
Abstract
The green alga Chlamydomonas (C.) reinhardtii is a model organism for photosynthesis research. State transitions regulate redistribution of excitation energy between photosystem I (PS I) and photosystem II (PS II) to provide balanced photosynthesis. Chlorophyll (Chl) a fluorescence induction (the so-called OJIPSMT transient) is a signature of several photosynthetic reactions. Here, we show that the slow (seconds to minutes) S to M fluorescence rise is reduced or absent in the stt7 mutant (which is locked in state 1) in C. reinhardtii. This suggests that the SM rise in wild type C. reinhardtii may be due to state 2 (low fluorescence state; larger antenna in PS I) to state 1 (high fluorescence state; larger antenna in PS II) transition, and thus, it can be used as an efficient and quick method to monitor state transitions in algae, as has already been shown in cyanobacteria (Papageorgiou et al. 1999, 2007; Kaňa et al. 2012). We also discuss our results on the effects of (1) 3-(3,4-dichlorophenyl)-1,4-dimethyl urea, an inhibitor of electron transport; (2) n-propyl gallate, an inhibitor of alternative oxidase (AOX) in mitochondria and of plastid terminal oxidase in chloroplasts; (3) salicylhydroxamic acid, an inhibitor of AOX in mitochondria; and (4) carbonyl cyanide p-trifluoromethoxyphenylhydrazone, an uncoupler of phosphorylation, which dissipates proton gradient across membranes. Based on the data presented in this paper, we conclude that the slow PSMT fluorescence transient in C. reinhardtii is due to the superimposition of, at least, two phenomena: qE dependent non-photochemical quenching of the excited state of Chl, and state transitions.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Chlorophyll (Chl) a fluorescence transient measurement is a sensitive and non-invasive tool to investigate various processes of photosynthetic apparatus in vivo (see e.g., Papageorgiou et al. 2007; Kalaji et al. 2012a, 2014; Stirbet et al. 2014). Fast (microseconds to a second) fluorescence transient has been used to monitor primary events in photosynthesis, including quantum yield of photosynthesis and electron transport both in photosystem (PS) II and I (Maxwell and Johnson 2000; Baker 2008; see also chapters in Govindjee and Fork 1986, and in Papageorgiou and Govindjee 2004; Schreiber 2004; Strasser et al. 2004). It has been used to monitor effects of various abiotic and biotic stresses on the photosynthetic machinery (see e.g., Lichtenthaler 1988; DeEll and Toivnen 2003; Suggett et al. 2010; Kalaji et al. 2012b, Zivcak et al. 2014). Further, Chl a fluorescence can be used to monitor the slow (seconds to minutes) fluorescence transient in algae and plants to evaluate regulatory mechanisms such as “state changes”, as has already been done in cyanobacteria (Papageorgiou et al. 1999, 2007; Kaňa et al. 2012). In this study, we have examined these changes in the green alga Chlamydomonas (C.) reinhardtii, during the slow (minute range) S to M rise in Chl a fluorescence induction. C. reinhardtii is a model organism for research on both photosynthesis and respiration (see chapters in Rochaix et al. (1998); and Stern et al. (2008) for a Chlamydomonas source book).
In higher plants, algae, and cyanobacteria, PS I and PS II operate in series to generate reduced nicotinamide adenine dinucleotide phosphate (NADPH) and ATP, which are then used for CO2 fixation, by the Calvin–Benson cycle, to form sugars and starch (see a general scheme in Fig. 1). PS II oxidizes water and reduces plastoquinone (PQ), whereas PS I oxidizes plastocyanin (PC) and reduces ferredoxin (Fd). During electron transfer from PS II to PS I, a cytochrome (Cyt) b 6 f complex serves to oxidize plastoquinol (PQH2) and to reduce plastocyanin. In addition to this linear electron flow (LEF), there is also a cyclic electron flow (CEF) around PS I, where electrons at the end of PS I, e.g., on reduced Fd flow back to P700 via the Cyt b 6 f complex (see e.g. Munekage et al. 2004). Both non-cyclic as well as cyclic electron transfer lead to ATP synthesis. The proton-motive force (pmf) (made up of a pH gradient, ΔpH, and a membrane potential, ΔΨ), which is generated during electron flow, is used by ATP synthase to produce ATP (see e.g., Blankenship 2014).

A schematic representation of the interactions between electron transfer flows and carbon assimilation in the chloroplast of C. reinhardtii and metabolic reactions driven by mitochondria. Chemicals used in this study for different treatments of the algal cells are shown close to the processes affected by them. See the main text for explanation. AOX, alternative oxidase; CEF, cyclic electron flow; Cyt, cytochrome; DCMU, 3-(3,4-dichlorophenyl)-1,4-dimethyl urea; FCCP, carbonyl cyanide p-trifluoromethoxyphenylhydrazone; LEF, linear electron flow; NADP, nicotinamide adenine dinucleotide phosphate; NDA2, Type II NADPH dehydrogenase; PC, plastocyanin; PG, n-propyl gallate; PQ, PQH 2 , plastoquinone, plastoquinol; pmf, proton-motive force; PTOX, plastid (or plastoquinol) terminal oxidase; SHAM, salicylhydroxamic acid. A few caveats are (1) DCMU is known to inhibit reduction of PQ to PQH2, but this happens not in the membrane, but inside PS II, at the QB site (see text); (2) proton uptake (nH+) from the stroma matrix between “NDA2” and “PTOX” and its release in the lumen are related to “chlororespiration”; and (3) CEF includes electron flow from NADPH to NDA2. The scheme shown here was modified from that by Alric (2010)
In this paper, we have used Chl a fluorescence induction to monitor several photosynthetic events. Chl a fluorescence induction, measured under saturating light, shows a fast multiphasic rise in µs to second (s) range, the O–J–I–P transient (see e.g., Strasser et al. 1995; Haldimann and Strasser 1999; and reviews by Stirbet and Govindjee 2011, 2012; Schansker et al. 2014; Stirbet et al. 2014), and a slow, in s to minute range, the SMT transient (Papageorgiou 1968; Papageorgiou and Govindjee 1968a, b, 2011). In dark-adapted photosynthetic samples, Q A, an electron acceptor of PS II, is in the oxidized state at the O level (the initial, minimum fluorescence). The O–J fluorescence rise (J being an inflection between O and the peak P) is a photochemical phase related to the reduction of Q A to Q −A , whereas J–I (I being a second inflection between O and P) and I–P are thermal phases involved in reduction of the PQ pool (see e.g., Tóth et al. 2007; Stirbet and Govindjee 2011, 2012) as well as that of the electron acceptor side of PS I (Munday and Govindjee 1969; Schansker et al. 2006). At the “P” level, under saturating light, all the electron carriers between the PS II reaction center and NADP are in the reduced state. In short, there is a “traffic jam” of electrons at the “P” level. In the slow fluorescence transient, the P to S decline in green algae and higher plants may reflect ΔpH changes that induce non-photochemical quenching of Chl a fluorescence (see chapters in Demmig-Adams et al. 2014), and the SMT phase reflects several processes including state transitions (Papageorgiou and Govindjee 2011; Kaňa et al. 2012). We, however, note that the PS decay is too fast to involve xanthophyll cycle, which is a much slower process (see chapters in Demmig-Adams et al. 2014). For a possible relation of conformational change to the OJIPS transient, see Schanker et al. (2011, 2014).
For optimal photosynthesis, the amount of excitation energy absorbed by the two photosystems must be balanced in natural environmental conditions where the quality and quantity of light undergo fluctuations (Allen et al. 1981; Mohanty et al. 2012; Rochaix 2014). State transitions (Murata 1969; Bonaventura and Myers 1969; Allen and Mullineaux 2004; Forti and Caldiroli 2005) are known to balance absorbed energy between the two photosystems by mobilizing specific light-harvesting complex (LHC) II proteins (see e.g., Takahashi et al. 2006; Minagawa 2013; for evolutionary aspects, see Peers et al. 2009). Ünlü et al. (2014) have found important differences between state transitions in plants and algae (also see Rochaix 2014). In plants, 10–15 % of LHC II shuttle between photosystems, whereas in C. reinhardtii, 70–80 % of LHC II seems to be somehow involved in state transitions (Delosme et al. 1996). However, only ~10 to 20 % of these are actually incorporated in PS I antenna, whereas the rest form an array that quench Chl a excited state (Iwai et al. 2010; Nagy et al. 2014).
