
Abstract
Nematodes of the genus Rhabdias Stiles & Hassall, 1905 (Rhabditoidea: Rhabdiasidae) have a dioecious free-living stage and a hermaphroditic stage that parasitises the lungs of amphibians and reptiles. Approximately 94 species of Rhabdias have been described. Because the similar morphological characteristics such as the labial structures, the location of the vulva and the shape of the tail of Rhabdias spp. hinder their identification, molecular biology techniques and scanning electron microscopy have been employed to diagnose species of this genus. This study describes Rhabdias breviensis n. sp., parasitic in the lungs of two Neotropical frog species Leptodactylus petersii Steindachner and Leptodactylus macrosternum Miranda-Ribeiro. The description of this species integrates classical taxonomy, scanning electron microscopy and a molecular analysis of the mitochondrial COI gene. The new species differs from all other Rhabdias species parasitic in Neotropical hosts in certain morphometric parameters, the position of the vulva, the host group and the cephalic characters.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
The genus Rhabdias includes approximately 94 species distributed worldwide (Kuzmin & Tkach, 2012). The life-cycle of these nematodes consists of a dioecious free-living stage and a hermaphroditic stage that parasitises the lungs of amphibians and reptiles (Vicente et al., 1991; Anderson, 2000). Several researchers, including Kuzmin et al. (2003), Tkach et al. (2006), Kuzmin et al. (2007), Martínez-Salazar (2008) and Martínez-Salazar et al. (2009), have suggested that the similarity in the characters used to identify Rhabdias individuals at the species level leads to confusion and misidentification. Therefore, complementary approaches using molecular biology techniques, scanning electron microscopy, biogeographical analysis and identification of the host species are necessary to correctly identify species of this genus.
The application of molecular biology techniques to taxonomy, systematics and phylogeny has resulted in the reassessment of many species of Rhabdias and the reassignment of some previously described species (Lupchinski Jr. et al., 2006; Kuzmin et al., 2007). Taxonomic and phylogenetic studies of this type frequently use the nuclear ribosomal markers 5.8S, 28S and 18S rDNA and the two internal transcribed spacers (ITS1 and ITS2) in addition to mitochondrial markers such as cytochrome b (cyt b), cytochrome c oxidase subunit I (COI) and 12S rRNA genes (Dorris et al., 2002; Dare et al., 2008; Dubey & Shine, 2008; Lhermitte-Vallarino et al., 2009).
In Brazil, species of Rhabdias have been recorded in anurans belonging to the following families: Bufonidae Gray, Ranidae Rafinesque, Leptodactylidae Werner, Hylidae Rafinesque, and Microhylidae Günther (see Kung & Wu, 1945). Only one species has been reported in the latter family i.e. Rhabdias globocephala Kung & Wu, 1945 from Microhyla ornata Duméril & Bibron (see Kung & Wu, 1945).
The species of the leptodactylid genus Leptodactylus Fitzinger are distributed throughout South America, Southern North America and the West Indies (Frost, 2013). According to Vicente et al. (1991), the following species of Leptodactylus act as hosts for nematodes of the genus Rhabdias in South America: Leptodactylus chaquensis Cei (Rhabdias sp.), Leptodactylus labyrinthicus Spix (Rhabdias fuelleborni Travassos, 1926), Leptodactylus syphax Bokermann (Rhabdias sp.), Leptodactylus podicipinus Cope (Rhabdias sp.), Leptodactylus melanonotus Hallowell (Rhabdias elegans Gutierrez, 1945), and Leptodactylus ocellatus Linnaeus (Rhabdias sp.).
Among the species of Leptodactylus only L. petersii Steindachner and L. macrosternum Miranda-Ribeiro inhabit the Amazon Basin and have been reported to host parasitic helminths. Leptodactylus macrosternum, whose synonyms include L. ocellatus and Leptodactylus latrans Steffen (see Heyer et al., 2012), is the only host of Rhabdias spp. that has been reported in this region (Vicente et al., 1991; Goldberg et al., 2009).
