
Abstract
While subject to illumination, photosystem I (PSI) has the potential to produce reactive oxygen species (ROS) that can cause photo-oxidative damage in oxygenic photoautotrophs. The reaction center chlorophyll in PSI (P700) is kept oxidized in excess light conditions to limit over-excitation of PSI and alleviate the production of ROS. Oxidation of P700 requires a sufficient electron sink for PSI, which is responsible for flavodiiron proteins (FLV) safely dissipating electrons to O2 in cyanobacteria, green algae, and land plants except for angiosperms during short-pulse light (SP) illumination under which photosynthesis and photorespiration do not occur. This fact implies that O2 usage is essential for P700 oxidation but also raises the question why angiosperms lost FLV. Here, we first found that aquatic photoautotrophs in red plastid lineage, in which no gene for FLV has been found, could keep P700 oxidized during SP illumination alleviating the photo-oxidative damage in PSI even without O2 usage. We comprehensively assessed P700 oxidation during SP illumination in the presence and absence of O2 in cyanobacteria (Cyanophyta), green algae (Chlorophyta), angiosperms (Streptophyta), red algae (Rhodophyta), and secondary algae (Cryptophyta, Haptophyta, and Heterokontophyta). A variety of dependencies of P700 oxidation on O2 among these photoautotrophs clearly suggest that O2 usage and FLV are not universally required to oxidize P700 for protecting PSI against ROS damage. Our results expand the understanding of the diverse strategies taken by oxygenic photoautotrophs to oxidize P700 and mitigate the risks of ROS.
Similar content being viewed by others
Avoid common mistakes on your manuscript.
Introduction
Oxygenic photoautotrophs, including plants and algae, generally develop the same photosynthetic electron transport system composed of photosystems (PS) I and II. These photosystems drive the photo-oxidation/reduction cycles of their reaction center chlorophyll (P700 and P680, respectively) to build NADPH and ATP for the production of sugars from CO2, with H2O as the electron donor. The photosynthetic linear electron flow is performed on the thylakoid membrane from PSII to PSI by the plastoquinone (PQ), cytochrome (Cyt) b6/f, and plastocyanin (or Cyt c6), finally reducing NADP+ to NADPH on the electron acceptor side of PSI. In this process, thylakoid lumen acidification is accompanied with the oxidation of H2O at PSII and the Q-cycle in Cyt b6/f, which functions as the proton motive force of chloroplast ATP synthase. Both NADPH and ATP produced in the photosynthetic electron transport are required afterwards for photosynthetic CO2 assimilation in the Calvin–Benson cycle.
Photo-excitation of the reaction center chlorophyll in PSI (P700) is tuned with the oxidized form of P700 (P700+) for the safe utilization of NADPH in the Calvin–Benson cycle. This happens because the excess photo-excitation energy has the potential to generate reactive oxygen species (ROS) in PSI (Sejima et al. 2014; Shimakawa et al. 2016a). The redox state of P700 is modulated by a variety of molecular mechanisms (i.e., P700 oxidation systems) on the PSI donor- and acceptor-sides. On the donor side, photosynthetic linear electron flow is limited during the oxidation of reduced PQ in Cyt b6/f, if enough electron acceptors are supplied to PSI (Stiehl and Witt 1969; Anderson 1992). Additionally, in response to the limitation of electron acceptors to PSI, the electron transport in Cyt b6/f is modulated by the proton gradient across the thylakoid membrane, i.e., the lumen acidification suppresses the electron transport from PSII into PSI (Nishio and Whitmarsh 1993; Rott et al. 2011). Furthermore, the reduction of the PQ pool may also slow down the Q-cycle and suppress the electron transport in Cyt b6/f (Shaku et al. 2016). These regulatory mechanisms can contribute to P700 oxidation in response to environmental variations, with a sufficient electron sink on the acceptor side of PSI. At the steady state of photosynthesis, the CO2 assimilation in the Calvin–Benson cycle constitute the most electron sink in the photosynthesis. Additionally, photorespiration consumes the excess photo-excitation energy by using O2 and releasing both CO2 and ribulose 1,5-bisphosphate at the steady state of photosynthesis in land plants, including liverworts, ferns, gymnosperms, and angiosperms, but excluding C4 plants (Hanawa et al. 2017). Further, alternative electron flow (AEF) is defined as the molecular mechanism responsible for dissipating excess electrons uncoupled with photosynthesis. In most cases, AEF is mediated with O2 as the final electron acceptor by flavodiiron proteins (FLV), cyanobacterial and plastidial terminal oxidases, and the water–water cycle (Miyake and Asada 1992; Miyake et al. 1998; Helman et al. 2003; Shimakawa et al. 2015; Curien et al. 2016; Ermakova et al. 2016). Hence, photorespiration is an AEF in a wide sense. However, O2-insensitive AEF has also been reported. Cyclic electron flow is one of the commonest AEF, and it works by donating electrons from ferredoxin at the acceptor side of PSI to the PQ pool (Heber and Walker 1992). In addition, H+ and NO3− can function as electron acceptors in the AEF mediated by hydrogenase and nitrate reductase, respectively (Appel et al. 2000; Flores et al. 2005). Thus, these AEF mechanisms can fulfil the prerequisites of P700 oxidation even when photosynthesis and photorespiration do not occur.
Diverse molecular mechanisms of AEF in oxygenic photoautotrophs are responsible for different strategies to oxidize P700. Thus, a reliable method to evaluate P700 oxidation is of prime importance. Previously, Sejima et al. (2014) established a method using the short-pulse light (SP) to investigate the impact of P700 oxidation on the protection of PSI against ROS damage. They found that saturated SP (e.g., 2000–20,000 µmol photons m−2 s−1, 0.3–1 s) being illuminated onto plant leaves instantaneously filled the photosynthetic electron transport with electrons. Therefore, P700 was not oxidized during the SP illumination, unless an AEF uncoupled with photosynthesis rapidly started to function. Moreover, photo-oxidative damage in PSI was also observed after repetitive SP (rSP) illumination (Sejima et al. 2014; Zivcak et al. 2015a). Recently, this method has been broadly applied to a variety of land plants (e.g., liverworts, ferns, gymnosperms, and angiosperms), indicating that land plants except for angiosperms rapidly induce P700 oxidation during the SP illumination in the presence, but not absence, of O2 (Takagi et al. 2017). The genes encoding FLV are conserved in liverworts, mosses, ferns, and gymnosperms (Zhang et al. 2009; Allahverdiyeva et al. 2015; Yamamoto et al. 2016). Additionally, the mutants of the liverwort Marchantia polymorpha and the moss Physcomitrella patens, which are deficient in FLV, showed the inability to oxidize P700 (Gerotto et al. 2016; Shimakawa et al. 2017a). These data suggest that O2-dependent AEF mediated by FLV is a prerequisite for P700 oxidation during the SP illumination in land plants. However, it is still poorly understood why FLV was lost at the genetic level in the evolution to angiosperms.
