
Abstract
Objective
The goal of this paper is to discuss cancer-related fatigue (CRF) and address issues related to the investigation into potential biological and genetic causal mechanisms. The objectives are to: (1) describe CRF as a component of quality of life (QOL); (2) address measurement issues that have slowed progress toward an understanding of mechanisms underlying this symptom; (3) review biological pathways and genetic approaches that have promise for the exploration of causal mechanisms of CRF; and (4) offer directions for future research.
Methods
Review, synthesis, and interpretation of the literature.
Results
Until recently, CRF and QOL have been understood primarily as subjective patient-reported experiences. With increased understanding of human genetics, theories and research are being expanded to incorporate biological and genetic understandings of these subjective experiences. Proposed biological and genetic mechanisms of CRF that have been examined include cytokine dysregulation, hypothalamic–pituitary–adrenal (HPA) axis dysfunction, five hydroxy tryptophan (5-HT) neurotransmitter dysregulation, circadian rhythm disruption, alterations in adenosine triphosphate (ATP) and muscle metabolism, and vagal afferent activation. Approaches to the study of genetic mechanisms have also been addressed including candidate genes, genome-wide scanning, and gene expression. Based on the review and synthesis of the literature, directions for future research are proposed.
Conclusions
Understanding the biological and genetic basis of CRF has the potential to contribute to a more complete understanding of the genetic determinants of QOL.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
The goal of this paper is to discuss fatigue as a component of quality of life (QOL) and address issues related to the investigation into potential biological and genetic causal mechanisms. There is a large body of research on fatigue that has focused on it as a cancer-related symptom, so this discussion will address fatigue in the context of cancer. First, we will describe cancer-related fatigue (CRF) as a component of QOL. Next, we will address measurement issues that have slowed progress toward an understanding of mechanisms underlying this highly prevalent symptom. This will be followed by a review of biological pathways that have promise for the exploration of causal mechanisms. Finally, we will offer directions for future research. Elucidating the genetic basis for CRF has the potential to contribute to a more complete understanding of the genetic basis of QOL.
We are on the brink of a new understanding of subjective experiences such as CRF and QOL. To date, these constructs have been understood largely as subjective patient-reported experiences. This limited view is probably due in part to outdated notions of the mind–body dichotomy in which subjective experiences are relegated to the domain of the mind as separate from the body or biological domain. As our understanding of genetics has grown, it has become clear that genetic mechanisms are involved in a broad range of human functions including subjective experiences such as symptoms and QOL. Clearly, our theories and research must be expanded to incorporate biological as well as psychological and behavioral determinants of subjective experience. This paper will highlight some of the issues that need to be addressed in this transition using the specific example of CRF.
Fatigue and quality of life
Cancer-related fatigue (CRF) is a highly prevalent symptom. The incidence ranges from 25 to 99% of patients depending on the means of assessment and which patients are assessed [1]. Fatigue is not confined to the active phase of cancer. It has been identified as prevalent in survivors with 17–30% reporting fatigue [2, 3]. Fatigue has been identified as a multidimensional construct including physical and mental fatigue, activity reduction, and motivation reduction [4]. Fatigue has also been coupled with decrements in physical, psychological, and social functioning that are also important domains of QOL. The high incidence of fatigue is coupled with distress; patients have reported fatigue to be the most distressing symptom they have experienced [5, 6], even more so than pain [7].
The link between CRF and health-related quality of life has been described both conceptually and empirically. The adapted model of Wilson and Cleary [8] that provides the conceptual framework for this series of papers proposes that symptoms and the resultant functional decrements impact overall quality of life [9]. As a highly prevalent symptom resulting in significant functional deficits, CRF can diminish QOL profoundly. The proposed associations between CRF and QOL have been demonstrated empirically. A study of breast cancer patients showed that those who reported clinically significant CRF (cases) at the conclusion of treatment had worse physical and social functioning and greater mood disturbance than those who did not report CRF (non-cases) [10]. Another study of breast cancer survivors showed that there were large differences between fatigue cases and non-cases in all domains of QOL except subjective perception of cognitive disturbance [11]. Women with clinically significant CRF had worse physical, emotional, and social functioning as well as worse global health status than those without CRF. This study also showed that fatigued survivors had poorer body image and sexual functioning and greater mood disturbance than those without CRF. These examples of CRF and QOL are part of a large body of research, showing that fatigue is strongly linked with health-related quality of life.
Measurement issues
Until recently, a consistent definition of CRF has been lacking, but consensus is building through the work of various consortia and working groups including the National Comprehensive Cancer Network Fatigue Guidelines Committee [12] of what constitutes CRF. Recently, an independent working group, Assessing the Symptoms of Cancer using Patient-Reported Outcomes (ASCPRO), developed a consensus definition of CRF as the perception of unusual tiredness that varies in pattern of severity and has a negative impact on ability to function in people who have or have had cancer [13]. This definition represents the consensus of clinical, academic, and pharmaceutical investigators as well as cancer survivors.
While there is greater agreement about the conceptual definition of CRF, work is still needed to clarify the operational or case definition (See Appendix, Glossary). Most subjective measures of CRF examine the dimensions of severity and impact on functioning [13]. However, some measures include additional dimensions including cognitive [14–18] or emotional fatigue [15, 16, 18–20], energy level [21, 22], or motivation [17]. These variations in measured dimensions of CRF denote a lack of well-defined criteria for distinguishing clinically important cases of CRF from non-cases. Although some work has been done to clarify the case definition of CRF [2, 10, 23, 24], more work is needed. In order for the exploration of molecular and genetic mechanisms of CRF to proceed, a well-defined case definition is essential. Also, for simplicity and feasibility of measurement, subjective measures of CRF need to demonstrate congruence with the case definition.
