
Abstract
Interspecific hybridization between evergreen pot azalea (Tsutsusi) cultivars and genotypes of other Rhododendron subgenera or sections (Rhododendron, Hymenanthes, Pentanthera, Vireya) is significantly hampered by many prezygotic and postzygotic barriers. The objective of our work was to overcome spontaneous abortion and lack of endosperm formation and to increase germination rates by establishing an embryo rescue protocol. The optimal germination medium for immature Rhododendron seeds was a basal medium supplemented with 145 μM GA3. This medium induced germination of fertilized ovules after several sexual combinations of subgenera. Its use was clearly more efficient than in vivo sowing. The direction of the cross significantly influenced the occurrence of abortion, germination and albinism. The obtained seedlings were multiplied on Woody Plant Medium + 4.5 μM 2iP, and rooted afterwards. Finally, about 9% of the germinated ovules resulted in vigorously growing seedlings that were successfully acclimatized.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Interspecific hybridization is a possible tool for the introduction of extensive variation in ornamental crops (Van Tuyl and De Jeu 1997; Eeckhaut et al. 2006). The difficulty of creating interspecific hybrids increases along with the phylogenetic distance between the parents (Sharma 1995).
Many prezygotic and postzygotic barriers significantly hamper sexual recombination between evergreen azalea (Tsutsusi, TS) cultivars and other Rhododendron types. The genus Rhododendron ( ± 1000 species) is divided into 8 subgenera, the four most important subgenera being Tsutsusi (TS, evergreen azaleas except Brachycalyx section), Pentanthera (PE, deciduous azaleas), Rhododendron (RH, lepidote Rhododendrons) and Hymenanthes (HY, elepidote Rhododendrons) (Chamberlain et al. 1996). In our study, the section Vireya (VI) is considered as a separate group. In the past, numerous efforts have been undertaken to breed an evergreen yellow flowering azalea through interspecific hybridization. Prezygotic incongruity totally inhibits interspecific hybridization after PE × TS (Ureshino et al. 2000), PE × VI, TS × VI and RH × VI pollinations (Rouse 1993). Nevertheless, most incongruity problems have a postzygotic nature. Abortion and low hybrid vigour occur frequently. Albinism is regular, even after intrasubgeneric pollinations (Michishita et al. 2001; Kita et al. 2005). Preil and Ebbinghaus (1985) pollinated azalea with a number of wild Rhododendron species, but fruit development or germinating seeds were seldomly obtained.
Endosperm absence or retarded development is often the cause of spontaneous abortion of hybrid embryos after interspecific pollinations (Van Tuyl et al. 1991; Pickersgill et al. 1993; Sharma 1995). Johnston et al. (1980) published the endosperm balance number theory to explain the basis for normal seed production but currently there is no consensus on the requirements for the formation of hybrid endosperm. Using in vitro media, which replace the endosperm and allow maturation of the hybrid embryo, can overcome the lack of endosperm. For this reason, embryo rescue offers a promising tool for interspecific breeding as an aid in hybridization by sexual means (Zenkteler 1990; Pickersgill et al. 1993; Sharma, 1995; Sharma et al. 1996; De Jeu 2000; Eeckhaut et al. 2006). The main factors affecting its success are the genotype and growth conditions of the mother plant, the developmental stage of the embryo at isolation, the composition of the nutrient media and the culture environmental conditions (oxygen, light and temperature; Pierik 1999). In Japan, evergreen R. indicum hybrids could be crossed with deciduous azaleas thanks to culture of immature seeds, provided the evergreen azalea was the seed parent (Ureshino et al. 1998).
The objectives of our work were threefold. Firstly, we wanted to establish an optimal in vitro germination medium for evergreen azalea seeds, that would break down seed dormancy and enable to harvest fruits 3 months after pollination instead of awaiting full maturity after 9 months. Thus, spontaneous abortion on the seed parent after interspecific pollination can be avoided for the greater part. Our second aim was to evaluate the actual potential of this medium with regard to germination rates of seeds generated by interspecific crosses between evergreen azalea and genotypes belonging to other subgenera of the genus Rhododendron. Finally, we verified whether multiplication and acclimatization of the obtained seedlings were possible. Molecular evaluation of the alleged hybrids was not yet performed.