To expand the understanding of the diverse P700 oxidation systems in oxygenic photoautotrophs, we first investigated the redox state of P700 during the SP illumination in aquatic photoautotrophs categorized into red plastid lineage and compared it with that of green plastid lineage. It is widely considered that the genes for FLV had not been inherited from cyanobacteria to red algae (Allahverdiyeva et al. 2015). Nevertheless, we found that most of the photoautotrophs in red plastid lineage could keep P700 oxidized during the SP illumination as well as cyanobacteria and green algae. Finally, we found that most of algae, different from angiosperms, can oxidize P700 during the SP illumination, which are classified into O2-dependent and -insensitive aquatic photoautotrophs. From the results of our study, we rethink the reason for why angiosperms lost FLV in the evolutionary process.
Materials and methods
Cyanobacteria, algae, and plants
Cyanobacteria and unicellular algae were cultured in a baffled shake flask on a rotary shaker (100 rpm) in ambient air, and under a light (23 °C, 14 h, 100 µmol photons m−2 s−1, white fluorescent lamp) and dark condition (21 °C, 10 h). We used BG-11 medium (Allen 1968) for Synechococcus elongatus PCC 7942, Porphyridium aerugineum (NIES-1959), and Vischeria punctata (NIES-2147); high-salt medium (Sueoka 1960) for Chlamydomonas reinhardtii (137c); ASN-III medium (Rippka et al. 1981) for Porphyridium purpureum (NIES-2138), Chroomonas placoidea (NIES-705), Nannochloropsis oceanica (NIES-2145) and Isochrysis galbana; and artificial seawater medium with 0.31% half-strength Guillard’s ‘F’ solution (Guillard and Ryther 1962; Guillard 1975) for Phaeodactylum tricornutum (UTEX642).
Plants and seaweeds were obtained from Rokko (Ipomoea nil, Nymphaea tetragona, and Magnolia kobus) and Awaji Islands (Ulva pertusa, Codium fragile, Zostera marina, Pyropia yezoensis, Chondrus ocellatus, Chondrus giganteus, Callophyllis japonica, Ecklonia cava, Dictyota dichotoma, Sargassum horneri, and Undaria pinnatifida) in Hyogo, Japan.
Measurement of P700
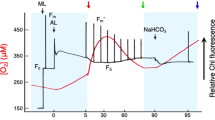
Samples of plants and algae acclimatized for 3–4 h to dark were used for the measurements. The P700+ transmittance was measured using a Dual-PAM-100 (Walz Heinz GmbH, Effeltrich, Germany) at room temperature (25 ± 2 °C) (Klughammer and Schreiber 1994; Schreiber and Klughammer 2008). Reaction cuvettes for gas (3010 DUAL, Walz, Effeltrich, Germany; 1.3 cm−2; for I. nil, M. kobus, and N. tetragona) and liquid phases (ED-101US/MD, Walz, Effeltrich, Germany; 1–2 mL; for the other species) were used. For the liquid phase measurements, plants and algae were washed and measured in a fresh media or natural seawater. Unicellular algae (or cyanobacteria) in cuvettes (20 µg chlorophyll a mL−1) (Grimme and Boardman 1972; Jeffrey and Humphrey 1975; Shimakawa et al. 2017b) were stirred with a magnetic microstirrer, which was temporarily stopped 5 s before the SP illumination. Total oxidizable P700 (i.e., maximum oxidation level of P700) was determined according to the standard method of Schreiber and Klughammer (2008). For the illumination treatment, SPs (2000 or 20,000 µmol photons m−2 s−1 for 1 s using LED with peak emissions at 635 nm) were repeatedly applied every 10 s to the plants and algae. The P700+ kinetics during SP illumination was analyzed 1 min after rSP illumination started. For the calculation of P700 oxidation index, the integration of P700+ during the 1s-SP illumination was estimated from the sum of relative P700+ values every 0.3 ms from Fig. 1, which was then divided by the maximum integrated area (i.e., 1 × 1 s). Finally, the value was shown as the percentage.

Kinetics of oxidized P700 (P700+) in the illumination with a short-pulse light (SP: 2000 µmol photons m−2 s−1, 1 s) in the cyanobacterium Synechococcus elongatus PCC 7942 (a), in the green algae Chlamydomonas reinhardtii (b), Ulva pertusa (c), and Codium fragile (d), in the angiosperms Ipomoea nil (e), Nymphaea tetragona (f), Magnolia kobus (g), and Zostera marina (h), in the red algae Pyropia yezoensis (i), Porphyridium aerugineum (j), Porphyridium purpureum (k), Chondrus ocellatus (l), Chondrus giganteus (m), Callophyllis japonica (n), and Grateloupia lanceolata (o), in the unicellular secondary algae Chroomonas placoidea (p), Isochrysis galbana (q), Nannochloropsis oceanica (r), Vischeria punctata (s), and Phaeodactylum tricornutum (t), and in the brown algae Ecklonia cava (u), Dictyota dichotoma (v), Sargassum horneri (w), and Undaria pinnatifida (x). Different colors (blue, green, red, yellow, and brown) separately show the cyanobacterium, green algae, angiosperms, red algae, unicellular secondary algae, and brown algae. SP was applied on each of all organisms in dark conditions in the presence (dark lines) and absence (light lines) of O2. Relative P700+ amount is normalized by the maximum oxidation level of P700 (i.e., total oxidizable P700) as 1.0. Each bold line and its shaded edge respectively indicate the mean and standard deviation of three measurements
Removal of O2
Eliminating O2 was carefully performed according to the experimental situations. For the measurement of the P700+ kinetics in a liquid phase, an anaerobic condition was prepared by glucose (5 mM), catalase (500 units mL−1, Wako, from bovine liver), and glucose oxidase (10 units mL−1, Wako, from Aspergillus niger). In the presence of these agents, the O2 concentration in the reaction culture rapidly decreases to the anaerobic level within 10 min (Shimakawa et al. 2016a, b). On the other hand, for the gas phase measurement we removed O2 by flushing pure N2 gas to the reaction chamber. In this study, we confirmed the similar effects of eliminating O2 on the P700+ kinetics 5, 10, and 15 min after a start of these procedures in advance for all measurements, and finally obtained the kinetics after the 10 min-procedures. Only in the experiment of Supplemental Fig. S1, we applied the N2 bubbling to the reaction culture to remove O2 also in a liquid phase treatment. After repetitive illumination with SP for 30 min, the reaction culture was bubbled with ambient air for 10 min, and total oxidizable P700 was evaluated again.