Pathophysiologic pathways
Interest in the underlying biological pathways and genetic mechanisms of fatigue is mounting. A pioneering study by Alexander and colleagues [11] examined the differences between CRF cases and non-cases with regard to biological and behavioral markers associated with different potential mechanisms of CRF: anemia, inflammatory processes, disruption of the HPA axis, metabolic disturbances, and physical de-conditioning. Thirty percent of breast cancer survivors met CRF criteria. With regard to mechanisms, there were subtle immune and inflammatory disturbances but no differences in indicators of other mechanisms.
The findings above suggest that inquiry into pathophysiologic pathways and genetic mechanisms hold promise for the identification of causal mechanisms and potential treatment targets. The next section will review current knowledge of pathways that have been associated with the development and persistence of cancer-related fatigue as well as current evidence for the genetic control of cancer-related fatigue. The search strategy consisted of a Medline search to identify articles on the etiology, genetics, and physiopathology of CRF. The articles identified were supplemented by articles previously identified by the authors.
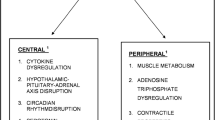
The pathophysiologic mechanisms involved in cancer-related fatigue are not completely understood [25]. Dysregulation of several systems, both biochemical and physiological, are likely involved in CRF and include both peripheral and central mechanisms [26]. Peripheral fatigue originates in the muscles and related tissues, whereas central fatigue develops in the central nervous system and results in progressive failure to transmit motor neuron impulses. Proposed mechanisms of CRF include cytokine dysregulation, hypothalamic–pituitary–adrenal (HPA) axis dysfunction, five hydroxy tryptophan (5-HT) neurotransmitter dysregulation, circadian rhythm disruption, alterations in adenosine triphosphate (ATP) and muscle metabolism, and vagal afferent activation [26, 27].
Cytokine dysregulation
Proinflammatory cytokines such as interleukin (IL)-1β, IL-6, and tumor necrosis factor (TNF)-α have been evaluated as markers of cancer-related fatigue [28, 29]. Breast cancer survivors with persistent fatigue 2 or more years after treatment had elevated levels of immune markers associated with proinflammatory cytokine activity [28]. Other changes in immunity have included lower levels of natural killer cells and natural killer cell activity associated with higher levels of CRF [30–32].
Cancer and its treatment have been associated with increases in plasma cytokines [26]. The mechanisms, however, are not fully understood. Research has demonstrated that the tumor micro-environment contains pro-inflammatory cytokines such as IL-1, IL-6, and TNF-α [33]. For various cancer diagnoses, elevated levels of circulating cytokines and their receptors (most commonly IL-6) have been observed at initial diagnosis and after diagnosis of metastasis. These cytokines can contribute to anemia, cachexia, anorexia, fever, infection, and depression, all of which can contribute to fatigue [34]. In addition to acting as a growth factor for the neoplasm, specific cytokines may contribute to fatigue through more specific pathways. IL-1 and IL-6 and TNF-α have been shown to suppress erythropoiesis, resulting in anemia [34]; TNF has been associated with alterations in central nervous system neurotransmission [26]; and interleukins, tumor necrosis factor, interferon-γ, and leukemia inhibitory factor can act as chachectins [34, 35]. Cachexia can result in tissue wasting and resulting fatigue, and interferon can produce neurasthenia, a neurological fatigue [36]. Cytokines, especially IL-6, have also been associated with depression, with fatigue as a manifestation [37, 38].
Schubert and colleagues examined the link between CRF and specific inflammatory markers [39]. Conclusions of this review were limited by the small number of studies, small sample sizes, heterogeneity of study populations, and inconsistency in selection of inflammatory markers for study. However, general analyses based on weighting of sample size demonstrated a significant positive correlation between fatigue and levels of circulating inflammatory markers. Specific markers that were associated with fatigue included IL6, IL-1ra, and neopterin. Although IL-1β or TNF-α have been proposed as markers in the fatigue pathway, neither correlated with fatigue in several studies of cancer-related fatigue [40–42]. The findings to date provide initial support for further exploration of the inflammatory pathway in the etiology of fatigue.
Hypothalamic–pituitary–adrenal axis dysfunction
The hypothalamic–pituitary–adrenal (HPA) axis controls the release of cortisol in response to physical or psychosocial stress [26]. Specifically, corticotropin-releasing hormone (CRH) is secreted from the paraventricular nucleus of the hypothalamus and acts with vasopressin to release adrenocorticotropic hormone (ACTH) from the anterior pituitary. ACTH stimulates the release of cortisol from the adrenal cortex. The HPA axis also influences the development of immune cells and cytokine production. Low levels of cortisol are associated with fatigue and may result from the direct suppression of the HPA axis by cancer treatment or changes in 5-HT that result in decreased stimulation of 5-HT1A receptors that signals the release of CRH [43].
In healthy people, there is a diurnal pattern of cortisol production with peak levels prior to awakening and a steady decline throughout the day [44]. Breast cancer survivors with persistent fatigue (in comparison with non-fatigued survivors) had a flatter cortisol slope and significant cortisol elevation in the late evening [45]. These results suggest that alterations in HPA axis activity could influence fatigue.