Molecular events during state transitions have been described as follows. Preferential excitation of PS II leads to excess amount of reduced PQ (i.e., PQH2) in the PQ pool, which when bound to the Qo site of the Cyt b 6 f, triggers activation of Stt7 kinase in C. reinhardtii (Zito et al. 1999), or of STN7 kinase in higher plants (Depege et al. 2003; Bellafiore et al. 2005). This then results in the following chain of events: phosphorylation of LHC II (Bennett et al. 1980), unbinding of phosphorylated LHC II from PS II, and its migration and attachment to the PS I–LHC I complex (Andersson et al. 1982). This increases the antenna size of PS I at the expense of the antenna size of PS II. This is referred to as state 2 since it was initiated by excess light in PS II; state 2 has a lower fluorescence yield since PS I fluorescence yield is lower than that of PS II (see a review by Papageorgiou and Govindjee 2011). On the other hand, preferential excitation of PS I leads to an increase in the amount of oxidized PQ pool, which induces dephosphorylation of LHC II by PPH1/TAP38 phosphatase (Pribil et al. 2010; Shapiguzov et al. 2010). Dephosphorylated LHC II dissociates from PS I–LHC I supercomplex, and reassociates with PS II supercomplex. The result is the formation of state 1, which has a higher fluorescence yield since PS II fluorescence yield is higher than that of PS I. Further, abiotic stress conditions, such as high temperature and anaerobiosis, also trigger state transitions (Sane et al. 1984; Mohanty et al. 2002; Nellaepalli et al. 2011, 2012; Marutani et al. 2014). In C. reinhardtii, cyclic electron flow, around PS I, has also been associated with state transitions (Hemschemeier and Happe 2011).
We note that cyanobacteria display a prominent SM rise, with the M level much higher than the P level, which is not blocked by DCMU (i.e., under conditions where PS I keeps the PQ pool in the oxidized state; Papageorgiou and Govindjee 1968b). Because of this, the SM fluorescence rise was attributed to a state 2 to state 1 transition (see e.g., Papageorgiou et al. 2007), a concept that was further confirmed by the relative contributions of PS II and PS I in the 77 K fluorescence spectra of cyanobacteria, and by cells locked in state 2 (Stamatakis et al. 2007). In addition, Kaňa et al. (2012) demonstrated, using a RpaC − mutant of Synechocystis PCC6803, that the SM fluorescence rise was due to state 2 to state 1 change, since this mutant (which is locked in state 1) and the wild type, when kept in state 2, did not have the SM rise. We emphasize here that cyanobacteria are usually in state 2 in darkness; in addition, they have a high PS I-to-PS II ratio, which provides an effective means to induce state 2 to state 1 transition upon illumination.
Here, we have used both the wild type and its stt7 mutant of the green alga C. reinhardtii (Depege et al. 2003) to investigate the possible reasons for the S to M fluorescence rise. Our results, obtained using a series of Chl a fluorescence induction measurements on C. reinhardtii cells, demonstrate that state 2 to state 1 transition plays a major role in the S to M rise. To further understand the nature of the SM fluorescence rise, we used (1) 3-(3,4-dichlorophenyl)-1,4-dimethyl urea (DCMU), an inhibitor of electron transport between PS II and PS I; (2) n-propyl gallate (PG), an inhibitor of alternative oxidase (AOX) in mitochondria and of plastid terminal oxidase (PTOX) in chloroplasts; (3) salicylhydroxamic acid (SHAM), an inhibitor of AOX in mitochondria; and (4) carbonyl cyanide p-trifluoromethoxyphenylhydrazone (FCCP), an uncoupler of electron transport from phosphorylation, which dissipates proton gradient (see scheme in Fig. 1). A preliminary report of this work was presented in a poster by Kodru et al. (2013); here, we present all the observations of the above-mentioned experiments.
Based on results presented in this paper, and in the literature (see above), we suggest that the P to S decline, as well as the S to M rise, and the M to T decline in Chl a fluorescence transient, are due to superimposition of at least two phenomena: qE dependent non-photochemical quenching (NPQ) of the excited state of Chl, and state transitions. However, other processes may also contribute to the SMT phase, besides those mentioned above (see discussion on the slow SMT phase and Chl fluorescence oscillations in Stirbet et al. (2014)). Papageorgiou et al. (2007) and Bernát et al. (2014) have shown that in cyanobacteria, the M to T decline may be due to photoinhibition; however, we do not know if the same is true for C. reinhardtii. For a discussion on the relation of photoinhibition to state changes, see Canaani et al. (1989) and Finazzi et al. (2001).
Materials and methods
Growth conditions
Cells of wild type C. reinhardtii strain CC125 and of the stt7 mutant were grown at 25 °C in TAP (tris-acetate phosphate) medium (Gorman and Levine 1965), under continuous white light (60–70 µmol photons m−2 s−1), to an optical density of ~1.0.
Chemical treatment
Actively growing cells were harvested, and transferred into a test tube; this cell suspension was kept in dark for 30 min at room temperature. One ml of this dark-adapted suspension was transferred to the sample tube of the fluorescence instrument (see below) and used as control, or mixed thoroughly with (1) 10 µM Diuron, DCMU, or (2) 1 mM PG, or (3) 1 mM SHAM, or (4) 10 µM FCCP. These samples were then kept in dark for 10 min before fluorescence measurements (see below). The chosen concentration, as well as time with each chemical, was based on earlier research, cited under “Results and discussion”.
The OJIPSMT Chl a fluorescence transient measurements
Chlorophyll a fluorescence induction measurements of these dark-adapted C. reinhardtii cells were made with Handy PEA (Plant Efficiency Analyzer) fluorometer (Hansatech, King’s Lynn, Norfolk, UK). In the OJIPSMT transient (see Introduction), O is for the minimum fluorescence (also referred to as F o), measured as soon as light is turned on, J (F J) and I (F I) are two inflections, P (F P) is the peak (maximum fluorescence, or F m in saturating light), S is for semi-steady state, M is for another maximum, and T is for terminal steady state. (For a discussion of nomenclature, see Govindjee 1995, 2004, and for analysis, see Strasser et al. 2004.) Dark-adapted C. reinhardtii cells were excited with 650 nm light (1,000 μmol photons m−2 s−1), obtained from LEDs. These samples were not stirred before and during measurements. In order to compare measurements on different samples, F(t), fluorescence at time t was normalized as V(t) = [F(t) − F o]/(F m − F o), where V(t) represents relative variable fluorescence (see e.g., Stirbet and Govindjee 2011). Such a method allowed us to compare kinetics in different samples.
Results and discussion
SM rise in OJIPSMT fluorescence transients
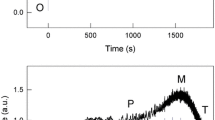
Chl a fluorescence transients were recorded on dark-adapted (see “Materials and methods”) wild type and stt7 mutant C. reinhardtii cells by illuminating them with 650-nm light of 1,000 μmol photons m−2 s−1 (Fig. 2). In wild type cells, we observe both the faster OJIP (up to ~200 ms) and the slower PSMT (P to S decline, followed by SMT, the second wave lasting up to ~300 s) phase of the transient. Under our experimental conditions, the appearance of the SM rise was prominent when recorded with 1,000 μmol photons m−2 s−1 light (see Fig. 2), while at higher light intensities (e.g., 3,000 μmol photons m−2 s−1), the SM phase was less evident (data not shown). This was expected since the two phases (OJIP and SMT) have different reactions that control them (the fast phase by the electron flow and the slow phase by other regulatory events; see e.g., Stirbet et al. 2014); further, it is in agreement with earlier results on Chlorella (see Papageorgiou and Govindjee 1968a; Munday and Govindjee 1969). Since the focus of this study is the SM rise phase of the fluorescence induction curve, we chose 1,000 μmol photons m−2 s−1 light (Fig. 2) for all fluorescence induction measurements.

(A) Chl a fluorescence transient (OJIPSMT) in wild type (wt) and stt7 mutant C. reinhardtii cells, plotted on logarithmic time scale; and (B) the same on a linear time scale. (Data are double normalized at the O and P levels.) Excitation of samples was with 1,000 µmol photons m−2 s−1 650 nm light (here as well as in Figs. 3, 4, 5, 6). One of the several possible components involved in quenching or stimulation of fluorescence are shown in red
Fast and slow fluorescence transients; link between SM rise and state transitions
Spalding et al. (1984), Govindjee et al. (1991), and Srivastava et al. (1995) have published O(JI)PS transients in wild type as well as in various PS II mutants of C. reinhardtii. Here, we have measured OJIPSMT transients in both wild type and stt7 mutant of C. reinhardtii, the latter is known not to show state transitions (Depege et al. 2003) since it is locked in state 1. As noted above, Fig. 2 shows fluorescence transients in these two samples when cells were exposed to 1,000 μmol photons m−2 s−1 of 650 nm light. The fast fluorescence transient (OJIP) in the wild type and in the stt7 cells showed some differences (see discussion below), but had similar general features (see Fig. 2A, plotted on logarithmic time scale). However, after the P level, the slow SMT phase in the two strains differed significantly (see Fig. 2B, plotted on a linear time scale).