Results
Oxidation of P700 in PSI during short-pulse light illumination in aquatic photoautotrophs
To assess the physiological significances and molecular mechanisms of P700 oxidation, we measured the redox state of P700 in PSI during SP illumination (2000 µmol photons m−2 s−1, 1 s) in a variety of aquatic photoautotrophs, including cyanobacteria (Cyanophyta), green algae (Chlorophyta), angiosperms (Streptophyta), red algae (Rhodophyta), and secondary algae (Cryptophyta, Haptophyta, and Heterokontophyta). Cyanobacteria possess FLV and can rapidly induce P700 oxidation in response to an illumination treatment (Helman et al. 2003; Allahverdiyeva et al. 2013). In this study, we confirmed that the cyanobacterium Synechococcus elongatus PCC 7942 kept P700 oxidized during the SP illumination (Fig. 1a), which is consistent with previous results on Synechocystis sp. PCC 6803 (Shimakawa and Miyake 2018). In cyanobacterial cells, where photosynthetic and respiratory electron transport systems share the redox state of the PQ pool and Cyt b6/f, respiratory terminal oxidases (i.e., aa3-type Cyt c oxidase and Cyt bd quinol oxidase) also have an effect on P700 oxidation during a SP illumination (Shimakawa and Miyake 2018). Overall, O2 is essential for P700 oxidation during a SP illumination similar to the P700+ kinetics in the absence of O2 (Fig. 1a).
Next, we investigated the P700+ kinetics during a SP illumination in green algae (Fig. 1b–d). The seaweeds Ulva pertusa and Codium fragile showed a rapid induction of P700 oxidation during the SP illumination, similar to the cyanobacterium S. elongatus PCC 7942. Removing O2 inhibited the oxidation of P700 in both macrophytic green algae (Fig. 1c, d). It has been reported that the unicellular green alga Chlamydomonas reinhardtii presents FLV-mediated AEF with O2 to oxidize P700 (Chaux et al. 2017), which was supported by our results of the P700+ kinetics during the SP illumination (Fig. 1b). From these data, both U. pertusa and C. fragile seem to utilize FLV for P700 oxidation during SP illumination, similar to that by C. reinhardtii.
Responses of the redox state of P700 in PSI to the illumination with SP were also measured in angiosperm leaves (Fig. 1e–h). The aquatic angiosperm Zostera marina (seagrass), similar to a number of angiosperm species previously investigated (Sejima et al. 2014; Zivcak et al. 2015a, b; Takagi et al. 2017), did not show P700 oxidation during the SP illumination independently of O2 presence (Fig. 1h). Further, we confirmed the inability to induce P700 oxidation during SP illumination in the well-known angiosperm Ipomoea nil (Fig. 1e) and also in the primitive Nymphaea tetragona and Magnolia kobus (Fig. 1f, g). These data indicate that angiosperms did not develop a P700 oxidation system using FLV, in any of their habitats.
Red algae are considered to be photosynthetic eukaryotes that originated from an endosymbiont of cyanobacteria with a heterotroph host, but which evolved differently (the so-called red plastid lineage) then the green algae (the green plastid lineage) (Falkowski et al. 2004). In contrast to green algae, genes encoding FLV have never been found in the genomes of red algae (Allahverdiyeva et al. 2015). Hence, red algae are expected to be unable to keep P700 oxidized during SP illumination, similar to that in the angiosperms. Indeed, we obtained P700+ kinetics during SP illumination in accord with the expectation for the macrophytic red alga Pyropia yezoensis (Fig. 1i). In contrast, the unicellular red algae Porphyridium aerugineum and Porphyridium purpureum showed P700 oxidation during the SP illumination. Surprisingly, P700 was kept oxidized in those two Porphyridium species during SP application even when O2 was absent (Fig. 1j, k). Additionally, the macrophytic red algae Chondrus ocellatus, Chondrus giganteus, Callophyllis japonica, and Grateloupia lanceolata also showed O2-insensitive P700 oxidation during SP illumination (Fig. 1l–o). Overall, these red algae developed a different strategy to oxidize P700 for alleviating the risks of ROS production in PSI, as compared to cyanobacteria, green algae, and land plants.
We evaluated the P700+ kinetics during SP illumination in unicellular secondary algae that have red plastids. The cryptomonad (Cryptophyta) Chroomonas placoidea kept a large part of P700 oxidized during SP illumination regardless of O2 presence (Fig. 1p), similar to that in the secondary alga (Haptophyta) Isochrysis galbana (Fig. 1q). These results suggest that the molecular mechanism for the O2-insensitive P700 oxidation has been inherited by these secondary algae with red plastid. Additionally, we measured the redox state of P700 during SP illumination in the two eustigmatophytes (Heterokontophyta) Nannochloropsis oceanica and Vischeria punctata. These unicellular secondary algae showed oxidation of P700 during the SP illumination, but the relationships between P700 oxidation and O2 presence in both species are strikingly different: N. oceanica could rapidly induce P700 oxidation even in the absence of O2 (Fig. 1r), whereas V. punctata could not oxidize P700 without O2, similar to cyanobacteria and green algae (Fig. 1s). Thus, V. punctata strongly relied on O2 to keep P700 oxidized, although the underlying molecular mechanism is still unclear. For the diatom (Heterokontophyta) Phaeodactylum tricornutum, P700 was fully oxidized during SP illumination in both O2 conditions (Fig. 1t). Previously, we quantitatively estimated the electron flux capacity through an O2-dependent electron flow at the steady state of photosynthesis, and we found that P. tricornutum hardly utilized O2 as an alternative electron sink (Shimakawa et al. 2017b). These data suggest that P. tricornutum does not rely on O2 to protect PSI against photo-oxidative damages.
Brown algae (Heterokontophyta) are macrophytic algae with red plastid. Here, we measured the P700+ kinetics during SP illumination in four brown algae species, namely Ecklonia cava, Dictyota dichotoma, Sargassum horneri, and Undaria pinnatifida. We found that P700 was kept oxidized during the SP illumination in all of them (Fig. 1u–x). Additionally, we observed strong relationships between P700 oxidation and O2 presence in these brown algae, except for E. cava, which partially showed P700 oxidation during SP illumination. Therefore, brown algae utilized O2 to oxidize P700, but this O2 importance may vary depending on the species.