Dysregulation of serotonin
Another hypothesis is that increased levels of 5-HT (serotonin) in the brain and/or upregulation of 5-HT receptors due to cancer or cancer treatment could result in fatigue [26]. A suggested mechanism for this increased fatigue is that activity of proinflammatory cytokines including interleukin, interferon, IFN-γ, and TNF-α may stimulate indoleamine 2,3-dioxygenase to alter 5-HT metabolism. Increases in 5-HT levels or upregulation of 5-HT receptors signals the release of CRH, in turn modifying the HPA axis function to reduce somatomotor drive and produce a sensation of decreased physical ability. In animal studies, 5-HT levels were shown to increase during sustained exercise; fatigue was observed at the highest concentration of 5-HT [46]. Research in humans on exercise-induced fatigue and chronic fatigue syndrome suggests that 5-HT dysregulation may be implicated in the etiology of fatigue [47].
The observed occurrence of fatigue and depression together has led to a proposed common pathophysiological mechanism involving 5-HT regulation in cancer patients [48]. Two related clinical trials examined this hypothesis. The first trial randomized 479 fatigued patients receiving chemotherapy for any cancer diagnosis to receive paroxetine hydrochloride (Paxil), a selective serotonin reuptake inhibitor, 20 mg or placebo for 8 weeks [49]. The second trial conceptually replicated the first trial in 94 breast cancer patients [48]. Neither trial showed benefit in reducing fatigue; however, both trials demonstrated a significant effect of the drug on depressed mood. The results do not support the hypothesis that central serotonin regulation is a primary mechanism of cancer-related fatigue during chemotherapy.
Circadian rhythm disruption
Circadian rhythms could play a significant role in the etiology of fatigue through the modulation of arousal and sleep [42]. The central circadian system and downstream network of relay stations in the hypothalamus regulates arousal and sleep. This system is most significantly affected by the input of light that creates neurophysiological changes in the hypothalamus and melatonin that affects the darkness period, neurons in the suprachiasmatic nuclei (SCN) that regulate 24-h sleep patterns, and by afferent nerve fibers groups that relay signals from the SCN to the hypothalamus. Signals from the hypothalamus affect the parasympathetic and sympathetic autonomic centers in the brainstem and in turn affect the secretion of stress and physiology regulating hormones. Circadian rhythm may also be affected through SCN downstream signal disruption that may occur in the dorsal or ventral nuclei or from input from the brain’s visceral, limbic, and cortical systems.
Alterations in any part of the circadian system can result in disruption of arousal and sleep patterns. SCN ablation in animals has resulted in the permanent loss of rhythmic rest, activity, and sleep behaviors. Research has shown that fatigue in cancer patients undergoing treatment was inversely related to activity during the day and restless sleep at night [50, 51]. SCN peptides and ligands of epidermal growth factor (EGF) receptor, released by the cancer or in response to the stress of the cancer, have been shown to reversibly inhibit activity and deregulate 24-h sleep patterns [42]. Specific peptides include EGF, TGF-α, and neuregulin-1, prokineticine-2, and cardiotrophin-like cytokine. Individuals with dampened or inconsistent circadian rhythms had higher levels of fatigue than those with consistent rhythms [26]. Circadian rhythms have been affected significantly in advanced stage or metastatic disease. Little or no effect is noted with early-stage cancers. Specific mediators of circadian rhythms, including cortisol levels and melatonin, have been shown to change in patients with cancer [42].
Alterations in adenosine triphosphate
Dysregulation of ATP, the muscle’s major source of energy, can result in peripheral fatigue. It is hypothesized that cancer or its treatment, anorexia, or cachexia can lead to defects in the ability to regenerate ATP in skeletal muscles. This may be through the mechanism of altered muscle protein metabolism [26]. Research investigating chronic fatigue syndrome has demonstrated defects in ATP synthesis and metabolism. Evidence of ATP dysregulation in cancer patients is limited. However, a randomized clinical trial of ATP infusions for advanced lung cancer patients resulted in improved muscle strength and reduced fatigue [52].
Vagal afferent nerve activation
Cancer or its treatment may cause release of peripheral neuroactive agents that activate vagal afferent fibers with suppression of somatic muscle activity and induction of “sickness behavior” in animals [26]. Based on the existing data, it has been proposed that reflex inhibition of skeletal muscle activity due to proinflammatory cytokines or 5-HT could be responsible for cancer-related fatigue [33, 53]. There is little evidence to support this hypothesis although the results of a few animal studies provide some support for this mechanism [26].
Summary
The discussion of multiple pathophysiological pathways suggests that CRF is multidimensional [5–7]. While we do not thoroughly understand all of the pathways, a number of them have been identified as playing a role in CRF [25]. It is possible that different pathophysiological mechanisms are responsible for different dimensions of fatigue. For instance, the existence of peripheral fatigue may be a result of ATP dysregulation while circadian rhythm may contribute mainly to mental fatigue. More generalized forms of fatigue could involve central mechanisms such as cytokine dysregulation. As fatigue severity increases, it is possible that more pathways will be involved. Further delineation of the aspects of fatigue associated with each of these pathways will assist in the identification of new intervention targets more specifically aimed at the type of fatigue experienced.
Genetics and fatigue
Advances in molecular and genetic technology now enable the use of different methods for identifying the genes that control the development and persistence of cancer-related fatigue. Genome-wide scanning can be done without a specific hypothesis or proposed pathway because thousands of single nucleotide polymorphisms (SNP) can be examined simultaneously. Cancer-related symptoms are likely to be influenced by the cumulative effect of several gene polymorphisms [54]. Also cytokine genes are pleiotropic in that the activity of one gene can have more than one effect [55]. Also, genes that control other cancer-related symptoms (such as depression or pain) could influence fatigue [54–56]. Genome-wide scanning would be useful in identifying combinations of genes that control fatigue.
