosynthetic photon flux. Acta Hortic 230:121–127
Kozai T, Kubota C, Jeong BR (1997) Environmental control for the large-scale production of plants through in vitro techniques. Plant Cell Tissue Organ Cult 51:49–56. doi:10.1023/A:1005809518371
Kozai T, Nguyen QT, Xiao Y (2006) A commercialized photoautotrophic micropropagation system using large vessels with forced ventilation: plant growth and economic benefits. ISHS Acta Horticulturae 725: V international symposium on in vitro culture and horticultural breeding
Kubota C, Kozai T (1992) Growth and net photosynthetic rate of Solanum tuberosum in vitro under forced ventilation. HortScience 27:1312–1314
Li XL, George E, Marschner H (1997) Phosphorus acquisition of VA mycorrhizal hyphae from compacted soil in clover. J Can Botony 75:723–729
Liu WK, Yang QC (2006) Integrated environment control system for closed plantlet production using photoautotrophic micropropagation: cleanliness control subsystem. The 18th international symposium & exhibition on contamination control, Beijing, China, September 6th–8th, 2006, pp 1305–1312
Liu WK, Yang QC, Wu GD, Wei Q (2006) Integrated environment control system for closed plantlet production using photoautotrophic micropropagation: design and performance. J Shenyang Agric Univ 37:455–458
Louche-Tessandier D, Samson G, Hernández-Sebastiá C, Chagvardieff P, Desjardins Y (1999) Importance of light and CO2 on the effects of endomycorrhiza colonization on growth and photosynthesis of potato plantlets (Solanum tuberosum) in an in vitro tripartite system. New Phytol 142:539–550. doi:10.1046/j.1469-8137.1999.00408.x
Lovato P, Guillemin JP, Gianinazzi S (1996) Application of commercial abuscular endomycorrhizal fungal inoculants to the establishment of micropropagated grapevine rootstock and pineapple plants. Agronomie 12:673–880
Mark GL, Murphy J, Cassells AC (1997) Microbial characterisation and preparation of inoculants for in vitro mycorrhization of strawberry in autotrophic culture. In: Cassells AC (ed) Pathogen and microbial contamination management in micropropagation. Kluwer Academic Publishers, The Netherlands, pp 345–350
Murashige T, Skoog F (1962) A revised medium for rapid growth and bioassays with tobacco tissue cultures. Physiol Plant 15:473–493. doi:10.1111/j.1399-3054.1962.tb08052.x
Newsham KK, Fitter AH, Watkinson AR (1995) Multifunctionality and biodiversity in arbuscular mycorrhizas. Trends Ecol Evol 10:407–411. doi:10.1016/S0169-5347(00)89157-0
Nowak J (1998) Benefits of in vitro” biotization’’of plant tissue cultures with microbial inoculants (Review). In Vitro Cell Dev Biol Plant 34:122
Nowak J, Shulaev V (2003) Priming for transplant stress resistance in in vitro propagation. In Vitro Cell Dev Biol Plant 39:107–124. doi:10.1079/IVP2002403
Nguyen QT, Kozai T, Niu GH, Nguyen UV (1999) Photosynthetic characteristics of coffee (Coffea arabusta) plantlets in vitro in response to different CO2 concentrations and light intensities. Plant Cell Tissue Organ Cult 55:133–139. doi:10.1023/A:1006224026639
Nhut DT, Takamura T, Watanabe H, Murakami A, Murakami K, Tanaka M (2002) Sugar-free micropropagation of Eucalyptus citriodora using light-emitting diodes (LEDs) and film-rockwool culture system. Environ Contr Biol 40:147–155
Pons J, Gianinazzi-Pearson V, Gianinazzi S, Navatel JC (1983) Studies on VA-mycorrhizae in vitro: mycorrhizal synthesis of axenically propagated wild cherry (Prunus avium L.). Plant Soil 71:217–221. doi:10.1007/BF02182657
Rai MK (2001) Current advances in mycorrhization in micropropagation. In Vitro Cell Dev Biol Plant 37:158–167
Sahay NS, Varma A (2000) A biological approach towards increasing the rates of survival of micropropagated plants. Curr Sci 78(2):127–129
Saito M, Marumoto T (2002) Inoculation with arbuscular mycorrhizal fungi: the status quo in Japan and the future prospects. Plant Soil 244:273–279. doi:10.1023/A:1020287900415
Salamanca CP, Herrera MA, Barea JM (1992) Mycorrhizal inoculation of micro-propagated woody legumes used in re-vegetation programes for desertified Mediterranean ecosystems. Agronomie 12:869–872. doi:10.1051/agro:19921023
Smith SE, Read DJ (1997) Mycorrhizal symbiosis, 2nd edn. Academic Press, California, pp 11–32