P700 oxidation index during the short-pulse light illumination in the presence and absence of O2 in aquatic photoautotrophs
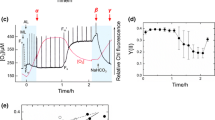
The amount of oxidized P700 during the SP illumination of 1 s in the presence and absence of O2 was calculated from Fig. 1. We defined P700 oxidation index as the ratio (%) of P700+ to the total oxidizable P700 during the SP illumination. All angiosperms used in this study could not keep P700 oxidized during SP illumination, and the P700 oxidation indices were overall low regardless of the O2 presence (Fig. 2). Among algae, the red alga P. yezoensis was the only exception that could be categorized into the same group as angiosperms (Fig. 2). Cyanobacteria and green algae showed high P700 oxidation indices during the SP illumination only in the presence of O2, probably due to FLV (Fig. 2). Conversely, except for P. yezoensis, red algae showed the highest P700 oxidation indices also in O2 absence (Fig. 2). In the secondary algae with the chloroplasts derived from red algae, including unicellular and multicellular, P700 oxidation indices in O2 absence were observed across a wide range (Fig. 2). In this study, we defined two categories: O2-dependent P700 oxidation group, which shows > 50% and < 20% of P700 oxidation indices respectively in the presence and absence of O2; and O2-insensitive P700 oxidation group, which shows > 50% of P700 oxidation indices regardless of the existence of O2 (Fig. 2). Overall, the various algae used in this study can be classified into (1) the O2-use, (2) O2 non-use for keeping P700 oxidized during SP illumination, (3) some intermediates (C. placoidea, I. galbana, and E. cava), and (4) angiosperm-like species (i.e., P. yezoensis).

Relationship between the P700 oxidation indices in the presence and absence of O2. The ratio of the oxidized form to total oxidizable P700 (P700 oxidation index, see Materials and Methods) was calculated during a short-pulse light illumination (SP: 2000 µmol photons m−2 s−1, 1 s). Dashed green and red boxes show O2-dependent and -insensitive P700 oxidation groups. Note that, these results were calculated from the average data in Fig. 1
Impacts of P700 oxidation on the protection of PSI against photo-oxidative damages in aquatic photoautotrophs
Oxidation of P700 suppresses the production of ROS in PSI in cyanobacteria and land plants (Sejima et al. 2014; Shimakawa et al. 2016a, 2017a; Takagi et al. 2017). In this study, we verified the physiological significance of P700 oxidation during the SP illumination in diverse aquatic photoautotrophs. The plants and algae investigated were exposed to a strong SP (20,000 µmol photons m−2 s−1, 1 s) for 30 min between intervals of 10 s. After treatments with repetitive SP illumination (rSP illumination; Sejima et al. 2014), the amount of total oxidizable P700 was spectrophotometrically determined and compared with the initial values before treatment application. The total oxidizable P700 is an indicator of the photochemical activity of PSI (Sejima et al. 2014; Zivcak et al. 2015a; Shimakawa et al. 2016a; Mosebach et al. 2017; Shimakawa and Miyake 2018). Measuring P700 oxidation is also useful for evaluating the photo-oxidative damages in PSI, and it has already been applied in plant leaves and cyanobacteria (Sejima et al. 2014; Zivcak et al. 2015a, b; Shimakawa et al. 2017a; Takagi et al. 2017; Shimakawa and Miyake 2018). We found that rSP illumination caused a decrease in total oxidizable P700 even when weaker SPs were used (e.g., 2000 µmol photons m−2 s−1; Sejima et al. 2014). Contrastingly, all angiosperms and the red alga P. yezoensis showed 60–80% decrease in total oxidizable P700 after rSP illumination, and the other algae kept more than 60% of the total oxidizable P700 values intact (Fig. 3). We further applied rSP illumination to the studied oxygenic photoautotrophs in anaerobic conditions. Suppression of the decrease in total oxidizable P700 in O2 absence suggested that the inactivation of PSI during rSP illumination was caused by ROS (Supplemental Fig. S1). Overall, the consistency of the results on P700 oxidation during SP illumination with the protection of PSI from photo-oxidative damages was strongly recognized in the aquatic photoautotrophs used in this study.

Residual total oxidizable P700 after rSP illumination (20,000 µmol photons m−2 s−1, 1 s, every 10 s, for 30 min) in the cyanobacterium (Synechococcus elongatus PCC 7942), green algae (Chlamydomonas reinhardtii, Ulva pertusa, and Codium fragile), angiosperms (Ipomoea nil, Nymphaea tetragona, Magnolia kobus, and Zostera marina), red algae (Pyropia yezoensis, Porphyridium aerugineum, Porphyridium purpureum, Chondrus ocellatus, Chondrus giganteus, Callophyllis japonica, and Grateloupia lanceolata), unicellular secondary algae (Chroomonas placoidea, Isochrysis galbana, Nannochloropsis oceanica, Vischeria punctata, and Phaeodactylum tricornutum), and brown algae (Ecklonia cava, Dictyota dichotoma, Sargassum horneri, and Undaria pinnatifida). Bars represent mean ± SD (n = 3)
Discussion
In this study, we first investigated the redox state response of P700 to the SP illumination in a variety of aquatic photoautotrophs and land plants. Almost all algae that we studied here kept P700 oxidized during SP illumination (Fig. 1) and saved a large part of PSI photochemically active after exposure to rSP illumination (Fig. 3), which differed from the angiosperms such as N. tetragona and Z. marina. Oxidation of P700 required O2, particularly in green and brown algae. However, most of red algae kept P700 oxidized during SP illumination even in O2 absence (Figs. 1, 2). Overall, these algae commonly oxidized P700 with varying levels of O2 dependence. The seaweeds we used in this study grow near intertidal or subtidal zone in rocky seashore, and are constantly exposed to dynamic light fluctuations ranging from < 0.05 to approximately 10 Hz due to tides and waves (Greene and Gerard 1990; Wing and Patterson 1993). Therefore, these algae might have developed their molecular mechanisms for P700 oxidation in direct response to strong illumination changes in order to thrive in these habitats.