Another type of association study is hypothesis driven with candidate genes examined in specific pathways. The pathway-based approach requires prior knowledge of pathophysiologic processes and the formulation of specific hypotheses about gene SNP that are likely to regulate the development of specific symptoms such as fatigue. The hypothesis-driven candidate gene approach can be used to provide evidence in support of specific hypotheses about the mechanisms of cancer-related fatigue.
Collado-Hidalgo used the candidate gene approach to test whether variation in fatigue was influenced by gene polymorphisms in the regulatory regions (promoters) of the genes that encode proinflammatory cytokines [57]. SNP in the promoter region of IL-1β (−511) and IL-6 (−174) have been shown to control gene expression of cytokines associated with breast cancer susceptibility and prognosis. These candidate genes were evaluated as risk factors for fatigue. IL-1β −511 and IL-6 −174 were each independently associated with post-treatment fatigue in breast cancer survivors. There was an overrepresentation of the IL-1 CC allele and an underrepresentation of the TT allele among survivors with persistent fatigue. There was also an increased occurrence of homozygosity for the C allele variant and the wild-type G allele for the IL-6 −174 (G/C) polymorphism in fatigued survivors. The results suggest that increased production of proinflammatory cytokines are involved in the development of CRF.
Several cytokine genes and their polymorphisms have been proposed as candidate markers for the study of CRF [54].
-
IL-1B—511 (C/T): As noted previously, there was overrepresentation of the CC alleles and underrepresentation of TT alleles among fatigued survivors [54, 57].
-
IL-6—174 (G/C): It was previously noted that homozygosity for both the variant C allele and the wild-type G allele of the IL-6—174 (G/C) polymorphism was overrepresented in the fatigued group [40–42, 54, 57].
-
TNF-α—308 (G/A): This polymorphism has been proposed for involvement in neoplastic cachexia which is characterized, in part, by fatigue [54]. However, none of the studies that examined the association between TNF-α and fatigue demonstrated an association [30, 58].
-
IL8—251 (T/A): Variation in the promoter SNP IL8—251 (T/A) was found to affect risk for pain severity in newly diagnosed untreated lung cancer patients by influencing concentrations of IL8 [54, 55]. The association of this gene with fatigue is unknown.
-
IL2—330 (T/G): This genetic polymorphism is believed to regulate IL-2 production which has been implicated in complex regional pain syndrome and painful neuropathy [54]. Its association with CRF is unknown.
An alternative to analyzing DNA using SNP is examining the transcript of genes, RNA. RNA synthesis is the process by which DNA nucleotide sequence information is transcribed into RNA nucleotide sequence information. If the DNA codes for a protein, transcription is the first step that leads to the expression of a gene. This is done by the production of mRNA, a “chemical blueprint” for a protein product. While genetic variation in DNA is constant throughout life, RNA changes over time and is found to be tissue specific; not all genes are similarly expressed in all tissue.
Among the studies that have addressed global gene expression and fatigue, a common problem is the lack of statistical precision. The data sets are small, and measures to correct for multiple testing have not been performed, or at least not described. Generally, methods have not been described in detail making inferences of the findings difficult. One could also argue that gene expression in blood cells is not the optimal (but probably easiest accessible) biomarker of fatigue.
Global gene expression has been used in a few studies of non-cancer fatigue. Although this work has not been done specifically in CRF, the findings of a non-cancer-related fatigue syndrome could provide clues about gene expression profiles that would be applicable to CRF. In most cases, gene expression of blood lymphocytes has been used. Whistler and colleagues found 839 genes to be associated with fatigue when contrasting 40 patients with chronic fatigue and 37 healthy controls [59]. The genes regulated oxidative phosphorylation, glucogenesis, lipid metabolism, and several signal transduction pathways. Given the fact that 19,000 probes were analyzed, the problem of multiple tests was substantial. Later, the same group adopted a different approach by using candidate genes [60]. Gene sets for T- and B-cell regulation were constructed, and expression was contrasted between cases with fatigue and healthy controls. The results pointed at a B-cell dysfunction in the blood array profiles.
Kerr adopted a two-stage approach first identifying 182 genes related to fatigue when examining the global gene expression of lymphocytes in 25 cases with chronic fatigue syndrome and myalgic encephalomyelitis and 50 controls [61]. These genes were validated in 55 cases and 75 controls and also demonstrated an association with immune response, cancer, and cell death.
A study available through the CDC (public health genomics office) evaluated 227 individuals from Wichita with chronic fatigue syndrome [62]. SNPs in HPA axis–associated genes, in neuroendocrine effector and receptor genes, and in genes in the serotonergic system were found to be related to fatigue.
Implications for future research
The preceding discussion was intended to broaden our understanding of mechanisms that could be responsible to the development and persistence of CRF. The next section will suggest directions for future research.
Multiple clinical trials are currently underway to determine the efficacy of interventions to prevent and/or treat CRF [13]. As the science continues to progress toward individualized medicine, understanding the genetic dimensions of CRF will become increasingly important in the design of specific interventions to prevent and combat CRF.