This study describes a new species of the genus Rhabdias parasitising two leptodactylid frog species (L. petersii and L. macrosternum) in Breves Municipality, Marajó Island (eastern Amazon). The description combines light and scanning electron microscopy observations with molecular data for the COI gene.
Materials and methods
Eleven specimens of L. petersii and three specimens of L. macrosternum were collected in January 2011 in peridomiciliary areas at Breves Municipality, Marajó Island, Pará State, eastern Amazon, Brazil (1°40′55″S, 50°28′48″W).
Morphological data
The nematodes found in frog lungs were fixed in AFA (2% glacial acetic acid, 3% formaldehyde and 95% ethanol) at 60°C, dehydrated and cleared in an increasing ethanol/glycerine series, and examined under an Olympus BX41 microscope equipped with a drawing tube (with no zoom adjustment) to obtain drawings for subsequent morphometric analysis. All measurements are in micrometres unless otherwise stated and are expressed as the range followed by the mean and standard deviation in parentheses. After fixation in AFA, specimens from both host species were postfixed in 1% osmium tetroxide, dehydrated in acetone up to the carbon dioxide (CO2) critical point and coated with gold for subsequent analysis by scanning electron microscopy (LEO 1450 and LEO 1430 instruments).
Molecular data
Nematode DNA was obtained by phenol/chloroform extraction following the standard procedures of Sambrook et al. (1989). A fragment of the mitochondrial COI gene of each sample was amplified by polymerase chain reaction (PCR) in a total reaction volume of 25 μL containing 10 ng of genomic DNA, 1.0 mM of each dNTP, 3.0 mM of MgCl2, 1 unit of Taq DNA polymerase and 5 μM of each primer [HCO (5′-TAA ACT TCA GGG TGA CCA AAA AAT CA-3′) and LCO (5′-GGT CAA CAA ATC ATA AAG ATA TTG G-3′) (Folmer et al., 1994)]. The amplification profile consisted of 30 cycles comprising 1 min at 95°C, 1 min at 55°C and 1 min at 72°C. These cycles were preceded by an initial 2-min step at 95°C to homogenise the temperature of the block and followed by a final 5-min step at 72°C to polymerise any remaining molecules from which the polymerase may have dissociated before the fragment was fully synthesised.
Amplification products were cloned into the pGEM-T Vector (Promega®), inserted into electrocompetent Escherichia coli TOP10 (Invitrogen®) and sequenced using an automated DNA analyser [model ABI 3130, Applied Biosystems] in combination with the BigDye Terminator V3.1 kit according to the manufacturer’s instructions. To confirm the observed mutations, each sample was sequenced with both primers, forward and reverse. The nucleotide sequences were edited and aligned using the BioEdit software (Hall, 2007).
-
Order Rhabditida Chitwood, 1933
-
Family Rhabdiasidae Railliet, 1916
Rhabdias breviensis n. sp.
Type-host: Leptodactylus petersii Steindachner.
Other host: Leptodactylus macrosternum Miranda-Ribeiro.
Type-locality: Breves Municipality, Marajó Island, Pará State, Brazil (1°40′55″S, 50°28′48″W).
Site of infection: Lungs.
Type-specimens: Holotype MPEG 00010, paratype MPEG 00011 and paratype MPEG 00012, deposited in the Museu Paraense Emílio Goeldi (MPEG), Belém, Pará State, Brazil.
Prevalence: In L. petersii: 63.64%; in L. macrosternum: 100%.
Mean abundance: In L. petersii: 30 (range 20–40); in L. macrosternum: 36 (range 30–45).
Molecular data: Three sequences of the mitochondrial COI gene (706 nt) were obtained, two from specimens from L. petersii and one from L. macrosternum. All three sequences shared a single haplotype (GenBank accession number KC512382). No termination codons or indels were observed.
Etymology: The species is named after the type-locality, Breves Municipality.
Description (Figs. 1, 2)
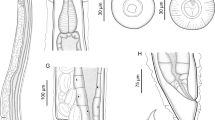
[Based on the holotype and nineteen paratypes (Fig. 1A–E)]. Anterior third of hermaphroditic female truncated, posterior extremity curved dorsally (Fig. 1A–B). Total body length 2.63–3.63 (3.14 ± 0.34) mm; width near vulva region 370–543 (423 ± 59); width at lower oesophageal dilation 145–187 (160 ± 12).