One hypothetical scenario for the evolution and diversification of electron sinks for P700 oxidation is shown in Fig. 4. In this scenario, aquatic photoautotrophs in red plastid lineage have developed unique P700 oxidation systems on the electron acceptor side of the PSI, opposed to the oxygenic photoautotrophs in the green plastid lineage. The genes for FLV are broadly conserved in cyanobacteria, green algae, and basal land plants (i.e., liverworts, mosses, ferns, and gymnosperms), but considered to be lost at the origin of red algae, glaucophytes, or angiosperms (Allahverdiyeva et al. 2015; Yamamoto et al. 2016). Indeed, studies using mutants reported that FLV functions as an electron sink with O2 to oxidize P700 in cyanobacteria (Helman et al. 2003; Allahverdiyeva et al. 2013; Shimakawa et al. 2016a), the green alga C. reinhardtii (Chaux et al. 2017), the liverwort M. polymorpha (Shimakawa et al. 2017a), and the moss P. patens (Gerotto et al. 2016). Recent studies have proposed that the physiological significance of FLV is conserved also in ferns and gymnosperms (Ilík et al. 2017; Noridomi et al. 2017; Takagi et al. 2017). From these facts, the O2-dependent P700 oxidation in U. pertusa and C. fragile is likely to be driven by FLV (Fig. 1c, d). Moreover, genes for FLV have not been found in the genomes of red algae (Allahverdiyeva et al. 2015), implying that red algae did not inherit FLV from cyanobacteria at an early symbiotic event. Hence, red algae and secondary algae with red plastid may show O2-insensitive or O2-dependent P700 oxidation strategies, which might have been driven by unique P700 oxidation mechanisms (Fig. 1i–x). Only in the red alga P. yezoensis, we failed to observe a P700 oxidation during SP illumination (Fig. 1l). Pyropia yezoensis (Bangiaceae) is classified into a different group when compared to Porphyridium (Porphyridiophyceae), Chondrus, Callophyllis, and Grateloupia (Florideophyceae) in the red algal phylogeny (Freshwater et al. 1994), suggesting that the O2-insensitive P700 oxidation system in the former species might have evolved during the diversification of red algae. In addition to FLV, photorespiration may also be considered an electron sink with O2 on the acceptor side of PSI. Genes for the photorespiratory C2 cycle are recognized in all oxygenic photoautotrophs, and they play a critical role on plants and algae growth (Kozaki and Takeba 1996; Eisenhut et al. 2008; Zelitch et al. 2009; Hagemann et al. 2013; Rademacher et al. 2016). Nevertheless, the estimated electron sink capacity of photorespiration is expected to be smaller for cyanobacteria and eukaryotic algae (Bidwell and McLachlan 1985; Hayashi et al. 2014; Shimakawa et al. 2016b, 2017b), compared to the oxygenic photoautotrophs, given that these experience more adequate O2 conditions (Hanawa et al. 2017). Overall, all algae derived from red algae (red plastid lineage) are likely to utilize neither FLV nor photorespiration for P700 oxidation (Fig. 4).

Hypothetical illustrations of the evolutionary history of the O2-usage strategy for P700 oxidation in oxygenic photoautotrophs. a The evolutionary paths of green and red plastid lineages, with three endosymbiosis events shown as blue lines. The estimated evolution with and without the endosymbiosis is indicated by solid and dashed arrows, respectively. b The change of electron-sink mechanisms in millions of years for PSI during the evolutionary history of oxygenic photoautotrophs
In this study, we found for the first time that P700 was kept oxidized during SP illumination even in the absence of O2 in some species of red algae and secondary algae with red plastid (Fig. 1). Our results suggest that these algae utilize an O2-insensitive AEF on the acceptor side of PSI as an appropriate electron sink, which is required for P700 oxidation. Probably, the Calvin–Benson cycle is activated to prepare a sufficient electron sink in response to the SP illumination even in the dark in these algae. Additionally, the P700 oxidation systems on the electron donor side of PSI are probably dominant for P700 oxidation in these algae. Photosynthetic linear electron transport between PSII and PSI is basically limited in Cyt b6/f (Anderson 1992), and it can be further suppressed by regulatory mechanisms (Kramer et al. 2003; Shaku et al. 2016). Suppression of the electron transport in Cyt b6/f by the thylakoid lumen acidification is the best understood regulatory mechanism, also known as the photosynthetic control (Schöttler et al. 2015). In plant leaves, the lumen acidification is promoted by proton conductance modulations of chloroplast ATP synthase (Rott et al. 2011) and cyclic electron flow (Nandha et al. 2007) in response to situations of limitation on the electron acceptor side of PSI. In the diatom P. tricornutum, which showed O2-insensitive P700 oxidation during SP illumination (Fig. 1t), NADP+ and ATP can be modulated by the interaction with mitochondria (Bailleul et al. 2015; Flori et al. 2017), possibly contributing to the oxidation of P700 during SP illumination from the electron donor and acceptor sides of the PSI. The O2-insensitive mechanisms for P700 oxidation in some species of red algae and secondary algae with red plastid should be investigated in future surveys.
In addition to cyanobacteria and green algae harboring FLV, we also observed the O2-dependent P700 oxidation during SP illumination in the unicellular secondary alga V. punctata and in four brown algae species (Fig. 1k, m–p). Given that red algae lack FLV (Allahverdiyeva et al. 2015), these algae are likely to safely dissipate electrons to O2 through alternative pathways other than FLV such as plastidial terminal oxidase (i.e., chlororespiration; Krieger-Liszkay and Feilke 2016) and the Mehler reaction (i.e., water–water cycle; Mehler 1951; Miyake and Asada 1992). Conversely, the dinoflagellate Symbiodinium species exceptionally have analogous genes for FLV, and it was shown that they present an O2-dependent AEF (Roberty et al. 2014). The evolutionary origin of FLV genes may be difficult to be fully explained along with the conventional phylogeny of oxygenic photoautotrophs (Falkowski et al. 2004). Unfortunately, we could not exclude the possibility that FLV functions as a P700 oxidation system in V. punctata, and in the brown and green algae.