Several avenues of research are recommended. First, work is needed to further delineate the case definition of CRF. This includes determining whether there are CRF sub-types such as CRF with or without clinical depression. Also more work is needed to examine the similarities and dissimilarities of CRF and other chronic fatigue syndromes. Such comparisons would further our understanding of the mechanisms behind different fatigue syndromes. This work is critical in developing a clear and specific of CRF phenotype as a basis for studying CRF genotypes. Related to the case definition of CRF is the need for further research linking patient-reported measures of CRF to the case definition.
A second area requiring further investigation is the biological and genetic mechanisms of CRF to supplement our more robust understanding of the subjective experience. Biological markers of CRF need to be identified, so these can be tracked as part of the natural history of this symptom along with subjective reports of CRF severity and impact. Future research must elucidate the relationship of these biological markers and currently known or postulated pathophysiologic pathways involved in CRF with emerging understanding of genetics, genes, and their expression in CRF. As medical science continues to progress toward individualized medicine, understanding the genetic dimensions of CRF will become increasingly important in the design of specific interventions to prevent and combat CRF.
Summary
This paper has examined issues that need to be addressed to increase our understanding of CRF as a component of QOL as well as current evidence for biological pathways and genetic mechanisms that could regulate the development and persistence of cancer-related fatigue. Further research is needed to expand our understanding of the biology and genetics of fatigue in order to identify persons at risk for this debilitating symptom as well as targets for intervention to alleviate it.
References
Higginson, I. J., Armes, J., & Krishnasamy, M. (2004). Introduction, in fatigue in cancer. In J. Armes, M. Krishnasamy, & I. J. Higginson (Eds.), Fatigue in cancer (pp. xvii–xxi). Oxford: Oxford University Press.
Cella, D., Davis, K., Breitbart, W., & Curt, G. (2001). Cancer-related fatigue: prevalence of proposed diagnostic criteria in a United States sample of cancer survivors. Journal of Clinical Oncology, 19(14), 3385–3391.
Servaes, P., Verhagen, C., & Bleijenberg, G. (2002). Fatigue in cancer patients during and after treatment: Prevalence, correlates and interventions. European Journal of Cancer, 38(1), 27–43.
Smets, E. M., Garssen, B., Cull, A., & de Haes, J. C. (1996). Application of the multidimensional fatigue inventory (MFI-20) in cancer patients receiving radiotherapy. British Journal of Cancer, 73(2), 241–245.
Curt, G. A., Breitbart, W., Cella, D., Groopman, J. E., Horning, S. J., Itri, L. M., et al. (2000). Impact of cancer-related fatigue on the lives of patients: New findings from the Fatigue Coalition. The Oncologist, 5(5), 353–360.
Lawrence, D. P., Kupelnick, B., Miller, K., Devine, D., & Lau, J. (2004). Evidence report on the occurrence, assessment, and treatment of fatigue in cancer patients. Journal of the National Cancer Institute Monographs, 32, 40–50.
Vogelzang, N. J., Breitbart, W., Cella, D., Curt, G. A., Groopman, J. E., Horning, S. J., et al. (1997). Patient, caregiver, and oncologist perceptions of cancer-related fatigue: Results of a tripart assessment survey. The Fatigue Coalition. Seminars in Hematology, 34(3 Suppl 2), 4–12.
Wilson, I. B., & Cleary, P. D. (1995). Linking clinical variables with health-related quality of life. A conceptual model of patient outcomes. JAMA, 273(1), 59–65.
Sprangers, M. A. G., Sloan, J. A., Barsevick, A., Chauhan, C., Dueck, A. C., Raat H., et al. (2010). Scientific Imperatives, clinical implications, and theoretical underpinnings for the investigation of the relationship between genetic variables and patient-reported quality-of-life outcomes. Quality of Life Research. doi:10.1007/s11136-010-9759-5.
Andrykowski, M. A., Schmidt, J. E., Salsman, J. M., Beacham, A. O., & Jacobsen, P. B. (2005). Use of a case definition approach to identify cancer-related fatigue in women undergoing adjuvant therapy for breast cancer. Journal of Clinical Oncology, 23(27), 6613–6622.
Alexander, S., Minton, O., Andrews, P., & Stone, P. (2009). A comparison of the characteristics of disease-free breast cancer survivors with or without cancer-related fatigue syndrome. European Journal of Cancer, 45(3), 384–392.
NCCN. (2010). The complete library of NCCN clinical practice guidelines in oncology. Fort Washington, PA: National Comprehensive Cancer Network.
Barsevick, A. M., Cleeland, C. S., Manning, D. C., O’Mara, A. M., Reeve, B. B., Scott, J. A., et al. (2010). ASCPRO Recommendations for the assessment of fatigue as an outcome in clinical trials. Journal of Pain and Symptom Management, 39(6), 1086–1099.
Mendoza, T. R., Wang, X. S., Cleeland, C. S., Morrissey, M., Johnson, B. A., Wendt, J. K., et al. (1999). The rapid assessment of fatigue severity in cancer patients: Use of the brief fatigue inventory. Cancer, 85(5), 1186–1196.
Piper, B. F., Lindsey, A. M., & Dodd, M. J. (1989). The development of an instrument to measure the subjective dimension of fatigue. In E. M. T. S. G. Funk & M. T. Campagne (Eds.), Key aspects of comfort: Management of pain, fatigue, and nausea (pp. 199–208). New York: Springer.
Schwartz, A. L. (1998). The schwartz cancer fatigue scale: Testing reliability and validity. Oncology Nursing Forum, 25(4), 711.
Smets, E. M., Garssen, B., Bonke, B., & De Haes, J. C. (1995). The multidimensional fatigue inventory (MFI) psychometric qualities of an instrument to assess fatigue. Journal of Psychosomatic Research, 39(3), 315–325.