As can be seen in Fig. 2A, the fast OJIP phases in wt and stt7 differ in their initial slopes of the OJ rise (calculated as M o ≈ (ΔV/Δt o)/V J), where ΔV = (F 0.3 ms − F o)/(F m − F o), F o = F 0.02 ms, Δt o = 0.28 ms, and V J = (F 2 ms − F o)/(F m − F o); Strasser et al. 2004), as well as in (the relative) fluorescence levels at J (2 ms) and I (30 ms). The initial slope of the OJ rise in wt was less steep than in the stt7 mutant (0.6 vs. 0.94), which suggests a lower PS II absorption cross section, σ(PS II), in the wild type (see e.g., Stirbet and Govindjee 2011). Further, the J level was lower in wt (0.2 vs. 0.28 in stt7), which indicates a lower Q −A accumulation at this step, due to lower light excitation resulting from smaller PS II absorption cross section. Finally, the I level was lower in wt compared to stt7 (0.5 vs. 0.6). (For discussion, see Ceppi et al. 2012.) The above results suggest that after dark-adaptation, the wt cells might be in a state closer to state 2, since they had a lower inferred PS II absorption cross section, and a higher PS I activity than the stt7 cells, which are locked in state 1.
We note that Nellaepalli et al. (2013) have also reported differences in fast Chl a fluorescence transients (OJIP) under state 1 (high fluorescence state) and state 2 (low fluorescence state) in Arabidopsis thaliana. We speculate that this may have been due to insufficient oxygen level (or slightly anaerobic condition) during the dark-adaptation period in TAP medium, since the cells were not stirred. Indeed, C. reinhardtii cells, grown mixotrophically, attain state 1 only through vigorous stirring (Wollman and Lemaire 1988; Takahashi et al. 2013). Further, Madireddi et al. (2014) reported LHC II phosphorylation in wild type C. reinhardtii cells grown in TAP medium under low light conditions indicating that these cells were in state 2 under those conditions. These results support our conclusion that the wild type C. reinhardtii cells were, most likely, in state 2 at the beginning of our fluorescence measurements.
The P to S fluorescence decline was observed during 0.2–2 s, the S to M rise from 2 to 70 s, and the MT decline from 70 to 300 s. There are remarkable differences between the wt and the stt7 mutant during their P to S decline. The decrease in fluorescence from P to S is larger and reached a lower semi-steady state S level in wt compared to that in stt7 cells (Fig. 2). The PS phase has been often correlated partly with qE, the energy-dependent NPQ of the excited Chl (Demmig-Adams 1990), which has been suggested to be initiated by protonation events, as inferred from the work of Briantais et al. (1979, 1980, 1986). This quenching, however, may not involve xanthophyll cycle. For earlier observations on the OPS transient in wild type C. reinhardtii and their NPQ mutants, see e.g., Govindjee and Seufferheld (2002) and Holub et al. (2007).
The S to M rise, during fluorescence transient, has been related to structural changes in the thylakoid membrane by Papageorgiou (1968) and Papageorgiou and Govindjee (1968a); see a review by Papageorgiou and Govindjee (2011). In our experiments, we observed that the SM rise is essentially absent in the stt7 mutant (Fig. 2), which is locked in state 1. This suggests that the SM rise in the wild type C. reinhardtii is due to state 2 (low fluorescence state, larger antenna in PS I) to state 1 (high fluorescence state; larger antenna in PS II) transition. Such a state change has been shown to take place as a result of LHC II kinase inactivation induced by high light, when LHC IIs are dephosphorylated and migrate from PS I to PS II region (Rintamaki et al. 2000). Holub et al. (2007) did observe, using 2,750 µmol photons m−2 s−1, SM rise in wild type and npq1 (violaxanthin accumulating) mutant, but not in npq2 (zeaxanthin accumulating) mutant. However, based on lifetime (and fractional amplitude) fluorescence analyses, they were unable to reach any conclusion as to its relation to state changes.
Finally, in analogy with the P to S decline, we suggest that the M to T decline that follows the SM rise (Fig. 2) may be also partly due to qE. In the stt7 mutant, which is locked in state 1, the MT decline can not be affected by state transition, and therefore appears more pronounced than in wt cells (Fig. 2). However, we note that photoinhibition (Adir et al. 2003; Papageorgiou et al. 2007; Murata et al. 2012) can also contribute to this decline, as shown in Synechocystis PCC 6803 (Bernát et al. 2014).
The concept that SM rise is a state 2 to state 1 transition is supported, as noted earlier, by the fact that Synechocystis mutant (RpaC −), locked in state 1, does not show S to M fluorescence rise (Kaňa et al. 2012). Moreover, in a non-photochemical fluorescence quenching study of C. reinhardtii, Allorent et al. (2013) found that wt cells, which were initially in state 2, after high light adaptation, showed a transient increase in fluorescence during illumination, which they assigned to a state 2 to state 1 transition, as it was absent in the stt7-9 mutant (locked in state 1) and the phosphorylation level of LHC II decreased upon illumination. Since this transient fluorescence increase in light-adapted sample, and the SM rise observed in our experiments, are related, these results reinforce our conclusion that SM rise is due to state changes. Thus, we suggest that measurements on SMT transient may be used as a quick monitor of state changes in green algae as already known for several cyanobacteria (Papageorgiou et al. 1999, 2007; Kaňa et al. 2012). For discussion of state changes in an atypical cyanobacterium Gloeobacter violaceus, see Bernát et al. (2012).
Role of the redox state of PQ pool on the SM rise
The redox state of PQ pool is an important factor in the regulation of several processes, such as LHC II phosphorylation during state transition, metabolic adjustment, or translation of gene expression (Dietz and Pfannschmidt 2011). Mus et al. (2005) have studied the effect of various inhibitors on the redox status of PQ in C. reinhardtii; thus, we tested, although indirectly, the role of the redox state of the PQ pool in the SM phase of Chl a fluorescence transient as follows.
DCMU (Diuron)
It is well known that high level of Chl fluorescence in the presence of Diuron (DCMU) is due to the closure of reaction center II, which is caused by the presence of high concentration of Q −A (see Duysens and Sweers 1963). This is because DCMU binds to the QB site and displaces it (Velthuys 1981; Lavergne 1982), and, thus, inhibits electron transport from reduced Q A to the PQ pool (see Fig. 1; in this scheme, we do not show the precise location where DCMU functions, but only its effect). Photosystem I would oxidize PQH2 during illumination and the cells would go in state 1 (high fluorescence state; see e.g., Allorent et al. 2013) because a phosphatase would be activated leading to movement of mobile LHCII from PS I to PS II (see e.g., Minagawa 2013). (For earlier work, see e.g., Wollman and Delepelaire 1984; Delepelaire and Wollman 1985; Wollman and Bulté 1990.)
Our experiments confirm that upon addition of 10 μΜ DCMU, fluorescence yield increases very rapidly, reaching maximum fluorescence (F m) without showing the distinctive J and I inflections (see Fig. 3) that are present in untreated samples (Strasser and Govindjee 1992). During the entire measurement, fluorescence yield remains relatively close to maximum, decreasing only slightly, and the transient does not show the PSMT phase (Fig. 3). Similar results were obtained in DCMU-treated stt7 cells (data not shown). However, we note that in contrast, the SM rise in cyanobacteria is enhanced in the presence of DCMU due to increased energy transfer from phycobilisomes to PS II that induces a high fluorescence state 1 (Tsimilli-Michael et al. 2009; Kaňa et al. 2012; Stamatakis and Papageorgiou 2014).