Materials and methods
General culture practices
Preceding in vitro initiation of immature seeds, Rhododendron fruits were rinsed in 70% ethanol, sterilized for 15 min in a 10% NaOCl solution with 0.005% teepol and finally rinsed 3 times in autoclaved water. The basal medium (BM) was based on Lloyd and McKown Woody Plant Medium (1980) supplemented with 111 μM sucrose and solidified with 7 g/l MC29 agar (pH 5.4). The media were autoclaved (121°C, 30 min, 500 hPa). Petri dishes (φ = 5.5 cm) were filled with 10 ml medium and sealed with low density polyethylene foil. Meli-jars (De Proft et al. 1985) contained 100 ml medium/jar. Cultures were maintained at 23 ± 2°C under a 16 h photoperiod at 40 μMol m−2 s−1 photosynthetic active radiation, supplied by cool white fluorescent lamps (OSRAM L36W/31).
Immature seed germination
After autoclaving, BM was supplemented with 0, 29, 58, 145 or 289 μM GA3 (filter sterilized). On each medium, 120 immature R. simsii ‘Dogwood Bont’ × R. simsii ‘Kingfisher’ seeds (TS × TS, ovaries harvested 3 months after pollination) were initiated in six Petri dishes. The number of germinated seeds was counted after 4 weeks.
Interspecific hybridization
Sixteen evergreen Rhododendron simsii Planch hybrids were selected from the ILVO Rhododendron collection in Melle. This selection included eight Belgian pot azaleas (R. simsii ‘Aline’, ‘Dame Blanche’, ‘Dogwood’, ‘Gerard Kint wit’, ‘Gerda Keessen’, ‘Mevrouw Jozef Heursel’, ‘Mont Blanc’ and ‘Starlight’), five seedlings resulting from crosses within the Belgian pot azalea group (2 ‘Firmin De Waele’ × ‘Dame Blanche’ seedlings, 2 ‘Mevrouw André Heungens’ × ‘Mistral’ seedlings and 1 ‘Mistral’ × ‘Phoenix’ seedling) and three seedlings originated from fully compatible interspecific breeding between Belgian pot azalea and other TS genotypes (R. simsii ‘Phoenix’ × R. obtusum ‘Casablanca tetra’, R. simsii ‘Madame Roggeman’ × R. noriakianum and R. kaempferi × R. simsii ‘Mevrouw Andre Heungens’).
They were reciprocally crossed, whenever possible, with 9 HY, 4 PE, 20 RH and 5 VI genotypes (Table 1), retrieved from as well Glendoick and Esveld nurseries as the ILVO collection. Standard pollination procedures were followed. All plants were 3–8-years-old, depending on their genotype; all pollinations were performed between january and may, based upon the secretion of stigma exudates by the seed parent and the availability of pollen. Seed parents were emasculated 1 day before pollination. For every parental combination, 10 or more flowers were pollinated when possible. The occurrence of possible prezygotic barriers was checked by aniline blue staining (Cuevas et al. 1994).
In vitro sowing
One and a half to two months (VI × TS) or 4–5 months (all other types) after pollination, seeds were collected to prevent spontaneous abortion, and initiated on BM supplemented with the optimal GA3 concentration. Per Petri dish, 20 seeds were sown.
In vivo sowing
For the in vivo control, 5 seed capsules of each combination were left on the mother plant; whenever not aborted, they were harvested respectively 3 months (VI × TS) and 9 months (all other types) after pollination. Seeds were sown in vivo in the greenhouse; germination rates were observed after 2 months.
Multiplication and acclimatization
After evaluation of germination rates (6 weeks after ovule initiation), in vitro seedlings were transferred to multiplication medium (BM + 4.5 μM 2iP). The rooting medium consisted of 1/10 strength MS (Murashige and Skoog 1962) macro and micro salts, 114 μM sucrose, 11.4 μM IAA and 2.5 g/l charcoal (Mertens and Samyn, 1994). Plantlets were acclimatized for 12 weeks after rooting in plastic seedling trays, in a fog unit of the greenhouse (18–20°C, 16 h photoperiod).