Here, we rethink the reason for why angiosperms lost FLV during its evolutionary history. From the evaluation of P700 oxidation during SP illumination, the red algae from Porphyridium, Chondrus, Callophyllis, and Grateloupia could keep oxidized P700 without O2 and FLV (Fig. 1j–o). This indicates that FLV is not always essential for P700 oxidation during SP illumination. Moreover, FLV was lost upon the origin of red algae earlier than for angiosperms during the evolutionary history of oxygenic photoautotrophs (Fig. 4). Although its activity should be strictly modulated in response to environmental variations, FLV is likely to have the potential to deprive the acceptor side of PSI of electrons for photosynthetic CO2 assimilation. Additionally, FLV has a diiron site that demands iron and is highly costly (Vicente et al. 2008). Further, FLV has two flavin modules in one protein, which have the potential to react with O2 and produce superoxide anion radical (O2−) (Massey et al. 1969; Miyake et al. 1998; Foyer and Noctor 2000; Mubarakshina et al. 2010; Schmitt et al. 2015). Therefore, the lack of FLV in angiosperms may be reasonable for their success to thrive in terrestrial habitats. Finally, during the evolutionary history of land plants, some angiosperms have developed CO2-concentrating mechanisms (CCM) to stop utilizing O2 in photorespiration in addition to FLV. This is interestingly similar to the strategies used to prepare electron sinks in aquatic photoautotrophs of the red plastid lineage (Fig. 4). Each oxygenic photoautotroph would develop the survival strategies suit to their lifestyle and/or habitat.
References
Allahverdiyeva Y, Mustila H, Ermakova M, Bersanini L, Richaud P, Ajlani G, Battchikova N, Cournac L, Aro EM (2013) Flavodiiron proteins Flv1 and Flv3 enable cyanobacterial growth and photosynthesis under fluctuating light. Proc Natl Acad Sci USA 110:4111–4116
Allahverdiyeva Y, Isojarvi J, Zhang P, Aro EM (2015) Cyanobacterial oxygenic photosynthesis is protected by flavodiiron proteins. Life 5:716–743
Allen MM (1968) Simple conditions for growth of unicellular blue-green algae on plates1, 2. J Phycol 4:1–4
Anderson JM (1992) Cytochrome b 6 f complex: dynamic molecular organization, function and acclimation. Photosynth Res 34:341–357
Appel J, Phunpruch S, Steinmuller K, Schulz R (2000) The bidirectional hydrogenase of Synechocystis sp. PCC 6803 works as an electron valve during photosynthesis. Arch Microbiol 173:333–338
Bailleul B, Berne N, Murik O, Petroutsos D, Prihoda J, Tanaka A, Villanova V, Bligny R, Flori S, Falconet D, Krieger-Liszkay A, Santabarbara S, Rappaport F, Joliot P, Tirichine L, Falkowski PG, Cardol P, Bowler C, Finazzi G (2015) Energetic coupling between plastids and mitochondria drives CO2 assimilation in diatoms. Nature 524:366–369
Bidwell RGS, McLachlan J (1985) Carbon nutrition of seaweeds: photosynthesis, photorespiration and respiration. J Exp Mar Biol Ecol 86:15–46
Chaux F, Burlacot A, Mekhalfi M, Auroy P, Blangy S, Richaud P, Peltier G (2017) Flavodiiron proteins promote fast and transient O2 photoreduction in Chlamydomonas. Plant Physiol 174:1825–1836
Curien G, Flori S, Villanova V, Magneschi L, Giustini C, Forti G, Matringe M, Petroutsos D, Kuntz M, Finazzi G (2016) The water to water cycles in microalgae. Plant Cell Physiol 57:1354–1363
Eisenhut M, Ruth W, Haimovich M, Bauwe H, Kaplan A, Hagemann M (2008) The photorespiratory glycolate metabolism is essential for cyanobacteria and might have been conveyed endosymbiontically to plants. Proc Natl Acad Sci USA 105:17199–17204
Ermakova M, Huokko T, Richaud P, Bersanini L, Howe CJ, Lea-Smith DJ, Peltier G, Allahverdiyeva Y (2016) Distinguishing the roles of thylakoid respiratory terminal oxidases in the cyanobacterium Synechocystis sp. PCC 6803. Plant Physiol 171:1307–1319
Falkowski PG, Katz ME, Knoll AH, Quigg A, Raven JA, Schofield O, Taylor FJR (2004) The evolution of modern eukaryotic phytoplankton. Science 305:354–360
Flores E, Frías JE, Rubio LM, Herrero A (2005) Photosynthetic nitrate assimilation in cyanobacteria. Photosynth Res 83:117–133
Flori S, Jouneau P-H, Bailleul B, Gallet B, Estrozi LF, Moriscot C, Bastien O, Eicke S, Schober A, Bártulos CR, Maréchal E, Kroth PG, Petroutsos D, Zeeman S, Breyton C, Schoehn G, Falconet D, Finazzi G (2017) Plastid thylakoid architecture optimizes photosynthesis in diatoms. Nat Commun 8:15885
Foyer CH, Noctor G (2000) Oxygen processing in photosynthesis: regulation and signalling. New Phytol 146:359–388
Freshwater DW, Fredericq S, Butler BS, Hommersand MH, Chase MW (1994) A gene phylogeny of the red algae (Rhodophyta) based on plastid rbcL. Proc Natl Acad Sci USA 91:7281–7285
Gerotto C, Alboresi A, Meneghesso A, Jokel M, Suorsa M, Aro E-M, Morosinotto T (2016) Flavodiiron proteins act as safety valve for electrons in Physcomitrella patens. Proc Natl Acad Sci USA 113:12322–12327
Greene RM, Gerard VA (1990) Effects of high-frequency light fluctuations on growth and photoacclimation of the red alga Chondrus crispus. Mar Biol 105:337–344
Grimme LH, Boardman NK (1972) Photochemical activities of a particle fraction P1 obtained from the green alga Chlorella fusca. Biochem Biophys Res Commun 49:1617–1623
Guillard RRL (1975) Culture of phytoplankton for feeding marine invertebrates. In: Smith WL, Chanley MH (eds) Culture of marine invertebrate animals. Springer US, Boston, pp 29–60
Guillard RRL, Ryther JH (1962) Studies of marine planktonic diatoms: I. Cyclotella nana hustedt, and Detonula confervacea (cleve) gran. Can J Microbiol 8:229–239
Hagemann M, Fernie AR, Espie GS, Kern R, Eisenhut M, Reumann S, Bauwe H, Weber APM (2013) Evolution of the biochemistry of the photorespiratory C2 cycle. Plant Biol 15:639–647