Chl a fluorescence transient (OJIPSMT) in wild type C. reinhardtii cells (Control) and those that had been for 10 min in darkness, with 10 µM DCMU (3-(3,4-dichlorophenyl)-1,4-dimethyl urea) (+DCMU). Data are double normalized at the O and P levels and plotted on logarithmic time scale. DCMU is known to block electron flow beyond Q A in PS II by displacing Q B (Velthuys 1981)
n-Propyl gallate
n-Propyl gallate inhibits both the AOX in mitochondria (Siedow and Bickett 1981), as well as the plastid (or plastoquinol) terminal oxidase (PTOX) in chloroplasts (Cournac et al. 2000a, b); this inhibition leads, for example, to ~60 % inhibition of O2 uptake rate in Vicia faba leaves (Yoshida et al. 2006). AOX is a non-energy conserving terminal oxidase in the mitochondrial electron transport chain of algae and higher plants, which directly couples the oxidation of ubiquinol with the reduction of O2 to H2O (see a review by Vanlerberghe 2013). PTOX participates in chlororespiration (Bennoun 1994; Nixon 2000; Peltier and Cournac 2002; see Fig. 1), which involves electron donation from NADPH to PQ via Ndh (i.e., NAD(P)H dehydrogenase in plants; Rumeau et al. 2007) and NDA2 (type II NADPH dehydrogenase in C. reinhardtii; Jans et al. 2008, Desplats et al. 2009), and PQH2 oxidation (coupled with O2 reduction to H2O) by PTOX (Bennoun 1982; Houille-Vernes et al. 2011). With PTOX inhibited by PG treatment, non-photochemical reduction of the PQ pool will be promoted (Cournac et al. 2000a, b; Bennoun 2001; Peltier and Cournac 2002), and the cells will have a tendency to remain in state 2 (low fluorescence state; see e.g., Forti and Caldiroli 2005). Moreover, Yoshida et al. (2006) showed that in light-adapted PG-treated V. faba leaves, non photochemical quenching of Chl a fluorescence was increased, while the photodamage induced under high light was accelerated.
Chlorophyll a fluorescence measurements in samples with 1-mM PG showed that the M level reached faster, but was much lower than in the control (see Fig. 4). These changes in the slow fluorescence phase suggest a relation of the SM rise to the redox state of the PQ pool, and, thus, to state transitions. The non-photochemical reduction of PQ pool in the presence of PG is indicated in the fluorescence curves by an increased J level compared to that in untreated cells (Fig. 4A), since a higher J level was shown to reflect a higher reduced state of PQ pool, established in darkness prior to the measurement (Haldimann and Strasser 1999; Tóth et al. 2007; Tsimilli-Michael et al. 2009). Thus, these results further support the origin of SM rise to be due to state 2 to state 1 transition. However, the P to S decline was found to be similar in both control and PG-treated samples (Fig. 4B), indicating that qE-NPQ was not affected by PG treatment. Finally, during the MT phase in wt cells, and after the S level in cells with PG present, fluorescence remained relatively constant in both the samples (Fig. 4), which might suggest that qE was already at its maximum under these conditions.

(A) Chl a fluorescence transient (OJIPSMT) in wild type C. reinhardtii cells (Control) and those that had been for 10 min in darkness, with 1 mM n-propyl gallate (propyl 3,4,5-trihydroxybenzoate) (+PG)—plotted on logarithmic time scale; and (B) the same on a linear time scale. (Data are double normalized at the O and P levels.) PG inhibits both the alternate oxidase (AOX) in mitochondria (Siedow and Bickett 1981) and the plastid terminal oxidase (PTOX) in chloroplasts (Cournac et al. 2000a, b)
Salicylhydroxamic acid
SHAM is known to inhibit AOX pathway in mitochondria (Elthon and Mcintosh 1987; Siedow and Umbach 2000; Fig. 1), leading, for example, to ~60 % inhibition of O2 uptake in V. faba leaves (Yoshida et al. 2006). However, the effect of SHAM on PTOX is extremely low in C. reinhardtii cells, even at high concentrations (Cournac et al. 2002). Photosynthesis and mitorespiration have been shown to interact through ATP, NADPH, and metabolite exchange between chloroplasts and mitochondria (Hoefnagel et al. 1998; see Fig. 1). It is now known that mitochondrial metabolism influences photosynthetic performance (see a review by Araújo et al. 2014). Inhibition of AOX by SHAM was shown to trigger NADPH accumulation in chloroplasts that causes over-reduction of both the electron acceptor side of PS I and the PQ pool due to NDA2 activity (Yoshida et al. 2006; Zhang et al. 2012), which can keep the algal cells in state 2, as stated earlier (for the relation between respiration and state transitions in C. reinhardtii, see Cardol et al. (2009)). Moreover, studies on SHAM-treated V. faba leaves (Yoshida et al. 2006) showed that, during the light-adapted state, the photochemical quenching (i.e., qP), of the excited state of chlorophyll, was markedly suppressed, while the photodamage induced under high light was accelerated.
Here, we have examined the effects of 1-mM SHAM on Chl a fluorescence transient (see Fig. 5). We observed a significant increase in the initial fluorescence F o and the J level, and the OJ rise was followed by a dip before fluorescence reached F m (Fig. 5A shows only the normalized data). Further, the SM rise was abolished, with fluorescence decreasing slightly after the S level (Fig. 5B). We note that changes observed in the OJIP phase in the presence of SHAM are similar to those reported in samples kept under anoxic conditions (i.e., high F o, J level even higher than the I level, and low IP phase (see Fig. 5A and results published by Haldimann and Strasser 1999; Antal et al. 2001; Tsimilli-Michael et al. 2009; Hohmann-Marriott et al. 2010); these reflect a highly reduced PQ pool and, thus, state 2. We note that Hohmann-Marriott et al. (2010) had used a mutant that does not assemble PS II and PS I core complexes (therefore, it has no Q A), yet its PQ pool suppresses F o when it is oxidized, and increases it when reduced. We do not yet know the mechanism behind it. Changes in fluorescence induction in SHAM-treated cells observed here are also related to the PQ pool reduction, which is accompanied by oxidation of excess NADPH (catalyzed by NDA2; see Fig. 1) that is generated as a result of AOX inhibition (Yoshida et al. 2006; Zhang et al. 2012; Araújo et al. 2014); under these conditions, cells have the tendency to remain in state 2. Therefore, the above results support the idea of a connection between mitorespiration and state transitions in C. reinhardtii (see also Cardol et al. 2009), and show that even the inhibition of AOX pathway by SHAM treatment can diminish or even eliminate the SM rise.

(A) Chl a fluorescence transient (OJIPSMT) in wild type C. reinhardtii cells (Control) and those that had been for 10 min in darkness with 1 mM salicylhydroxamic acid (+SHAM), plotted on a logarithmic time scale; and (B) the same on a linear time scale. (Data are double normalized at the O and P levels.) SHAM is known to inhibit alternative oxidase (AOX) in mitochondria (Elthon and Mcintosh 1987; Siedow and Umbach 2000)
Carbonyl cyanide trifluoromethoxyphenylhydrazone
FCCP is an ionophore that uncouples electron flow from phosphorylation by dissipating ΔpH gradient (see Fig. 1), so that a decrease of the ATP level in the cell is expected. A series of events that influence the balance of chlororespiration follow this decrease in the ATP level (see Fig. 1 and Bulté et al. 1990; Wollman 2001): (a) glycolysis in cytoplasm would be stimulated (Rebeille and Gans 1988); (b) inter-organelle transport of reducing equivalents between chloroplasts and mitochondria through the oxaloacetate−malate shuttle would occur (Bulté et al. 1990; Bennoun 1994; see Fig. 1) leading to increased NADPH levels in the chloroplast; this in turn (c) would reduce the PQ pool via NADPH oxidation by NDA2 (see e.g., Jans et al. 2008), thus keeping the algal cells in state 2 (see e.g., Allorent et al. 2013).
FCCP (10 µM) has been shown to inhibit the SM rise in Anacystis nidulans, and to abolish the MT decline in Chlorella pyrenoidosa (Papageorgiou and Govindjee 1968a, b). Also, as discussed above, excess NADPH that is generated due to FCCP treatment reduces the PQ pool (via NDA2) and induces a transition to state 2 in C. reinhardtii cells (see Fig. 1; Bulté et al. 1990; Houille-Vernes et al. 2011; Allorent et al. 2013). Our results show that 10 µM FCCP largely inhibits the SM rise (Fig. 6A). Since FCCP is an uncoupler of proton gradient, thus, affecting the redox state of PQ pool, these data are in agreement with the thesis that state 2 to state 1 transition is involved in the SM rise. The increased J level, observed in the FCCP-containing sample, compared to that in the controls (see Fig. 6A), may suggest that FCCP induces non-photochemical reduction of the PQ pool in darkness, which slows Q A reoxidation during illumination and leads to Q −A accumulation at the J level. Further, our fluorescence data show only a slightly smaller PS decline in FCCP-containing samples than in the controls (Fig. 6B), which may suggest that 10 μM FCCP was not sufficient to completely abolish the pH gradient (and thus qE). The effect of uncoupling on the kinetics and redox level of PQ, however, remains to be measured. On the other hand, we have also observed a steeper fluorescence decrease during the MT phase (Fig. 6B), which may reflect an increased photosensitivity of samples to FCCP, leading to photoinhibition.