Stein, K. D., Martin, S. C., Hann, D. M., & Jacobsen, P. B. (1998). A multidimensional measure of fatigue for use with cancer patients. Cancer Practice, 6(3), 143–152.
Belza, B. L. (1995). Comparison of self-reported fatigue in rheumatoid arthritis and controls. Journal of Rheumatology, 22(4), 639–643.
Okuyama, T., Akechi, T., Kugaya, A., Okamura, H., Shima, Y., Maruguchi, M., et al. (2000). Development and validation of the cancer fatigue scale: A brief, three-dimensional, self-rating scale for assessment of fatigue in cancer patients. Journal of Pain and Symptom Management, 19(1), 5–14.
Lee, K. A., Hicks, G., & Nino-Murcia, G. (1991). Validity and reliability of a scale to assess fatigue. Psychiatry Research, 36(3), 291–298.
Ware, J. E., Jr., Snow, K. K., Kosinski, M., & Gandek, B. (1993). SF-36 health survey manual and interpretation guide. Boston: The Health Institute, New England Medical Center.
Alexander, S., Minton, O., Stone, P. C., Alexander, S., Minton, O., & Stone, P. C. (2009). Evaluation of screening instruments for cancer-related fatigue syndrome in breast cancer survivors. Journal of Clinical Oncology, 27(8), 1197–1201.
Sadler, I. J., Jacobsen, P. B., Booth-Jones, M., Belanger, H., Weitzner, M. A., & Fields, K. K. (2002). Preliminary evaluation of a clinical syndrome approach to assessing cancer-related fatigue. Journal of Pain and Symptom Management, 23(5), 406–416.
Gutstein, H. B. (2001). The biologic basis of fatigue. Cancer, 92(6 Suppl), 1678–1683.
Ryan, J. L., Carroll, J. K., Ryan, E. P., Mustian, K. M., Fiscella, K., & Morrow, G. R. (2007). Mechanisms of cancer-related fatigue. The Oncologist, 12(Suppl 1), 22–34.
Morrow, G. R., Andrews, P. L., Hickok, J. T., Roscoe, J. A., Matteson, S., Morrow, G. R., et al. (2002). Fatigue associated with cancer and its treatment. Supportive Care in Cancer, 10(5), 389–398.
Bower, J. E., Ganz, P. A., Aziz, N., Fahey, J. L., Cole, S. W., Bower, J. E., et al. (2003). T-cell homeostasis in breast cancer survivors with persistent fatigue. Journal of the National Cancer Institute, 95(15), 1165–1168.
Greenberg, D. B., Gray, J. L., Mannix, C. M., Eisenthal, S., & Carey, M. (1993). Treatment-related fatigue and serum interleukin-1 levels in patients during external beam irradiation for prostate cancer. Journal of Pain and Symptom Management, 8(4), 196–200.
Bower, J. E., Ganz, P. A., Aziz, N., & Fahey, J. L. (2002). Fatigue and proinflammatory cytokine activity in breast cancer survivors. Psychosomatic Medicine, 64(4), 604–611.
Levy, S., Herberman, R., Lippman, M., & d’Angelo, T. (1987). Correlation of stress factors with sustained depression of natural killer cell activity and predicted prognosis in patients with breast cancer. Journal of Clinical Oncology, 5(3), 348–353.
Levy, S. M., Herberman, R. B., Maluish, A. M., Schlien, B., & Lippman, M. (1985). Prognostic risk assessment in primary breast cancer by behavioral and immunological parameters. Health Psychology, 4(2), 99–113.
Seruga, B., Zhang, H., Bernstein, L. J., & Tannock, I. F. (2008). Cytokines and their relationship to the symptoms and outcome of cancer. Nature Reviews Cancer, 8(11), 887–899.
Kurzrock, R. (2001). The role of cytokines in cancer-related fatigue. Cancer, 92(6 Suppl), 1684–1688.
Menzies, H., Chochinov, H., & Breitbart, W. (2005). Cytokines, cancer, and depression: Connecting the dots [comment]. Journal of Supportive Oncology, 3, 55–57.
Quesada, J. R., Talpaz, M., Rios, A., Kurzrock, R., & Gutterman, J. U. (1986). Clinical toxicity of interferons in cancer patients: a review. Journal of Clinical Oncology, 4(2), 234–243.
Maes, M., Meltzer, H. Y., Bosmans, E., Bergmans, R., Vandoolaeghe, E., Ranjan, R., et al. (1995). Increased plasma concentrations of interleukin-6, soluble interleukin-6, soluble interleukin-2 and transferrin receptor in major depression. Journal of Affective Disorders, 34(4), 301–309.
Musselman, D. L., Miller, A. H., Porter, M. R., Manatunga, A., Gao, F., Penna, S., et al. (2001). Higher than normal plasma interleukin-6 concentrations in cancer patients with depression: preliminary findings. American Journal of Psychiatry, 158(8), 1252–1257.
Schubert, C., Hong, S., Natarajan, L., Mills, P. J., Dimsdale, J. E., Schubert, C., et al. (2007). The association between fatigue and inflammatory marker levels in cancer patients: A quantitative review. Brain, Behavior, and Immunity, 21(4), 413–427.
Ahlberg, K., Ekman, T., & Gaston-Johansson, F. (2004). Levels of fatigue compared to levels of cytokines and hemoglobin during pelvic radiotherapy: A pilot study. Biological Research Nursing, 5(3), 203–210.