Hanawa H, Ishizaki K, Nohira K, Takagi D, Shimakawa G, Sejima T, Shaku K, Makino A, Miyake C (2017) Land plants drive photorespiration as higher electron-sink: comparative study of post-illumination transient O2-uptake rates from liverworts to angiosperms through ferns and gymnosperms. Physiol Plant 161:138–149
Hayashi R, Shimakawa G, Shaku K, Shimizu S, Akimoto S, Yamamoto H, Amako K, Sugimoto T, Tamoi M, Makino A, Miyake C (2014) O2-dependent large electron flow functioned as an electron sink, replacing the steady-state electron flux in photosynthesis in the cyanobacterium Synechocystis sp. PCC 6803, but not in the cyanobacterium Synechococcus sp. PCC 7942. Biosci Biotechnol Biochem 78:384–393
Heber U, Walker D (1992) Concerning a dual function of coupled cyclic electron transport in leaves. Plant Physiol 100:1621–1626
Helman Y, Tchernov D, Reinhold L, Shibata M, Ogawa T, Schwarz R, Ohad I, Kaplan A (2003) Genes encoding A-type flavoproteins are essential for photoreduction of O2 in cyanobacteria. Curr Biol 13:230–235
Ilík P, Pavlovič A, Kouřil R, Alboresi A, Morosinotto T, Allahverdiyeva Y, Aro E-M, Yamamoto H, Shikanai T (2017) Alternative electron transport mediated by flavodiiron proteins is operational in organisms from cyanobacteria up to gymnosperms. New Phytol 214:967–972
Jeffrey SW, Humphrey GF (1975) New spectrophotometric equations for determining chlorophylls a, b, c 1 and c 2 in higher plants, algae and natural phytoplankton. Biochem Physiol Pflanz 167:191–194
Klughammer C, Schreiber U (1994) An improved method, using saturating light pulses, for the determination of photosystem I quantum yield via P700+-absorbance changes at 830 nm. Planta 192:261–268
Kozaki A, Takeba G (1996) Photorespiration protects C3 plants from photooxidation. Nature 384:557–560
Kramer DM, Cruz JA, Kanazawa A (2003) Balancing the central roles of the thylakoid proton gradient. Trends Plant Sci 8:27–32
Krieger-Liszkay A, Feilke K (2016) The dual role of the plastid terminal oxidase PTOX: Between a protective and a pro-oxidant function. Front Plant Sci 6:1147
Massey V, Strickland S, Mayhew SG, Howell LG, Engel PC, Matthews RG, Schuman M, Sullivan PA (1969) The production of superoxide anion radicals in the reaction of reduced flavins and flavoproteins with molecular oxygen. Biochem Biophys Res Commun 36:891–897
Mehler AH (1951) Studies on reactions of illuminated chloroplasts. Arch Biochem Biophys 33:65–77
Miyake C, Asada K (1992) Thylakoid-bound ascorbate peroxidase in spinach chloroplasts and photoreduction of its primary oxidation product monodehydroascorbate radicals in thylakoids. Plant Cell Physiol 33:541–553
Miyake C, Schreiber U, Hormann H, Sano S, Kozi A (1998) The FAD-enzyme monodehydroascorbate radical reductase mediates photoproduction of superoxide radicals in spinach thylakoid membranes. Plant Cell Physiol 39:821–829
Mosebach L, Heilmann C, Mutoh R, Gäbelein P, Steinbeck J, Happe T, Ikegami T, Hanke G, Kurisu G, Hippler M (2017) Association of Ferredoxin: NADP+ oxidoreductase with the photosynthetic apparatus modulates electron transfer in Chlamydomonas reinhardtii. Photosynth Res 134:291–306
Mubarakshina MM, Ivanov BN, Naydov IA, Hillier W, Badger MR, Krieger-Liszkay A (2010) Production and diffusion of chloroplastic H2O2 and its implication to signalling. J Exp Bot 61:3577–3587
Nandha B, Finazzi G, Joliot P, Hald S, Johnson GN (2007) The role of PGR5 in the redox poising of photosynthetic electron transport. Biochim Biophys Acta Bioenerg 1767:1252–1259
Nishio JN, Whitmarsh J (1993) Dissipation of the proton electrochemical potential in intact chloroplasts II. The pH gradient monitored by cytochrome f reduction kinetics. Plant Physiol 101:89–96
Noridomi M, Nakamura S, Tsuyama M, Futamura N, Vladkova R (2017) Opposite domination of cyclic and pseudocyclic electron flows in short-illuminated dark-adapted leaves of angiosperms and gymnosperms. Photosynth Res 134:149–164
Rademacher N, Kern R, Fujiwara T, Mettler-Altmann T, Miyagishima S, Hagemann M, Eisenhut M, Weber APM (2016) Photorespiratory glycolate oxidase is essential for the survival of the red alga Cyanidioschyzon merolae under ambient CO2 conditions. J Exp Bot 67:3165–3175
Rippka R, Waterbury JB, Stanier RY (1981) Isolation and purification of cyanobacteria: Some general principles. In: Starr MP, Stolp H, Trüper HG, Balows A, Schlegel HG (eds) The prokaryotes: a handbook on habitats, isolation, and identification of bacteria. Springer Berlin Heidelberg, Berlin, pp 212–220
Roberty S, Bailleul B, Berne N, Franck F, Cardol P (2014) PSI Mehler reaction is the main alternative photosynthetic electron pathway in Symbiodinium sp., symbiotic dinoflagellates of cnidarians. New Phytol 204:81–91
Rott M, Martins NF, Thiele W, Lein W, Bock R, Kramer DM, Schöttler MA (2011) ATP synthase repression in tobacco restricts photosynthetic electron transport, CO2 assimilation, and plant growth by overacidification of the thylakoid lumen. Plant Cell 23:304–321
Schmitt F, Kreslavski VD, Zharmukhamedov SK, Friedrich T, Renger G, Los DA, Kuznetsov VV, Allakhverdiev SI (2015) The multiple roles of various reactive oxygen species (ROS) in photosynthetic organisms. In Photosynthesis, Allakhverdiev SI (ed). https://doi.org/10.1002/9781119084150.ch1
Schöttler MA, Tóth SZ, Boulouis A, Kahlau S (2015) Photosynthetic complex stoichiometry dynamics in higher plants: biogenesis, function, and turnover of ATP synthase and the cytochrome b 6 f complex. J Exp Bot 66:2373–2400
Schreiber U, Klughammer C (2008) Saturation pulse method for assessment of energy conversion in PSI. PAM Appl Notes 1:11–14
Sejima T, Takagi D, Fukayama H, Makino A, Miyake C (2014) Repetitive short-pulse light mainly inactivates photosystem I in sunflower leaves. Plant Cell Physiol 55:1184–1193
Shaku K, Shimakawa G, Hashiguchi M, Miyake C (2016) Reduction-induced suppression of electron flow (RISE) in the photosynthetic electron transport system of Synechococcus elongatus PCC 7942. Plant Cell Physiol 57:1443–1453