Rhabdias breviensis n. sp. A, overall view of the body. B, anterior third of the body, showing the goblet-shaped buccal capsule and the nerve-ring located in the anterior third of the claviform oesophagus; C, posterior third of the body, showing the conical shape of the tail; D, lateral view of the post-equatorial vulva; E, embryonated eggs with thin shells. Scale-bars: A, C, D, 200 μm; B, 50 μm; E, 20 μm
Buccal capsule goblet-shaped (Fig. 1A–B), 4–9 (6 ± 1) × 7–13 (11 ± 2). Oesophagus claviform (Fig. 1B), 238–410 (355 ± 44) × 45–54 (51 ± 2) (width measured in the ganglia region).
Nerve-ring (Fig. 1B) located at 41–84 (61 ± 10) from anterior extremity. Vulva post-equatorial (Fig. 1C), 1,870–2,348 (1,988 ± 478) from the anterior extremity of the nematode [65–71 (63)% of body length]; tail short, 139–191 (160 ± 14).
Hermaphroditic females amphidelphic, with larvae found near the vulvar opening and eggs distant from this aperture (Fig. 1D–E). Embryonated eggs measure on average 81 ± 4 × 43 ± 0.3.
Scanning electron microscopy data
Scanning electron microscopy observations provided additional morphological details for the new species from the lungs of L. petersii and L. macrosternum. The body is covered by a delicate inflation of the cuticle extending from the oral aperture to the end of the tail (Fig. 2A–D). Two pairs of submedian cephalic papillae and one pair of lateral cephalic papillae are present at the anterior extremity, in addition to two lateral amphids. The oral aperture is circular and surrounded by four submedian and two lateral lips (Fig. 2A–B). The submedian cephalic papillae are spherical in shape and located on the internal portion of the submedian lips, whereas the lateral cephalic papillae are conical in shape and located on the internal portion of the lateral lips (Fig. 2A–B). The larvae present in the uterus are covered by thin shells (Fig. 2E–F).

Scanning electron micrographs of Rhabdias breviensis n. sp. A, frontal view of the anterior extremity, showing the six lips surrounding the circular oral aperture, the submedian cephalic papillae (arrowheads), the lateral cephalic papillae (asterisks) and two lateral amphids (arrows); B, detail of the anterior extremity, showing the submedian and lateral cephalic papillae; C, frontal-lateral view of the posterior third of the body, showing the conical tail; D, extremity of the conical tail, showing the inflation of the cuticle covering this region; E, embryonated eggs, showing the thin egg-shells covering the larvae; F, detail of the larvae without their egg-shells. Scale-bars: A, F, 10 μm; B, 2 μm; C, 30 μm; D, 5 μm; E, 20 μm
Discussion
Rhabdias breviensis n. sp. is a parasite of L. petersii and L. macrosternum which inhabit the same niche, the peridomiciliary area. This shared niche likely favours the transmission cycle and the adaptation of this nematode to both species of Leptodactylus. The concomitant parasitism of both hosts was inferred from the morphological data and confirmed by the molecular data i.e. the COI gene sequences of the lung parasites from L. macrosternum and L. petersii were identical thus confirming the parasitism by R. breviensis n. sp. in these distinct hosts. This finding is important because it corroborates the statements of Martínez-Salazar (2008) and Martínez-Salazar et al. (2009) that Rhabdias spp. usually do not parasitise hosts belonging to different families and are selective with regard to the genus of the host. Similarly, Tkach et al. (2006) observed that Rhabdias bakeri Walton, 1929 and Rhabdias ranae Walton, 1929 parasitised both Rana sylvatica LeConte and Lithobates pipiens Schreber (= Rana pipiens); this observation was later confirmed by Dare et al. (2008) through experimental infections and molecular biology studies.