Meyers, C. A., Albitar, M., Estey, E., Meyers, C. A., Albitar, M., & Estey, E. (2005). Cognitive impairment, fatigue, and cytokine levels in patients with acute myelogenous leukemia or myelodysplastic syndrome. Cancer, 104(4), 788–793.
Rich, T. A. (2007). Symptom clusters in cancer patients and their relation to EGFR ligand modulation of the circadian axis. The Journal of Supportive Oncology, 5(4), 167–174; discussion 176–177.
Bender, C. M., Sereika, S. M., Brufsky, A. M., Ryan, C. M., Vogel, V. G., Rastogi, P., et al. (2007). Memory impairments with adjuvant anastrozole versus tamoxifen in women with early-stage breast cancer. Menopause, 14(6), 995–998.
Posener, J. A., Schildkraut, J. J., Samson, J. A., & Schatzberg, A. F. (1996). Diurnal variation of plasma cortisol and homovanillic acid in healthy subjects. Psychoneuroendocrinology, 21(1), 33–38.
Bower, J. E., Ganz, P. A., Aziz, N., Bower, J. E., Ganz, P. A., & Aziz, N. (2005). Altered cortisol response to psychologic stress in breast cancer survivors with persistent fatigue. Psychosomatic Medicine, 67(2), 277–280.
Bailey, S. P., Davis, J. M., & Alhlborn, E. N. (1993). Neuroendocrine and substrate responses to altered brain 5-HT activity during prolonged exercise to fatigue. Journal of Applied Physiology, 74, 3006–3012.
Andrews, P. L. R., Morrow, G. R., & Hickok, J. T. (2004). Mechanisms and models of fatigue associated with cancer and its treatment. Evidence of pre-clinical and clinical studies. In J. Armes, M. Krishnasamy, & I. Higginson (Eds.), Fatigue in cancer (pp. 51–87). Oxford: Oxford University Press.
Roscoe, J. A., Morrow, G. R., Hickok, J. T., Mustian, K. M., Griggs, J. J., Matteson, S. E., et al. (2005). Effect of paroxetine hydrochloride (Paxil) on fatigue and depression in breast cancer patients receiving chemotherapy. Breast Cancer Research and Treatment, 89(3), 243–249.
Morrow, G. R., Hickok, J. T., Roscoe, J. A., Raubertas, R. F., Andrews, P. L., Flynn, P. J., et al. (2003). Differential effects of paroxetine on fatigue and depression: A randomized, double-blind trial from the University of Rochester Cancer Center Community Clinical Oncology Program. Journal of Clinical Oncology, 21(24), 4635–4641.
Berger, A. M. (1998). Patterns of fatigue and activity and rest during adjuvant breast cancer chemotherapy. Oncology Nursing Forum, 25(1), 51–62.
Mormont, M. C. (1997). Circadian-system alterations during cancer processes: A review. International Journal of Cancer, 70, 241–247.
Agteresch, H. J., Dagnelie, P. C., van der Gaast, A., Stijnen, T., & Wilson, J. H. (2000). Randomized clinical trial of adenosine 5′-triphosphate in patients with advanced non-small-cell lung cancer. Journal of the National Cancer Institute, 92(4), 321–328.
Segerstrom, S. C., & Miller, G. E. (2004). Psychological stress and the human immune system: A meta-analytic study of 30 years of inquiry. Psychological Bulletin, 130(4), 601–630.
Reyes-Gibby, C. C., Wu, X., Spitz, M., Kurzrock, R., Fisch, M., Bruera, E., et al. (2008). Molecular epidemiology, cancer-related symptoms, and cytokines pathway. Lancet Oncology, 9(8), 777–785.
Reyes-Gibby, C. C., Spitz, M., Wu, X., Merriman, K., Etzel, C., Bruera, E., et al. (2007). Cytokine genes and pain severity in lung cancer: Exploring the influence of TNF-alpha-308 G/A IL6–174G/C and IL8–251T/A. Cancer Epidemiology, Biomarkers and Prevention, 16(12), 2745–2751.
Irwin, M. R., Miller, A. H., Irwin, M. R., & Miller, A. H. (2007). Depressive disorders and immunity: 20 years of progress and discovery. Brain, Behavior, and Immunity, 21(4), 374–383.
Collado-Hidalgo, A., Bower, J. E., Ganz, P. A., Irwin, M. R., & Cole, S. W. (2008). Cytokine gene polymorphisms and fatigue in breast cancer survivors: Early findings. Brain, Behavior, and Immunity, 22, 1197–1200.
Shafqat, A., Einhorn, L. H., Hanna, N., Sledge, G. W., Hanna, A., Juliar, B. E., et al. (2005). Screening studies for fatigue and laboratory correlates in cancer patients undergoing treatment. Annals of Oncology, 16(9), 1545–1550.
Whistler, T., Taylor, R., Craddock, R. C., Broderick, G., Klimas, N., Unger, E. R., et al. (2006). Gene expression correlates of unexplained fatigue. Pharmacogenomics, 7(3), 395–405.
Aspler, A. L., Bolshin, C., Vernon, S. D., Broderick, G., Aspler, A. L., Bolshin, C., et al. (2008). Evidence of inflammatory immune signaling in chronic fatigue syndrome: A pilot study of gene expression in peripheral blood. Behavioral and Brain Functions Electronic Resource: BBF, 4, 44.