Results and discussion
Immature seed germination
As germination of immature seeds is expected to be inhibited by dormancy, the best gibberellin content of the germination medium was determined using a model system. The optimal germination medium for immature seeds/fertilized ovules was BM supplemented with 145 μM GA3 (Fig. 1). This medium resulted in significantly higher germination rates than all other treatments except BM + 289 μM. However, 29 μM was already sufficient to increase the germination rate significantly. These data correspond to the results of similar research (Michishita et al. 2001).

Germination rate of R. simsii ‘Dogwood Bont’ × ‘Kingfisher’ seeds 3 months after pollination on BM + GA3. Means indicated by the same letter are not statistically different (LSD 95%)
Three months after a compatible pollination, the ovules had sufficiently matured to allow successful in vitro germination. To allow in vivo maturation of interspecific embryos on the seed parent as much as possible, it was decided to harvest seed capsules 4–5 months after pollination, as in former experiments, massive abortion usually occurred after 5 months (data not provided). Since VI seed capsules are fully mature 3 months after compatible pollinations (Williams et al. 1991), VI × TS fertilized ovules were initiated 1.5–2 months after pollination.
Interspecific hybridization
Crosses from the PE × TS and TS × VI type suffered from prezygotic barriers and yielded no fertilized ovules. This confirms former results published by Rouse (1993) and Ureshino et al. (2000). Only fruits obtained from TS × HY, HY × TS, TS × PE, TS × RH, RH × TS and VI × TS crosses were harvested.
In vitro sowing
On average, after performing an interspecific cross, 38.8 ovules could be isolated from an immature fruit. The yield was higher after TS × HY crosses, but lower after HY × TS and RH × TS pollinations. In vitro germination rates of fertilized ovules exceeded 20% in HY × TS and TS × RH populations, but were lower than 5% in VI × TS, TS × HY and TS × PE combinations (Table 2). Total albinism was observed in TS × PE hybrids. Other chloroplast deficiencies, mainly virescence, were present after all TS crosses with HY and RH but absent among VI × TS seedlings. Virescence led to a severely decreased biosynthesis of functional chloroplasts and pale green leaf formation, although both cotyledons were green, and consequently resulted in poor growth rates. Seedling vigor was reduced in most subgenus combinations, except for non-virescent RH × TS seedlings, which were relatively vigorous growers. Throughout all ovule culture experiments, no unequivocal correlation between the exact parental genotype and cross efficiency was observed.
The combination of Tsutsusi with any other subgenus or section, except RH, led to clearly different results than the reciprocal pollinations. For as well abortion, germination failure and albinism HY × TS leads to better results than TS × HY. Also for crosses with VI it is recommendable to use the TS genotype as the pollen donor. Due to either prezygotic incongruity or albinism, evergreen and deciduous azaleas could not be hybridized.
In vivo sowing
Due to a combination of fruit abortion, bad seed development and absence of seed germination, all crosses but one were unsuccessful and didn’t yield any seedlings. Five green seedlings were recovered after in vivo sowing of 4 ‘Sunny’ × (‘Mevrouw Andre Heungens’ × ‘Mistral’) mature seed capsules, showing a decreased efficiency compared to ovule culture (23 green seedlings/11 ovaries).
Multiplication and acclimatization
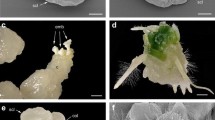
Multiplication of the vigorously growing fraction of the seedlings was not problematic. We suspect that variation in the multiplication and growth rate is not primarily due to a species dependent difference in sensitivity towards the medium, but rather to the interspecific nature of the plant. Indeed, so-called ‘hybrid vigor’ is able to cause dwarfism or drastically decrease vitality (Levin 1978). Many plants were lost during acclimatization. In general, the number of vigorous seedlings obtained 1 year after pollination was higher if the TS genotype was the pollen parent. Finally, about 9% of the germinated ovules resulted in vigorously growing seedlings that were successfully acclimatized; altogether, 10 seedling populations were obtained (Fig. 2).