(A) Chl a fluorescence transient (OJIPSMT) in wild type C. reinhardtii cells (Control) and those that had been for 10 min in darkness with 10 µM FCCP (carbonyl cyanide p-trifluoromethoxyphenylhydrazone) (+FCCP), plotted on logarithmic time scale; and (B) the same on a linear time scale. (Data are double normalized at the O and P levels.) FCCP is an uncoupler of proton gradient that decreases ATP level in cells (Bulté et al. 1990)
The above results on the effect of chemicals, which change the redox state of the PQ pool, strongly support the concept that the SM rise has a significant contribution from state 2 to state 1 transition. Together with the absence of SM rise in stt7 mutant, our conclusion is that the SM rise is indeed a signature of state 2 to state 1 transition in C. reinhardtii.
Concluding remarks
Using the stt7 mutant, and various inhibitors (DCMU, PG, salicylhydroxamic acid, and FCCP), we conclude that the slow SM Chl fluorescence rise can be used as a signature of state 2 to state 1 transition in the green alga C. reinhardtii.
It is now necessary to extend this work (1) to see the effect of an osmolyte glycine betaine, as was used by Papageorgiou and Stamatakis (2004) in cyanobacteria;Footnote 1 and (2) to see its relation to RNA helicase involved in energy redistribution, also in cyanobacteria (Sireesha et al. 2012).
Further, we suggest that the entire PSMT fluorescence transient is due to a superimposition of several processes in which qE (energy-dependent NPQ of Chl fluorescence), as well as state changes play an important role. Moreover, the possible involvement of photoinhibition during the MT decline must not be ignored.
Further research with mutants blocked in different steps of these processes is expected to lead to a better understanding of how Chl fluorescence could be further exploited to obtain insights into the physiology of cyanobacteria, algae, and plants.
Notes
In the Supplementary Material, which is not a part of the main text, we have provided our preliminary results on the effect of using a compatible osmolyte (N,N,N-trimethyl-glycine (C5H11NO2); also called glycine betaine) for the benefit of those who may be asking if its use would have helped us in answering the question of relation between state changes and the SM rise, as it had in cyanobacteria (see e.g., Papageorgiou and Stamatakis 2004). Although our results are complex, and, further studies are necessary to examine these issues, we have made it available to researchers interested in it.
Abbreviations
- AOX:
-
Alternative oxidase
- CEF:
-
Cyclic electron flow
- Chl:
-
Chlorophyll
- Cyt:
-
Cytochrome
- DCMU:
-
3-(3,4-Dichlorophenyl)-1,4-dimethyl urea; also known as Diuron
- FCCP:
-
Carbonyl cyanide p-trifluoromethoxyphenylhydrazone
- LEF:
-
Linear electron flow
- NADP:
-
Nicotinamide adenine dinucleotide phosphate
- Ndh:
-
NAD(P)H dehydrogenase
- NDA2:
-
Type II NADPH dehydrogenase
- OJIPSMT:
-
Chl a fluorescence transient, where “O” refers to the minimum fluorescence, J and I for inflections, P for peak, S for semi-steady state, M for maximum and T for terminal steady state
- PC:
-
Plastocyanin
- PG:
-
n-Propyl gallate
- pmf :
-
Proton-motive force
- PQ, PQH2 :
-
Plastoquinone, plastoquinol
- PS:
-
Photosystem
- PTOX:
-
Plastid (or plastoquinol) terminal oxidase
- SHAM:
-
Salicylhydroxamic acid
References
Adir N, Zer H, Schochat S, Ohad I (2003) Photoinhibition—a historical perspective. Photosynth Res 76:343–370
Allen JF, Mullineaux CW (2004) Probing the mechanism of state transitions in oxygenic photosynthesis by chlorophyll fluorescence spectroscopy, kinetics and imaging. In: Papageorgiou GC, Govindjee (eds) Chlorophyll fluorescence—a signature of photosynthesis, vol 19. Springer, Dordrecht, pp 447–461
Allen JF, Bennett J, Steinback KE, Arntzen CJ (1981) Chloroplast protein phosphorylation couples PQ redox state to distribution of excitation-energy between photosystems. Nature 291:25–29
Allorent G, Tokutsu R, Roach T, Peers G, Cardol P, Girard-Bascou J, Finazzi G (2013) A dual strategy to cope with high light in Chlamydomonas reinhardtii. Plant Cell 25:545–557
Alric J (2010) Cyclic electron flow around photosystem I in unicellular green algae. Photosynth Res 106:47–56
Andersson B, Åkerlund HE, Jergil B, Larsson C (1982) Differential phosphorylation of the light-harvesting chlorophyll–protein complex in appressed and non-appressed regions of the thylakoid membrane. FEBS Lett 149:181–185
Antal TK, Krendeleva TE, Laurinavichene TV, Makarova VV, Tsygankov AA, Seibert M, Rubin AB (2001) The relationship between the photosystem 2 activity and hydrogen production in sulfur deprived Chlamydomonas reinhardtii cells. Doklady Biochem Biophys 381:371–374
Araújo WL, Nunes-Nesi A, Fernie AR (2014) On the role of plant mitochondrial metabolism and its impact on photosynthesis in both optimal and sub-optimal growth conditions. Phosynth Res 119:141–156
Baker NR (2008) Chlorophyll fluorescence: a probe of photosynthesis in vivo. Annu Rev Plant Biol 59:89–113
Bellafiore S, Barneche F, Peltier G, Rochaix J-D (2005) State transitions and light adaptation require chloroplast thylakoid protein kinase STN7. Nature 433:892–895
Bennett J, Steinback KE, Arntzen CJ (1980) Chloroplast phosphoproteins regulation of excitation-energy transfer by phosphorylation of thylakoid membrane polypeptides. Proc Natl Acad Sci USA 77:5253–5257
Bennoun P (1982) Evidence for a respiratory chain in the chloroplast. Proc Natl Acad Sci USA 79:4352–4356
Bennoun P (1994) Chlororespiration revisited: mitochondrial–plastid interactions in Chlamydomonas. Biochim Biophys Acta 1186:59–66
Bennoun P (2001) Chlororespiration and the process of carotenoid biosynthesis. Biochim Biophys Acta 1506:133–142
Bernát G, Schreiber U, Sendtko E, Stadnichuk IN, Sascha Rexroth S, Matthias Rögner M, Koenig F (2012) Unique properties vs. common themes: the atypical cyanobacterium Gloeobacter violaceus PCC 7421 is capable of state transitions and blue-light-induced fluorescence quenching. Plant Cell Physiol 53:528–542
Bernát G, Kaňa R, Steinbach G, Govindjee, Misra AN, Kirilovsky D, Prašil O (2014) The very slow M to T chlorophyll fluorescence decay in cyanobacteria reflects photoinhibition. In: 9th European workshop on the molecular biology of cyanobacteria, Texel (the Netherlands), 7–11 September 2014, p 160
Blankenship RE (2014) Molecular mechanism of photosynthesis, 2nd edn. Blackwell, Oxford
Bonaventura C, Myers J (1969) Fluorescence and oxygen evolution from Chlorella pyrenoidosa. Biochim Biophys Acta 189:366–383
Briantais J-M, Vernotte C, Picaud M, Krause GH (1979) A quantitative study of the slow decline of chlorophyll a fluorescence in isolated chloroplasts. Biochim Biophys Acta 548:128–138
Briantais J-M, Vernotte C, Picaud M, Krause GH (1980) Chlorophyll fluorescence as a probe for the determination of the photo-induced proton gradient in isolated chloroplasts. Biochim Biophys Acta 591:198–202
Briantais J-M, Vernotte C, Krause GH, Weis E (1986) Chlorophyll a fluorescence of higher plants: chloroplasts and leaves. In: Govindjee, Amesz J, Fork DC (eds) Light emission by plants and bacteria. Academic Press, Orlando, pp 539–583
Bulté L, Gans P, Rebeille F, Wollman F-A (1990) ATP control on state transitions in vivo in Chlamydomonas reinhardtii. Biochim Biophys Acta 1020:72–80
Canaani O, Schuster G, Ohad I (1989) Photoinhibition in Chlamydomonas reinhardtii: effect of state transition, intersystem energy distribution and photosystem I cyclic electron flow. Photosynth Res 20:129–146
Cardol P, Alric J, Girard-Bascou J, Franck F, Wollman F-A, Finazzi G (2009) Impaired respiration discloses the physiological significance of state transitions in Chlamydomonas. Proc Natl Acad Sci USA 106:15979–15984