A wide variety of Rhabdias spp. parasitise amphibians and reptiles, but only four have been reported from Neotropical frogs: Rhabdias mucronata Schuurmans-Stekhoven, 1952 from Leptodactylus bolivianus Boulenger, R. truncata Schuurmans-Stekhoven, 1952 from Telmatobius schreiteri Vellard, R. tobagoensis Moravec & Kaiser, 1995 from Pristimantis charlottesvillensis Kaiser, Dwyer, Feichtinger & Schmid, and R. savagei Bursey & Goldberg, 2005 from Lithobates forreri Boulenger (see Moravec & Kaiser, 1995; Bursey & Goldberg, 2005; Kuzmin & Tkach, 2012).
Rhabdias breviensis n. sp. differs from R. savagei in having six lips around the oral aperture vs four lips (see Bursey & Goldberg, 2005). Additionally, the new species exhibits reduced morphometric dimensions compared with R. savagei and R. tobagoensis. Despite these morphometric differences, R. breviensis n. sp. resembles R. tobagoensis in the number of lips surrounding the oral aperture. These two species differ in the location of the vulva, which is post-equatorial in R. breviensis n. sp. and equatorial in R. tobagoensis (see Moravec & Kaiser, 1995).
No data are available in the literature for the hermaphroditic forms R. mucronata and R. truncata parasitic in amphibians, however only juvenile forms were described by Schuurmans-Stekhoven (1952); this hinders a comparison with R. breviensis n. sp.
When compared with the Neotropical species of Rhabdias that parasitise different host groups, R. breviensis n. sp. exhibits the same number of lips surrounding the oral aperture as Rhabdias anolis Bursey, Goldberg & Telford, 2003, R. vellardi Pereira, 1928, R. lamothei Martínez-Salazar & León-Règagnon, 2006, R. nicaraguensis Bursey, Goldberg & Vitt, 2007 and Rhabdias fuelleborni Travassos, 1926 (see Bursey et al., 2003, 2007; Martínez-Salazar & León-Règagnon, 2007). However, R. breviensis n. sp. differs markedly from these species in its morphology, morphometric dimensions and host group. Furthermore, R. anolis, R. nicaraguensis, R. vellardi and R. lamothei are parasites of reptiles (Anolis frenatus Cope, Anolis capito Peters, Philodryas schottii Girard, and Leptodeira maculata Hallowell, respectively) and R. fuelleborni parasitises toads and frogs (Rhinella marina Linnaeus, R. schneideri Werner, R. arenarum Hense, R. icterica Spix and Thoropa miliaris Spix) (Bursey et al., 2003; Martínez-Salazar & León-Règagnon, 2007; Kuzmin & Tkach, 2012).
The post-equatorial position of the vulva suggests that R. breviensis n. sp. is taxonomically closer to five Neotropical species, i.e. Rhabdias androgynia Kloss, 1971, R. elegans Gutierrez, 1945, R. hermaphrodita Kloss, 1971, R. leonae Martínez-Salazar, 2006, and R. paraensis Santos, Melo, Nascimento, Nascimento, Giese & Furtado, 2011. However, the new species differs from these taxa in its host group and cephalic characteristics. In addition, the cuticle of R. breviensis n. sp. lacks pores or canaliculi connecting these pores to the hypodermal cells, characteristics observed by Santos et al. (2011) in R. paraensis. Thus, R. breviensis n. sp. differs from all other Rhabdias spp. parasitising Neotropical hosts in terms of its morphometric dimensions, the position of the vulva, the host group and certain cephalic characteristics. This is the first report of a species of Rhabdias from L. petersii.
References
Anderson, R. C. (2000). Nematode parasites of vertebrates: Their development and transmission (2nd ed.). Wallingford, Oxon, UK: CAB International.
Bursey, C. R., & Goldberg, S. R. (2005). New species of Oswaldocruzia (Nematoda: Molineoidae), new species of Rhabdias (Nematoda: Rhabdiasidae), and other helminths in Rana cf. forreri (Anura: Ranidae) from Costa Rica. Journal of Parasitology, 91, 600–605.
Bursey, C. R., Goldberg, S. R., & Telford Jr., S. R. (2003). Rhabdias anolis n. sp. (Nematoda: Rhabdiasidae) from the lizard, Anolis frenatus (Sauria: Polychrotidae), from Panama. Journal of Parasitology, 89, 113–117.