Seedlings originating from intersubgeneric Rhododendron crosses, 1 year after pollination (bar = 1 cm): A = R. ‘Goldsworth Orange’ × R. ‘Dogwood’, B = R. ‘Nancy Evans’ × R. ‘Dogwood’, C = R. ‘Jingle Bells’ × R. ‘Dogwood’, D = (R. ‘Mevrouw Andre Heungens’ × R. ‘Mistral’) × R. chrysodoron, E = R. chrysodoron × (R. ‘Mevrouw Andre Heungens’ × R. ‘Mistral’), F = R. xanthostephanum × (R. ‘Mevrouw Andre Heungens’ × R. ‘Mistral’), G = R. ‘Azurwolke’ × (R. ‘Firmin De Waele’ × R. ‘Dame Blanche’), H = R. burmanicum × (R. kaempferi × R. ‘Mevrouw Andre Heungens’), I = R. ‘Euan Cox’ × R. ‘Dogwood’, J = R. ‘Sunny’ × (R. ‘Mevrouw Andre Heungens’ × R. ‘Mistral’)
In other words, during acclimatization a rigid selection pressure towards plants that are more autotroph and only moderately suffer from hybrid vigor, is established. In spite of this, the efficiency of in vitro sowing followed by acclimatization is far more successful than in vivo sowing. One or more multiplication cycles are recommended before acclimatization.
Molecular screening is needed to incontestably state that all of these populations are composed of true hybrids. As the aim of our research was to develop a suitable media sequence for embryo rescue in Rhododendron, rather than to elaborate a screening system for the selection of interspecific seedlings, this was not performed during our study. Nevertheless, the occurrence of either albinism or reduced growth vigor in five populations, R. ‘Nancy Evans’ × R. ‘Dogwood’, R. ‘Jingle Bells’ × R. ‘Dogwood’, R. burmanicum × (R. kaempferi × R. ‘Mevrouw Andre Heungens’), R. ‘Euan Cox’ × R. ‘Dogwood’ and R. ‘Sunny’ × (R. ‘Mevrouw Andre Heungens’ × R. ‘Mistral’), is an indication for the interspecific character of the seedlings (Van Tuyl and De Jeu 1997; Eeckhaut et al. 2006).
Conclusions
The optimal medium for immature seed germination obtained after compatible crosses was BM supplemented with 145 μM GA3. Yet 3 months after pollination, ovules can be initiated. Incongruent crosses were performed between evergreen Belgian pot azalea and genotypes from four phylogenetically distant taxonomic groups. The culture of the resulting interspecific embryos was attempted on this medium. Embryo rescue was far more efficient than in vitro sowing, and proved the potential of the medium to enable development of hybrid embryos. Indeed, despite losses through hybrid vigor and albinism, plantlets were successfully multiplied, rooted and acclimatized after four different subgenus combinations; on the other hand, in vitro sowing only yielded viable plantlets after one specific parental combination. We can therefore state that a model system for in vitro culture of interspecific Rhododendron embryos has been established. Further research will focus on efficient screening strategies for hybrid detection and the establishment of F2-generations to characterize introgression.