Ceppi MG, Oukarroum A, Çiçek N, Strasser RJ, Schansker G (2012) The IP amplitude of the fluorescence rise OJIP is sensitive to changes in the photosystem I content of leaves: a study on plants exposed to magnesium and sulfate deficiencies, drought stress and salt stress. Physiol Plantarum 144:277–288
Cournac L, Josse EM, Joet T, Rumeau D, Redding K, Kuntz M, Peltier G (2000a) Flexibility in photosynthetic electron transport: a newly identified chloroplast oxidase involved in chlororespiration. Philos Trans R Soc Lond Ser B Biol Sci 355:1447–1453
Cournac L, Redding K, Ravenel J, Rumeau D, Josse EM, Kuntz M, Peltier G (2000b) Electron flow between Photosystem II and oxygen in chloroplasts of Photosystem I-deficient algae is mediated by a quinol oxidase involved in chlororespiration. J Biol Chem 275:17256–17262
Cournac L, Latouche G, Cerovic Z, Redding K, Ravenel J, Peltier G (2002) In vivo interactions between photosynthesis, mitorespiration, and chlororespiration in Chlamydomonas reinhardtii. Plant Physiol 129:1921–1928
DeEll JR, Toivnen PMA (eds) (2003) Practical applications of chlorophyll fluorescence in plant biology. Kluwer (now Springer), Dordrecht
Delepelaire P, Wollman F-A (1985) Correlations between fluorescence and phosphorylation changes in thylakoid membranes of Chlamydomonas reinhardtii in vivo: a kinetic analysis. Biochim Biophys Acta 809(2):277–283
Delosme R, Olive J, Wollman FA (1996) Changes in light energy distribution upon state transitions: an in vivo photoacoustic study of the wild type and photosynthesis mutants from Chlamydomonas reinhardtii. Biochim Biophys Acta 1273:150–158
Demmig-Adams B (1990) Carotenoids and photoprotection in plants: a role for the xanthophyll zeaxanthin. Biochim Biophys Acta 1020:1–24
Demmig-Adams B, Garab G, Adams WWI, Govindjee (eds) (2014) Non-photochemical quenching and energy dissipation in plants, algae and cyanobacteria, advances in photosynthesis and respiration, vol 40. Springer, Dordrecht
Depege N, Bellafiore S, Rochaix JD (2003) Role of chloroplast protein kinase Stt7 in LHCII phosphorylation and state transition in Chlamydomonas. Science 299:1572–1575
Desplats C, Mus F, Cuiné S, Billon E, Cournac L, Peltier G (2009) Characterization of Nda2, a plastoquinone-reducing type II NAD(P)H dehydrogenase in Chlamydomonas chloroplasts. J Biol Chem 284:4148–4157
Dietz KJ, Pfannschmidt T (2011) Novel regulators in photosynthetic redox control of plant metabolism and gene expression. Plant Physiol 155:1477–1485
Duysens LNM, Sweers HT (1963) Mechanism of the two photochemical reactions in algae as studied by means of fluorescence. In: Japanese Society of Plant Physiologists (ed) Studies on microalgae and photosynthetic bacteria. University of Tokyo Press, Tokyo, pp 353–372
Elthon TE, McIntosh L (1987) Identification of the alternative terminal oxidase of higher plant mitochondria. Proc Natl Acad Sci USA 84:8399–8403
Finazzi G, Barbagallo RP, Bergo E, Barbato R, Forti G (2001) Photoinhibition of Chlamydomonas reinhardtii in State 1 and State 2: damages to the photosynthetic apparatus under linear and cyclic electron flow. J Biol Chem 276:22251–22257
Forti G, Caldiroli G (2005) State transitions in Chlamydomonas reinhardtii: the role of the Mehler reaction in State 2-to-State 1 transition. Plant Physiol 137:492–499
Gorman DS, Levine RP (1965) Cytochrome f and plastocyanin: their sequence in the photosynthetic electron transport chain of Chlamydomonas reinhardii. Proc Natl Acad Sci USA 54:1665–1669
Govindjee (1995) Sixty-three years since Kautsky: chlorophyll a fluorescence. Aust J Plant Physiol (now Funct Plant Biol) 22:131–160
Govindjee (2004) Chlorophyll a fluorescence: a bit of basics and history. In: Papageorgiou GC, Govindjee (eds) Chlorophyll a fluorescence: a signature of photosynthesis, vol 19. Advances in photosynthesis and respiration. Springer, Dordrecht, pp 2–42
Govindjee, Seufferheld MJ (2002) Non-photochemical quenching of chlorophyll a fluorescence: early history and characterization of two xanthophyll cycle mutants of Chlamydomonas reinhardtii. Funct Plant Biol 29:1141–1155
Govindjee, Amesz J, Fork DC (eds) (1986) Light emission by plants and bacteria. Academic Press, New York (now Elsevier, Amsterdam)
Govindjee, Schwarz B, Rochaix J-D, Strasser RJ (1991) The herbicide-resistant D1 mutant L275F of Chlamydomonas reinhardtii fails to show the bicarbonate-reversible formate effect on chlorophyll a fluorescence transients. Photosynth Res 27:199–208
Haldimann P, Strasser RJ (1999) Effects of anaerobiosis as probed by the polyphasic chlorophyll a fluorescence rise kinetic in pea (Pisum sativum L.). Photosynth Res 62:67–83
Hemschemeier A, Happe T (2011) Alternative photosynthetic electron transport pathways during anaerobiosis in the green alga Chlamydomonas reinhardtii. Biochim Biophys Acta 1807:919–926
Hoefnagel MHN, Atkin OK, Wiskich JT (1998) Interdependence between chloroplasts and mitochondria in the light and the dark. Biochim Biophys Acta 1366:235–255
Hohmann-Marriott HM, Takizawa K, Eaton-Rye JJ, Mets L, Minagawa J (2010) The redox state of the plastoquinone pool directly modulates minimum chlorophyll fluorescence yield in Chlamydomonas reinhardtii. FEBS Lett 584:1021–1026
Holub O, Seufferheld MJ, Gohlke C, Govindjee, Heiss GJ, Clegg RM (2007) Flourescence lifetime imaging microscopy of Chlamydomonas reinhardtii: non-photochemical quenching mutants and the effect of photosynthetic inhibitors on the slow chlorophyll fluorescence transient. J Microsc 226:90–120
Houille-Vernes L, Rappaport F, Wollman F-A, Alric J, Johnson X (2011) Plastid terminal oxidase 2 (PTOX2) is the major oxidase involved in chlororespiration in Chlamydomonas. Proc Natl Acad Sci USA 108:20820–20825
Iwai M, Yokono M, Inada N, Minagawa J (2010) Live-cell imaging of photosystem II antenna dissociation during state transitions. Proc Natl Acad Sci USA 107:2337–2342
Jans F, Mignolet E, Houyoux PA, Cardol P, Ghysels B, Cuiné S, Cournac L, Peltier G, Remacle C, Franck F (2008) A type II NAD (P) H dehydrogenase mediates light-independent plastoquinone reduction in the chloroplast of Chlamydomonas. Proc Natl Acad Sci USA 105:20546–20551
Kalaji MH, Goltsev V, Bosa K, Allakhverdiev IS, Strasser JR, Govindjee (2012a) Experimental in vivo measurements of light emission in plants: a perspective dedicated to David Walker. Photosynth Res 114:69–96
Kalaji MH, Carpentier R, Allakhverdiev SI, Bosa K (2012b) Fluorescence parameters as an early indicator of light stress in barley. J Photochem Photobiol B 112:1–6
Kalaji MH, Schansker G, Ladle RJ, Goltsev V, Bosa K, Allakhverdiev SI, Brestic M, Bussotti F et al (2014) Frequently asked questions about in vivo chlorophyll fluorescence: practical issues. Photosynth Res 122:121–158
Kaňa R, Kotabová E, Komárek O, Sedivá B, Papageorgiou GC, Govindjee, Prášil O (2012) The slow S to M fluorescence rise in cyanobacteria is due to a state 2 to state 1 transition. Biochim Biophys Acta 1817:1237–1247
Kodru S, Nellaepalli S, Malavath T, Devadasu E, Subramanyam R, Govindjee (2013) Does the slow S to M rise of chlorophyll a fluorescence induction reflect transition from state 2 to state 1 in the green alga Chlamydomonas reinhardtii? In: 16th international photosynthesis congress, St. Louis, MO, August 2013
Lavergne J (1982) Mode of action of 3-(3,4-dichlorophenyl)-1,1-dimethylurea. Evidence that the inhibitor competes with plastoquinone for binding to a common site on the acceptor side of Photosystem II. Biochim Biophys Acta 682:345–353