Subhan S, Sharmila P, Saradi PP (1998) Glomus fasiciculatum alleviates transplantation shock of micropropagated Sesbania sesban. Plant Cell Rep 17:268–272. doi:10.1007/s002990050390
Tanaka M, Goi M, Higashiura T (1988a) A novel disposable film culture vessel in micropropagation. Acta Hortic 226:663–670
Tanaka M, Jinno K, Goi M, Higashiura T (1988b) The use of disposable fluorocarbon polymer film culture vessel in micropropagation. Acta Hortic 230:73–80
Tanaka M, Nagae S, Takamura T, Kusanagi N, Ujike M, Goi M (1996) Efficiency and application of film culture system in the in vitro production of plantlets in some horticultural crops. J Soc High Technol Agric 8:280–285
Tanaka M, Yap DCH, Ng CKY, Hew CS (1999) The physiology of Cymbidium plantlets cultured in vitro under conditions of high carbon dioxide and low photosynthetic photon density. J Hortic Sci Biotechnol 74:632–638
Teixeira da Silva JA, Giang DDT, Tanaka M (2006) Photoautrophic micropropagation of Spathiphyllum. Photosynthetica 44:53–61. doi:10.1007/s11099-005-0158-z
van Aarle IM, Olsson PA, Söderström B (2002) Arbuscular mycorrhizal fungi respond to the substrate pH of their extraradical mycelium by altered growth and root colonization. New Phytol 155:173–182. doi:10.1046/j.1469-8137.2002.00439.x
van der Heijden MGA, Boller T, Wiemken A, Sanders IR (1998a) Different arbuscular mycorrhizal fungi species are potential determinants of plant community structure. Ecology 79:2082–2091
van der Heijden MGA, Klironomos JN, Ursic M, Moutogolis P, Streitwolf-Engel R, Boller T et al (1998b) Mycorrhizal fungal diversity determines plant biodiversity, ecosystem variability and productivity. Nature 396:69–72. doi:10.1038/23932
Varma A, Schūepp H (1995) Mycorrhization of the commercially important micropropagated plants. Crit Rev Biotechnol 15:313–328. doi:10.3109/07388559509147417
Vestberg M, Estaún V (1994) Micropropagated plants, an opportunity to positively manage mycorrhizal activities. In: Gianinazzi S, Schüepp H (eds) Impact of arbuscular mycorrhizas on sustainable agriculture and natural ecosystems. Birkhäuser Verlag, Basel, Switzerland, pp 217–225
Vestberg M, Cassells AC, Schubert A, Cordier C, Gianinazzi S (2002) AMF and micropropagation of high value crops. In: Gianinazzi S, Schüepp H, Barea JM, Haselwandter K (eds) Mycorrhizal technology in agriculture. Birkhäuser Verlag, Basel, pp 223–233
Vestberg M, Kukkonen S, Saari K, Parikka P, Huttunen J, Tainio L et al (2004) Microbial inoculation for improving the growth and health of micropropagated strawberry. Appl Soil Ecol 27:243–258. doi:10.1016/j.apsoil.2004.05.006
Voets L, Boulois DH, Renard L, Strullu D, Declerck S (2005) Development of an autotrophic culture system for the in vitro mycorrhizaqtion of potato plantlets. FEMS Microbiol Lett 248:111–118. doi:10.1016/j.femsle.2005.05.025
Wang B, Funakoshi DM, Dalpe Y, Hamel C (2002) Phosphorus-32 absorption and translocation to host plants by arbuscular mycorrhizal fungi at low root-zone temperature. Mycorrhiza 12:93–96. doi:10.1007/s00572-001-0150-9
Xiao YL, Kozai T (2004) Commercial application of a photoautotrophic micropropagation system using large vessels with forced ventilation. HortScience 39(6):1387–1391
Xiao Y, Zhao J, Kozai T (2000) Practical sugar-free micropropagation system using large vessels with forced ventilation. In: Kubota C, Chun C (eds) Transplant Production in the 21st Century. Kluwer Academic Publishers, The Netherlands, pp 266–273
Zobayed SMA, Afreen F, Xiao Y, Kozai T (2001) Recent advancement research on photoautotrophic micropropagation using large culture vessels with forced ventilation. In Vitro Cell Dev Biol Plant 40:450–458
Acknowledgements
We thank three anonymous reviewers for their helpful comments. Also we want to thank Dr. Da Ou yang for his language help. We thank the financial support by the President Fund of Chinese Academy of Agricultural Sciences (JBKY0032007025CAAS).
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Liu, WK., Yang, QC. Integration of mycorrhization and photoautotrophic micropropagation in vitro: feasibility analysis for mass production of mycorrhizal transplants and inoculants of arbuscular mycorrhizal fungi. Plant Cell Tiss Organ Cult 95, 131–139 (2008). https://doi.org/10.1007/s11240-008-9425-6
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11240-008-9425-6