Abbreviations
- 2iP:
-
Isopentenyladenine
- GA:
-
Gibberellic acid
- HY:
-
Subgenus Hymenanthes
- IAA:
-
Indole acetic acid
- MS:
-
Murashige & Skoog
- PE:
-
Subgenus Pentanthera
- RH:
-
Subgenus Rhododendron
- TS:
-
Subgenus Tsutsusi
- VI:
-
Section Vireya
References
Chamberlain D, Hyam G, Argent G et al (1996) The genus Rhododendron. Its classification & synonymy. Royal Botanical Garden Edinburgh
Cuevas J, Rallo L, Rapoport H et al (1994) Staining procedure for the observation of olive pollen tube behaviour. Acta Hort 356:264–267
De Jeu M (2000) In vitro techniques for ornamental breeding. Acta Hort 508:55–60
De Proft M, Maene L, Debergh P (1985) Carbon dioxide and ethylene evolution in the culture atmosphere of Magnolia cultured in vitro. Physiol Plant 65:375–379
Eeckhaut T, Van Laere K, De Riek J et al (2006) Overcoming interspecific barriers in ornamental plant breeding. In: Teixeira da, Silva J (eds) Floriculture, ornamental and plant biotechnology: advances and topical issues, 1st edn. Global Science Books, London, pp 540–551
Johnston S, Den Nijs T, Peloquin S et al (1980) The significance of gene balance to endosperm development in interspecific crosses. Theor Appl Genet 57:5–9
Kita K, Kurashige Y, Yukawa T et al (2005) Plastid Inheritance and Plastome-genome incompatibility of intergeneric hybrids between Menziesia and Rhododendron. J Japan Soc Hort Sci 74:318–323
Levin D (1978) The origin of isolating mechanisms in flowering plants. Evol Biol 11:185–197
Lloyd G, Mc Cown B (1980) Commercially-feasible micropropagation of mountain laurel, Kalmia latifolia, by use of shoot-tip culture. Proc Int Plant Prop Soc 30:421
Mertens M, Samyn G (1994) Regeneration and somaclonal variation of Rhododendron simsii offer prospects to the obtainment of commercially valuable sports. Mededelingen FLTBW RUG 59:2495–2497
Michishita A, Ureshino K, Miyajima I (2001) Shortening the period from crossing to the seedling stage through ovule culture of interspecific crosses of azalea (Rhododendron spp.). J Japan Soc Hort Sci 70:54–59
Murashige T, Skoog F (1962) A revised medium for rapid growth and bioassays with tobacco tissue cultures. Physiol Plant 15:473–497
Pickersgill B, Hayward M, Bosemark N et al (1993) Interspecific hybridization by sexual means. In: Hayward M, Bosemark N, Romagosa I (eds) Plant breeding: principles and prospects. Plant breeding series. Chapman and Hall, New York
Pierik R (1999) In vitro culture of higher plants. Kluwer Academic Publishers, Dordrecht
Preil W, Ebbinghaus R (1985) Bastardierungen von Topfazaleen (Rhododendron simsii) mit anderen Rhododendron-Arten. Rhododendron und immergrüne Laubgehölze, Jahrbuch pp 85–92
Rouse J (1993) Inter- and intraspecific pollinations involving Rhododendron species. J Amer Rhododendron Soc 47:23–45
Sharma D, Kaur R, Kumar K (1996) Embryo rescue in plants - a review. Euphytica 89:325–337
Sharma H (1995) How wide can a wide cross be? Euphytica 82:43–64
Ureshino K, Miyajima I, Akabane M (1998) Effectiveness of three-way crossing for the breeding of yellow-flowered evergreen azalea. Euphytica 104:113–118
Ureshino K, Kawai M, Miyajima I (2000) Factors of unilateral cross incompatibility between several evergreen azalea species and Rhododendron japonicum flavum. J Japan Soc Hort Sci 69:261–265
Van Tuyl J, De Jeu M (1997) Methods for overcoming interspecific crossing barriers. In: Shivanna K, Sawhney V (eds) Pollen biotechnology for crop production and improvement. University Press, Cambridge, pp 273–292
Van Tuyl J, Van Dien M, Van Creij M et al (1991) Application of in vitro pollination, ovary culture, ovule culture and embryo rescue for overcoming incongruity barriers in interspecific Lilium crosses. Plant Sci 74:115–116
Williams E, Rouse J, Kaul V et al (1991) Reproductive timetable for the tropical Vireya rhododendron R. macgregoriae. Sexual Plant Reprod 4:155–165
Zenkteler M (1990) In vitro fertilization and wide hybridization in higher plants. Critical Rev Plant Sci 9:267–279
Acknowledgements
This work was financed by DWTC-Belgium. The authors wish to thank Romain Uytterhaegen, Ronald Van den Oord, Kristien Janssens, Veerle Cools and Laurence Desmet for technical assistance.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Eeckhaut, T., De Keyser, E., Van Huylenbroeck, J. et al. Application of embryo rescue after interspecific crosses in the genus Rhododendron . Plant Cell Tiss Organ Cult 89, 29–35 (2007). https://doi.org/10.1007/s11240-007-9209-4
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11240-007-9209-4