Lichtenthaler HK (ed) (1988) Applications of chlorophyll fluorescence. Springer, Dordrecht
Madireddi SK, Nama S, Devadasu ER, Subramanyam R (2014) Photosynthetic membrane organization and role of state transition in cyt, cpll, stt7 and npq mutants of Chlamydomonas reinhardtii. J Photochem Photobiol B 137:77–83
Marutani Y, Yamauchi Y, Miyoshi A, Inoue K, Ikeda K-I, Mizutani M, Sugimoto Y (2014) Regulation of photochemical energy transfer accompanied by structural changes in thylakoid membranes of heat-stressed wheat. Int J Mol Sci 15:23042–23058
Maxwell K, Johnson GN (2000) Chlorophyll fluorescence—a practical guide. J Exp Bot 51:659–668
Minagawa J (2013) Dynamic reorganization of photosynthetic supercomplexes during environmental acclimation of photosynthesis. Front Plant Sci 4:article # 513. doi:10.3389/fpls.2013.00513
Mohanty P, Vani B, Prakash JSS (2002) Elevated temperature treatment induced alteration in thylakoid membrane organization and energy distribution between the two photosystems in Pisum sativum. Z Naturforsch 57c:836–842
Mohanty P, Nellaepalli S, Mishra S, Subramanyam R (2012) State shifts in photosynthetic organisms: tracking traits and techniques. In: Itoh S, Mohanty P, Guruprasad KN (eds) Photosynthesis: overviews on recent progress and future perspectives, chap 3. IK Publishers, New Delhi, pp 38–59
Munday JC, Govindjee (1969) Light induced changes in the fluorescence yield of chlorophyll a in vivo; III. The dip and the peak in the fluorescence transient of Chlorella pyrenoidosa. Biophys J 9:1–21
Munekage Y, Hashimoto M, Miyake C, Tomizawa K-I, Endo T, Tasaka M, Shikanai T (2004) Cyclic electron flow around photosystem I is essential for photosynthesis. Nature 429:558–579
Murata N (1969) Control of excitation transfer in photosynthesis. I. Light-induced change of chlorophyll a fluorescence in Porphyridium cruentum. Biochim Biophys Acta 172:242
Murata N, Allakhverdiev SI, Nishiyama Y (2012) The mechanism of photoinhibition in vivo: re-evaluation of the roles of catalase, α-tocopherol, non-photochemical quenching, and electron transport. Biochim Biophys Acta 1817:1127–1133
Mus F, Cournac L, Cardettini V, Caruana A, Peltier G (2005) Inhibitor studies on non-photochemical plastoquinone reduction and H2 photoproduction in Chlamydomonas reinhardtii. Biochim Biophys Acta 1708:322–332
Nagy G, Unnep R, Zsiros O, Tokutso R, Takizawa K, Porcar L, Moyet L, Petroutsos D, Garab G, Finazzi G, Minagawa J (2014) Chloroplast remodeling during state transitions in Chlamydomonas reinhardtii as revealed by noninvasive techniques in vivo. Proc Natl Acad Sci USA 111:5042–5047
Nellaepalli S, Mekala NR, Zsiros O, Mohanty P, Subramanyam R (2011) Moderate heat stress induces state transitions in Arabidopsis thaliana. Biochem Biophys Acta 1807:1177–1184
Nellaepalli S, Kodru S, Malavath T, Subramanyam R (2012) Anaerobiosis induced state transition: a non-photochemical reduction of PQ pool mediated by NDH in Arabidopsis thaliana. PLoS ONE 7:e49839. doi:10.1371/journal.pone.0049839
Nellaepalli S, Kodru S, Malavathi T, Subramanyam R (2013) Change in fast Chl a fluorescence transients, 2 dimensional protein profile and pigment protein interactions during state transitions in Arabidopsis thaliana. J Photochem Photobiol B 128:27–34
Nixon PJ (2000) Chlororespiration. Philos Trans R Soc B 355:1541–1547
Papageorgiou GC (1968) Fluorescence induction in Chlorella pyrenoidosa and Anacystis nidulans and its relation to photophosphorylation. PhD Thesis in Biophysics, University of Illinois at Urbana-Champaign. http://www.life.illinois.edu/govindjee/theses.html
Papageorgiou GC, Govindjee (1968a) Light-induced changes in the fluorescence yield of chlorophyll a in vivo. II. Chlorella pyrenoidosa. Biophys J 8:1316–1328
Papageorgiou GC, Govindjee (1968b) Light-induced changes in the fluorescence yield of chlorophyll a in vivo. I. Anacystis nidulans. Biophys J 8:1299–1315
Papageorgiou GC, Govindjee (eds) (2004) Chlorophyll a fluorescence: a signature of photosynthesis, vol 19. Advances in photosynthesis and respiration. Springer, Dordrecht
Papageorgiou GC, Govindjee (2011) Photosystem II fluorescence: slow changes—scaling from the past. J Photochem Photobiol B104:258–270
Papageorgiou GC, Stamatakis K (2004) Water and solute transport in cyanobacteria as probed by chlorophyll a fluorescence. In: Papageorgiou GC, Govindjee (eds) Chlorophyll a fluorescence: a signature of photosynthesis, vol 19. Advances in photosynthesis and respiration. Springer, Dordrecht, pp 663–678
Papageorgiou GC, Govindjee, Govindjee R, Mimuro M, Stamatakis K, Alygizaki-Zorba A, Murata N (1999) Light-induced and osmotically-induced changes in chlorophyll a fluorescence in two Synechocystic sp. PCC 6803 strains that differ in membrane lipid unsaturation. Photosynth Res 59:125–136
Papageorgiou GC, Tsimilli-Michael M, Stamatakis K (2007) The fast and slow kinetics of chlorophyll a fluorescence induction in plants, algae and cyanobacteria: a viewpoint. Photosynth Res 94:275–290
Peers G, Truong TB, Ostendorf E, Busch A, Elrad D, Grossman AR, Hippler M, Niyogi KK (2009) An ancient light-harvesting protein is critical for the regulation of algal photosynthesis. Nature 462:518–521
Peltier G, Cournac L (2002) Chlororespiration. Annu Rev Plant Biol 53:523–550
Pribil M, Pesaresi P, Hertle A, Barbato R, Leister D (2010) Role of plastid protein phosphatase TAP38 in LHCII dephosphorylation and thylakoid electron flow. PLoS Biol 8:e1000288. doi:10.1371/journal.pbio.1000288
Rebeille F, Gans P (1988) Interaction between chloroplasts and mitochondria in microalgae. Plant Physiol 88:973–975
Rintamaki E, Martinsuo P, Pursiheimo S, Aro EM (2000) Cooperative regulation of light-harvesting complex II phosphorylation via the plastoquinol and ferredoxin-thioredoxin system in chloroplasts. Proc Natl Acad Sci USA 97:11644–11649
Rochaix J-D (2014) Regulation and dynamics of the light-harvesting system. Annu Rev Plant Biol 65:287–309
Rochaix J-D, Goldschmidt-Clermont M, Merchant S (eds) (1998) The molecular biology of chloroplasts and mitochondria in Chlamydomonas, vol 7. Advances in photosynthesis and respiration. Springer, Dordrecht
Rumeau D, Peltier G, Cournac L (2007) Chlororespiration and cyclic electron flow around PSI during photosynthesis and plant stress response. Plant Cell Environ 30:1041–1051
Sane PV, Desai TS, Tatake VG, Govindjee (1984) Heat induced reversible increase in Photosystem I emission in algae, leaves and chloroplasts: spectra, activities, and relation to state changes. Photosynthetica 18:439–444
Schansker G, Tóth SZ, Strasser RJ (2006) Dark recovery of the Chl-a fluorescence transient (OJIP) after light adaptation: the qT component of non-photochemical quenching is related to an activated photosystem I acceptor side. Biochim Biophys Acta 1757:787–797
Schansker G, Tóth SZ, Kovács L, Holzwarth AR, Garab G (2011) Evidence for a fluorescence yield change driven by a light-induced conformational change within photosystem II during the fast chlorophyll a fluorescence rise. Biochim Biophys Acta 1807:1032–1043
Schansker G, Tóth SZ, Holzwarth AR, Garab G (2014) Chlorophyll a fluorescence: beyond the limits of the QA model. Photosynth Res 120:43–58
Schreiber U (2004) Pulse-amplitude-modulation (PAM) fluorometry and saturation pulse method: an overview. In: Papageorgiou GC, Govindjee (eds) Chlorophyll a fluorescence: a signature of photosynthesis, vol 19. Advances in photosynthesis and respiration. Springer, Dordrecht, pp 279–319
Shapiguzov A, Ingelsson B, Samol I, Andres C, Kessler F, Rochaix J-D, Vener AV, Goldschmidt-Clermont M (2010) The PPH1 phosphatase is specifically involved in LHCII dephosphorylation and state transitions in Arabidopsis. Proc Natl Acad Sci USA 107:4782–4787