Shimakawa G, Miyake C (2018) Respiratory terminal oxidases alleviate photo-oxidative damage in photosystem I during repetitive short-pulse illumination in Synechocystis sp. PCC 6803. Photosynth Res. https://doi.org/10.1007/s11120-018-0495-y
Shimakawa G, Shaku K, Nishi A, Hayashi R, Yamamoto H, Sakamoto K, Makino A, Miyake C (2015) FLAVODIIRON2 and FLAVODIIRON4 proteins mediate an oxygen-dependent alternative electron flow in Synechocystis sp. PCC 6803 under CO2-limited conditions. Plant Physiol 167:472–480
Shimakawa G, Shaku K, Miyake C (2016a) Oxidation of P700 in photosystem I is essential for the growth of cyanobacteria. Plant Physiol 172:1443–1450
Shimakawa G, Akimoto S, Ueno Y, Wada A, Shaku K, Takahashi Y, Miyake C (2016b) Diversity in photosynthetic electron transport under [CO2]-limitation: the cyanobacterium Synechococcus sp. PCC 7002 and green alga Chlamydomonas reinhardtii drive an O2-dependent alternative electron flow and non-photochemical quenching of chlorophyll fluorescence during CO2-limited photosynthesis. Photosynth Res 130:293–305
Shimakawa G, Ishizaki K, Tsukamoto S, Tanaka M, Sejima T, Miyake C (2017a) The liverwort, Marchantia, drives alternative electron flow using a flavodiiron protein to protect PSI. Plant Physiol 173:1636–1647
Shimakawa G, Matsuda Y, Nakajima K, Tamoi M, Shigeoka S, Miyake C (2017b) Diverse strategies of O2 usage for preventing photo-oxidative damage under CO2 limitation during algal photosynthesis. Sci Rep 7:41022
Stiehl HH, Witt HT (1969) Quantitative treatment of the function of plastoquinone in photosynthesis. Zeitschrift für Naturforschung B 24:1588
Sueoka N (1960) Mitotic replication of deoxyribonucleic acid in Chlamydomonas reinhardi. Proc Natl Acad Sci USA 46:83–91
Takagi D, Ishizaki K, Hanawa H, Mabuchi T, Shimakawa G, Yamamoto H, Miyake C (2017) Diversity of strategies for escaping reactive oxygen species production within photosystem I among land plants: P700 oxidation system is prerequisite for alleviating photoinhibition in photosystem I. Physiol Plant 161:56–74
Vicente JB, Carrondo MA, Teixeira M, Frazao C (2008) Structural studies on flavodiiron proteins. Methods Enzymol 437:3–19
Wing SR, Patterson MR (1993) Effects of wave-induced lightflecks in the intertidal zone on photosynthesis in the macroalgae Postelsia palmaeformis and Hedophyllum sessile (Phaeophyceae). Mar Biol 116:519–525
Yamamoto H, Takahashi S, Badger MR, Shikanai T (2016) Artificial remodelling of alternative electron flow by flavodiiron proteins in Arabidopsis. Nat Plants 2:16012
Zelitch I, Schultes NP, Peterson RB, Brown P, Brutnell TP (2009) High glycolate oxidase activity is required for survival of maize in normal air. Plant Physiol 149:195–204
Zhang P, Allahverdiyeva Y, Eisenhut M, Aro EM (2009) Flavodiiron proteins in oxygenic photosynthetic organisms: photoprotection of photosystem II by Flv2 and Flv4 in Synechocystis sp. PCC 6803. PLoS ONE 4:e5331
Zivcak M, Brestic M, Kunderlikova K, Sytar O, Allakhverdiev SI (2015a) Repetitive light pulse-induced photoinhibition of photosystem I severely affects CO2 assimilation and photoprotection in wheat leaves. Photosynth Res 126:449–463
Zivcak M, Brestic M, Kunderlikova K, Olsovska K, Allakhverdiev SI (2015b) Effect of photosystem I inactivation on chlorophyll a fluorescence induction in wheat leaves: does activity of photosystem I play any role in OJIP rise? J Photochem Photobiol B: Biol 152:318–324
Acknowledgements
The authors thank Prof. Yuichiro Takahashi (Okayama University) for supplying the culture of Chlamydomonas reinhardtii and Editage (http://www.editage.jp) for providing English corrections.
Funding
This work was supported by the Japan Society for the Promotion of Science (JSPS; Grant No. 26450079 to C.M.) and the Core Research for Evolutional Science and Technology (CREST) division of the Japan Science and Technology Agency (Grant No. AL65D21010 to C.M.). G.S. was supported as a JSPS research fellow (Grant No. 16J03443).
Author information
Authors and Affiliations
Contributions
CM conceived the original screening and research plans; CM supervised the experiments; GS performed most of the experiments; AM, KN, YM, and AW provided technical assistance to GS; CM and GS designed the experiments and analyzed the data; CM and GS conceived the project and wrote the manuscript.
Corresponding author
Ethics declarations
Conflict of interest
The authors have no conflict of interest to declare.
Electronic supplementary material
Below is the link to the electronic supplementary material.
Supplemental Fig. S1
. Residual total oxidizable P700 after rSP illumination (20,000 μmol photons m−2 s−1, 1 s, every 10 s, for 30 min) in N2 gas presence in the cyanobacterium (Synechococcus elongatus PCC 7942), in the green algae (Chlamydomonas reinhardtii, Ulva pertusa and Codium fragile), in the angiosperms (Ipomoea nil, Nymphaea tetragona, Magnolia kobus and Zostera marina), in the red algae (Pyropia yezoensis, Porphyridium aerugineum, Porphyridium purpureum, Chondrus ocellatus, Chondrus giganteus, Callophyllis japonica and Grateloupia lanceolata), in the unicellular secondary algae (Chroomonas placoidea, Isochrysis galbana, Nannochloropsis oceanica, Vischeria punctata and Phaeodactylum tricornutum) and in the brown algae (Ecklonia cava, Dictyota dichotoma, Sargassum horneri and Undaria pinnatifida). Bars represent mean ± SD (n = 3). (PDF 224 KB)
Rights and permissions
About this article

Cite this article
Shimakawa, G., Murakami, A., Niwa, K. et al. Comparative analysis of strategies to prepare electron sinks in aquatic photoautotrophs. Photosynth Res 139, 401–411 (2019). https://doi.org/10.1007/s11120-018-0522-z
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11120-018-0522-z