Kerr, J. R., Burke, B., Petty, R., Gough, J., Fear, D., Mattey, D. L., et al. (2008). Seven genomic subtypes of chronic fatigue syndrome/myalgic encephalomyelitis: A detailed analysis of gene networks and clinical phenotypes. Journal of Clinical Pathology, 61(6), 730–739.
Reyes, M., Nisenbaum, R., Hoaglin, D. C., Unger, E. R., Emmons, C., Randall, B., et al. (2003). Prevalence and incidence of chronic fatigue syndrome in Wichita, Kansas. Archives of Internal Medicine, 163(13), 1530–1536.
Plomin, R., DeFries, J. C., McClearn, G. E., & McGuffin, P. (2001). Behavioral Genetics. New York: Worth Publishers and W. H. Freeman and Co.
Hartyl, D. L., & Jones, E. W. (Eds.). (2009). Genetics: Analysis of genes, genomes (7th ed.). Sudbury, Massachusetts: Jones and Bartlett.
Acknowledgments
GENEQOL Consortium participants per March 2009: Amy P. Abertnethy, Duke Cancer Care Research Program, Duke University Medical Center, Durham, NC, US; Frank Baas, Laboratory of Neurogenetics, Academic Medical Center, University of Amsterdam, Amsterdam, The Netherlands; Andrea M. Barsevick, Cancer Prevention and Control Program, Fox Chase Cancer Center, Philadelphia, PA, US; Meike Bartels, Department of Biological Psychology, VU University, Amsterdam, the Netherlands; Dorret I. Boomsma, Department of Biological Psychology, VU University, Amsterdam, the Netherlands; Cynthia Chauhan, Cancer Advocay, Wichita, KS, US; Charles S. Cleeland, Department of Symptom Research, The University of Texas M. D. Anderson Cancer Center, Houston, TX, US; Amylou C. Dueck, Section of Biostatistics, Mayo Clinic, Scottsdale, AZ, US; Marlene H. Frost, Women’s Cancer Program, Mayo Clinic, Rochester, MN, US; Per Hall, Department of Medical Epidemiology and Biostatistics, Karolinska Institute, Stockholm, Sweden; Michele Y. Halyard, Department of Radiation Oncology, Mayo Clinic, Scottsdale, AZ, US; Pål Klepstad, Department of Intensive Care Medicine, St Olavs University Hospital, Norwegian University of Technology and Science, Trondheim, Norway; Nicholas G. Martin, Queensland Institute of Medical Research, Brisbane, Australia; Christine Miaskowski, School of Nursing, University of California, San Francisco, CA, US; Miriam Mosing, Queensland Institute of Medical Research, Brisbane, Australia; Benjamin Movsas, Department of Radiation Oncology, Henry Ford Health System, Detroit, MI, US; Cornelis J. F. Van Noorden, Department of Cell Biology and Histology, Academic Medical Center, University of Amsterdam, Amsterdam, The Netherlands; Donald L. Patrick, Department of Health Services, University of Washington, Seattle, WA, US; Nancy L. Pedersen, Department of Medical Epidemiology and Biostatistics, Karolinska; Institute, Stockholm, Sweden; Mary E. Ropka, Cancer Prevention and Control Program, Fox Chase Cancer Center, Cheltenham, PA, US; Quiling Shi, Department of Symptom Research, The University of Texas M. D. Anderson Cancer Center, Houston, TX, US; Gen Shinozaki, Department of Psychiatry and Psychology, Mayo Clinic, Rochester, MN, US; Jasvinder A. Singh, Minneapolis Veterans Affairs Medical Center and University of Minnesota, Minneapolis, MN and Mayo Clinic College of Medicine, Rochester, MN, US; Jeff A. Sloan, Department of Health Sciences Research, Mayo Clinic, Rochester, MN, US; Mirjam A. G. Sprangers, Department of Medical Psychology, Academic Medical Center, University of Amsterdam, Amsterdam, The Netherlands; Ruut Veenhoven, Faculty of Social Sciences, Erasmus University Rotterdam, Rotterdam, The Netherlands; Ping Yang, Department of Genetic Epidemiology, Mayo Clinic, Rochester, MN, US; Aeilko H. Zwinderman, Department of Clinical Epidemiology and Biostatistics, Academic Medical Center, University of Amsterdam, Amsterdam, The Netherlands.
Author information
Authors and Affiliations
Consortia
Corresponding author
Appendix: Glossary [63, 64]
Appendix: Glossary [63, 64]
-
Case Definition: Criteria or standards used to identify an instance of a disease, syndrome, or condition;
-
DNA (deoxyribonucleic acid): The two-stranded molecule that encodes genetic information;
-
Gene: The basic unit of inheritance. A sequence of DNA bases that codes for a particular product;
-
Genome: All the DNA sequences of an organism;
-
Genotype: The genetic constitution of an organism that is not manifested as outward characteristics;
-
Phenotype: Physical characteristics of an organism or the presence of a disease that may or may not be genetic;
-
Polymorphism: A locus with two or more alleles (alternative forms of a gene at a locus);
-
Functional polymorphism: DNA sequence variations that alter the expression and/or functioning of the gene product;
-
Single nucleotide polymorphism (SNP): Sequences in the genome that differ by a single nucleotide between one portion of the population and another.
Rights and permissions
About this article
Cite this article
Barsevick, A., Frost, M., Zwinderman, A. et al. I’m so tired: biological and genetic mechanisms of cancer-related fatigue. Qual Life Res 19, 1419–1427 (2010). https://doi.org/10.1007/s11136-010-9757-7
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11136-010-9757-7