Siedow JN, Bickett DM (1981) Structural features required for inhibition of cyanide-insensitive electron transfer by propyl gallate. Arch Biochem Biophys 207:32–39
Siedow JN, Umbach AL (2000) The mitochondrial cyanide-resistant oxidase: structural conservation amid regulatory diversity. Biochim Biophys Acta 1459:432–439
Sireesha K, Radharani B, Krishna PS, Sreedhar N, Subramanyam R, Mohanty P, Prakash JS (2012) RNA helicase, CrhR is indispensable for the energy redistribution and the regulation of photosystem stoichiometry at low temperature in Synechocystis sp. PCC6803. Biochim Biophys Acta 1817:1525–1536
Spalding MH, Critchley C, Govindjee, Ogren WL (1984) Influence of carbon dioxide concentration during growth on fluorescence induction characteristics of the green alga Chlamydomonas reinhardtii. Photosynth Res 5:169–176
Srivastava A, Strasser RJ, Govindjee (1995) Polyphasic rise of chlorophyll a fluorescence in herbicide-resistant D1 mutants of Chlamydomonas reinhardtii. Photosynth Res 43:131–141
Stamatakis K, Papageorgiou GC (2014) Delta pH-dependent non-photochemical quenching (qE) of excited chlorophylls in the photosystem II core complex of the freshwater cyanobacterium Synechococcus sp. PCC 7942. Plant Physiol Biochem 81:184–189
Stamatakis K, Tsimilli-Michael M, Papageorgiou GC (2007) Fluorescence induction in the phycobilisome-containing cyanobacterium Synechococcus sp. PCC 7942: analysis of the slow fluorescence transient. Biochim Biophys Acta 1767:766–772
Stern D, Witman G, Harris EH (eds) (2008) The Chlamydomonas Sourcebook, 3 vol set, 2nd edn. Academic Press, San Diego (also available as an e-book (Kindle edition))
Stirbet A, Govindjee (2011) On the relation between the Kautsky effect (chlorophyll a fluorescence induction) and Photosystem II: Basics and applications of the OJIP fluorescence transient. J Photochem Photobiol B 104:236–257
Stirbet A, Govindjee (2012) Chlorophyll a fluorescence induction: Understanding the thermal phase, the J-I-P rise. Photosynth Res 113:15–61
Stirbet A, Riznichenko GYu, Rubin AB, Govindjee (2014) Modeling chlorophyll a fluorescence transient: relation to photosynthesis. Biochemistry (Moscow) 79:291–323
Strasser RJ, Govindjee (1992) On the O–J–I–P fluorescence transients in leaves and D1 mutants of Chlamydomonas reinhardtii. In: Murata N (ed) Research in photosynthesis, vol 2. Kluwer, Dordrecht, pp 29–32
Strasser RJ, Srivastava A, Govindjee (1995) Polyphasic chlorophyll a fluorescence transient in plants and cyanobacteria. Photochem Photobiol 61:32–42
Strasser RJ, Tsimilli-Michael M, Srivastava A (2004) Analysis of the chlorophyll a fluorescence transient. In: Papageorgiou GC, Govindjee (eds) Chlorophyll a fluorescence: a signature of photosynthesis, vol 19. Advances in photosynthesis and respiration. Springer, Dordrecht, pp 321–362
Suggett J, Borowitzka MA, Prášil O (eds) (2010) Chlorophyll a fluorescence in aquatic sciences: methods and applications. Developments in applied phycology, vol 4, XVIII. Springer, Dordrecht
Takahashi H, Iwai M, Takahashi Y, Minagawa J (2006) Identification of the mobile light-harvesting complex II polypeptides for state transitions in Chlamydomonas reinhardtii. Proc Natl Acad Sci USA 103:477–482
Takahashi H, Clowez S, Wollman F-A, Vallon O, Rappaport F (2013) Cyclic electron flow is redox-controlled but independent of state transition. Nat Commun 4:Article number 1954. doi:10.1038/ncomms2954
Tóth SZ, Schansker G, Garab G, Strasser RJ (2007) Photosynthetic electron transport activity in heat-treated barley leaves: the role of internal alternative electron donors to photosystem II. Biochim Biophys Acta 1767:295–305
Tsimilli-Michael M, Stamatakis K, Papageorgiou GC (2009) Dark-to-light transition in Synechococcus sp. PCC 7942 cells studied by fluorescence kinetics assesses plastoquinone redox poise in the dark and photosystem II fluorescence component and dynamics during state 2 to state 1 transition. Photosynth Res 99:243–255
Ünlü C, Drop B, Croce R, van Amerongen H (2014) State transitions in Chlamydomonas reinhardtii strongly modulate the functional size of photosystem II but not of photosystem I. Proc Natl Acad Sci USA 111:3460–3465
Vanlerberghe GC (2013) Alternative oxidase: a mitochondrial respiratory pathway to maintain metabolic and signaling homeostasis during abiotic and biotic stress in plants. Int J Mol Sci 14:6805–6847
Velthuys BR (1981) Electron dependent competition between plastoquinone and inhibitors for the binding to PSII. FEBS Lett 126:277–281
Wollman F-A (2001) State transitions reveal the dynamics and flexibility of the photosynthetic apparatus. EMBO J 20:3623–3630
Wollman F-A, Bulté L (1990) Toward an understanding of the physiological role of state transitions. In: Hall DO, Grassi G (eds) Photoconversion processes for energy and chemicals. Elsevier, London, pp 198–207
Wollman F-A, Delepelaire P (1984) Correlation between changes in light energy distribution and changes in thylakoid membrane polypeptide phosphorylation in Chlamydomonas reinhardtii. J Cell Biol 98:1–7
Wollman F-A, Lemaire C (1988) Studies on kinase-controlled state transitions in Photosystem II and b6f mutants from Chlamydomonas reinhardtii which lack quinone-binding proteins. Biochim Biophys Acta 933:85–94
Yoshida K, Terashima I, Noguchi K (2006) Distinct roles of the cytochrome pathway and alternative oxidase in leaf photosynthesis. Plant Cell Physiol 47:22–31
Zhang L-T, Zhang Z-S, Gao H-Y, Meng X-L, Yang C, Liu J-G, Meng Q-W (2012) The mitochondrial alternative oxidase pathway protects the photosynthetic apparatus against photodamage in Rumex K-1 leaves. BMC Plant Biol 12:40. http://www.biomedcentral.com/1471-2229/12/40
Zivcak M. Brestic M, Kalaji HM, Govindjee (2014) Photosynthetic responses of sun- and shade-grown barley leaves to high light: Is the lower connectivity in shade leaves associated with protection against excess of light. Photosynth Res 119:339–354
Zito F, Finazzi G, Delosme R, Nitschke W, Picot D, Wollman FA (1999) The Qo site of cytochrome b6f complexes controls the activation of the LHCII kinase. EMBO J 18:2961–2969
Acknowledgments
Rajagopal Subramanyam was supported by the Department of Biotechnology (BT/PR15132/BRB/10/909/2011) and the Council of Scientific and Industrial Research (38(1381)/14/EMR-II), India, and Sireesha Kodru by the Department of Biotechnology for a research associate fellowship (DBT-RA), India. Govindjee was supported by the US Fulbright foundation during his stay at the University of Hyderabad, India. We are grateful to Reto J. Strasser and to the late Prasanna Mohanty for providing the Handy PEA instrument used in our studies. We thank Vandana Chakravartty of the University of Illinois at Urbana-Champaign for her valuable comments during the preparation of this manuscript.
Author information
Authors and Affiliations
Corresponding authors
Additional information
In earlier publications, Sireesha Kodru and Sreedhar Nellaepalli have used their names as Kodru Sireesha and Nellaepalli Sreedhar, respectively.
Rajagopal Subramanyam—in earlier publications, has used his name as Subramanyam Rajagopal.
Electronic supplementary material
Below is the link to the electronic supplementary material.
Rights and permissions
About this article

Cite this article
Kodru, S., Malavath, T., Devadasu, E. et al. The slow S to M rise of chlorophyll a fluorescence reflects transition from state 2 to state 1 in the green alga Chlamydomonas reinhardtii . Photosynth Res 125, 219–231 (2015). https://doi.org/10.1007/s11120-015-0084-2
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11120-015-0084-2