Bursey, C. R., Goldberg, S. R., & Vitt, L. J. (2007). New species of Rhabdias (Nematoda: Rhabdiasidae) and other helminths from Norops capito (Sauria: Polychrotidae) from Nicaragua. Journal of Parasitology, 93, 129–131.
Dare, O. K., Nadler, S. A., & Forbes, M. R. (2008). Nematode lungworms of two species of anuran amphibians: Evidence for co-adaptation. International Journal for Parasitology, 38, 1,729–1,736.
Dorris, M., Viney, M. E., & Blaxter, M. L. (2002). Molecular phylogenetic analysis of the genus Strongyloides and related nematodes. International Journal for Parasitology, 32, 1,507–1,517.
Dubey, S., & Shine, R. (2008). Origin of the parasites of an invading species, the Australian cane toad (Bufo marinus): Are the lungworms Australian or American? Molecular Ecology, 17, 4,418–4,424.
Folmer, O., Black, M., Hoeh, W., Lutz, R., & Vrijenhoek, R. (1994). DNA primers for amplification of mitochondrial cytochrome c oxidase subunit I from diverse metazoan invertebrates. Molecular Marine Biology and Biotechnology, 3, 5, 294–299.
Frost, D. R. (2013). Amphibian Species of the World: an Online Reference. Version 5.6 (9 January 2013). Electronic Database accessible at http://research.amnh.org/herpetology/amphibia/index.html. American Museum of Natural History, New York, USA.
Goldberg, S. R., Bursey, C. R., Caldwell, J. P., & Shepard, D. B. (2009). Gastrointestinal helminths of six sympatric species of Leptodactylus from Tocantins State, Brazil. Comparative Parasitology, 76, 258–266.
Hall, B. G. (2007). Phylogenetic Trees Made Easy: A How-to Manual (3rd ed.). Sunderland, MA: Sinauer Associates.
Heyer, R., Langone, J., La Marca, E., Azevedo-Ramos, C., Di Tada, I., Baldo, D., et al. (2012). Leptodactylus latrans. In: IUCN 2012. IUCN red list of threatened species. Version 2012.2. www.iucnredlist.org. Accessed 11 November 2012.
Kung, C. C., & Wu, H. W. (1945). Parasitic nematodes of amphibians from Pehpei Szechwan, China. Sinensia, 16, 73–83.
Kuzmin, Y., & Tkach, V. V. (2002–2012). Rhabdias. World Wide Web electronic publication. Accessed 4 February 2012.
Kuzmin, Y., Tkach, V. V., & Brooks, D. R. (2007). Two new species of Rhabdias (Nematoda: Rhabdiasidae) from the marine toad, Bufo marinus (L.) (Lissamphibia: Anura: Bufonidae), in Central America. Journal of Parasitology, 93, 159–165.
Kuzmin, Y., Tkach, V. V., & Snyder, S. D. (2003). The nematode genus Rhabdias (Nematoda: Rhabdiasidae) from amphibians and reptiles of the Nearctic. Comparative Parasitology, 70, 101–114.
Lhermitte-Vallarino, N., Barbuto, M., Junker, K., Boistel, R., Ineich, I., Wanji, S., et al. (2009). Rhabdias rhampholeonis n. sp. and Rhabdias mariauxi n. sp. (Nematoda, Rhabdiasoidea), first lung worms from leaf chameleons: Description, molecular evidence and notes on biology. Parasitology International, 58, 375–383.
Lupchinski Jr., E., Vargas, L., Ribeiro, R. P., Moreira, H. L. M., Valentim, M., & Povh, J. A. (2006). A importância da utilização da técnica RAPD para a identificação de dactilogirídeos em Tilápias do Nilo (Oreochromis niloticus). Arquivos de Ciências Veterinárias e Zoologia, 9, 49–57.
Martínez-Salazar, E. A. (2008). A new rhabdiasid species from Craugastor occidentalis (Anura: Brachycephalidae) from Sierra de Manantlán, Jalisco, Mexico. Revista Mexicana de Biodiversidad, 79, 81–89.
Martínez-Salazar, E. A., & León-Règagnon, V. (2007). New species of Rhabdias (Nematoda: Rhabdiasidae) from Bufo occidentalis (Anura: Bufonidae) from Sierra Madre del Sur, Mexico. Journal of Parasitology, 93, 1,171–1,177.
Martínez-Salazar, E. A., León, G. P. P., & Olea, G. P. (2009). First record of the genus Rhabdias (Nematoda: Rhabdiasidae), endoparasite from Scinax staufferi (Anura: Hylidae) in Mexico. Revista Mexicana de Biodiversidad, 80, 861–865.
Moravec, F., & Kaiser, H. (1995). Helminth parasites from West Indian frogs, with descriptions of two new species. Caribbean Journal of Science, 31, 252–268.
Sambrook, J., Fritsch, E. F., & Maniatis, T. (1989). Molecular cloning: A laboratory manual. Cold Spring Harbor, NY: Cold Spring Harbor Laboratory Press.
Santos, J. N., Melo, F. T. V., Nascimento, L. C. S., Nascimento, D. E. B., Giese, E. G., & Furtado, A. P. (2011). Rhabdias paraensis sp. nov.: A parasite of the lung of Rhinella marina (Amphibia: Bufonidae) from Brazilian Amazonia. Memórias do Instituto Oswaldo Cruz, 106, 433–440.
Schuurmans-Stekhoven, J. H. S. (1952). Nematodos parasitos de anfíbios, pájaros y mamíferos de la República Argentina. Acta Zoologica Lilloana, 10, 315–400.
Tkach, V. V., Kuzmin, Y., & Pulis, E. E. (2006). A new species of Rhabdias from lungs of the wood frog, Rana sylvatica, in North America: The last sibling of Rhabdias ranae? Journal of Parasitology, 92, 631–636.
Vicente, J. L., Rodrigues, H. O., Gomes, D. C., & Pinto, R. M. (1991). Nematoides do Brasil. 2ª parte: Nematoides de anfíbios. Revista Brasileira de Zoologia, 7, 549–561.
Acknowledgements
The authors thank Dr. Hilton Túlio Costi of the Institutional Laboratory of Scanning Electron Microscopy at the Paraense Emílio Goeldi Museum (Museu Paraense Emílio Goeldi, MPEG), Dr. Claudio Nery Lamarão of the Laboratory of Scanning Electron Microscopy at the Institute of Geosciences, UFPA, for their technical support, Dr. Marinus Steven Hoogmoed, Museu Paraense Emílio Goeldi, for identifying the amphibian specimens, Dr. Yurii Kuzmin, I.I.Schmalhausen Institute of Zoology, for discussion on the cephalic characters of the genus Rhabdias, and Caroline Melo, Emanuelle Argolo, Thaís Reis and Fabrísia Silva d’ Encarnação for their technical support in sample collection. Financial support was provided by PROCAD-NF 2009 [Programa Nacional de Cooperação Acadêmica – Ação Novas Fronteiras (National Program of Academic Cooperation – New Frontiers)], a special program of CAPES [Coordenação de Aperfeiçoamento de Pessoal de Nível Superior (Coordination for the Improvement of Higher Education Personnel)]; SISBiota 2010 [Sistema Nacional de Pesquisa em Biodiversidade (National Research System in Biodiversity)], a program of CNPq [Conselho Nacional de Desenvolvimento Científico e Tecnológico (National Council for Scientific and Technological Development]; PROPESP [Pró-Reitoria de Pesquisa e Pós-Graduação (Dean of Research and Postgraduate Studies)]; and FADESP [Fundação de Amparo e Desenvolvimento da Pesquisa (Foundation for Research Development)], divisions of UFPA; and Parasitologia Básica 2010 (Basic Parasitology), a special program of CAPES.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
de Cássia Silva do Nascimento, L., Gonçalves, E.C., de Vasconcelos Melo, F.T. et al. Description of Rhabdias breviensis n. sp. (Rhabditoidea: Rhabdiasidae) in two Neotropical frog species. Syst Parasitol 86, 69–75 (2013). https://doi.org/10.1007/s11230-013-9432-9
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11230-013-9432-9