
Abstract
The human tissue plasminogen activator (t-PA) is a thrombolytic protein that plays a key role in fibrinolysis by converting the plasminogen into plasmin which degrades fibrin clots in blood vessels. The t-PA gene encoding the human tissue plasminogen activator was cloned and expressed under transcriptional control of single and multiple rooting loci promoter (rolD) promoters in hairy roots derived from Oriental melon. t-PA gene integration in the genome of hairy roots was confirmed by PCR. Increased transcript and protein expression levels was observed in the transgenic hairy roots expressing t-PA under the control of single and dual rolD promoters, respectively. Enzyme-linked immunosorbent assay revealed the highest levels of the recombinant t-PA accumulation in transgenic hairy roots carrying the t-PA transgene under the control of single and dual rolD (0.17–0.15 μg/mg) promoters as compared to triple and quadruple rolD (0.07–0.03 μg/mg) promoters. Fibrinolytic activity indicated the proper function of the expressed recombinant t-PA in fibrin lysis. These results demonstrated that hairy roots could serve as alternatives to transgenic plants for expression of mammalian genes.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
The human tissue-plasminogen activator (t-PA) is a single-chain polypeptide consisting of 527 amino acids residues and 17 disulfide bonds. The tissue-plasminogen activator is comprised of five distinct structural domains such as: a finger domain, a growth factor-like subdomain, two kringle domains, and finally a serine protease domain (Pennica et al. 1983). t-PA plays a crucial role in the conversion of zymogen plasminogen to plasmin, a serine protease of broad specificity that degrades the fibrin network of the thrombus (Van Zonneveld et al. 1986). Both finger and kringle 2 domains bind specifically to the fibrin clot and facilitate t-PA protein activation. Next to the kringle 2 domain is the serine protease domain in which the catalytic site is located at the C terminus of the protein. This domain is responsible for plasminogen conversion into plasmin and eventually leading to fibrin homeostasis. The correct folding of t-PA requires correct pairing of multiple disulfide bridges in the molecule (Pennica et al. 1983).
The clinical uses of t-PA have shown it to be an effective thrombolytic agent with minimal effects on circulating plasminogen and fibrinogen levels (Collen et al. 1986). The Bowes melanoma cells were the primary source of t-PA production for therapeutic purposes (Griffiths and Electricwala 1987). Expression of recombinant t-PA has also been reported in diverse organisms, such as Escherichia coli (Kim et al. 2005; Manosroi et al. 2001; Pennica et al. 1983; Qiu et al. 1998), Aspergillus nidulans (Upshall et al. 1987), baculovirus-infected insect cells (Jarvis and Summers 1989), goat milk (Ebert et al. 1991), mammalian CHO cells (Cartwright 1992), yeast (Keivan et al. 2010), and Leishmania tarentolae (Nazari and Davoudi 2010). As an alternative to these systems recently, the successful use of tobacco plants for the production of t-PA has been reported (Hahn et al. 2009).
Many therapeutic proteins, valuable secondary metabolites and functional recombinant proteins have been produced using hairy roots cultures or transgenic plants, such as thymosin alpha 1 (Cui et al. 2010), human secreted alkaline phosphatase (Gaume et al. 2003), fibrinolytic enzyme (Ko et al. 2009), elevated ginsenoside (Liang et al. 2009), cry1Ac (Nandeshwar et al. 2009), monoclonal antibodies (Sharp and Doran 2001), synthetic heat-labile enterotoxin B subunit and hemagglutinin–neuraminidase-neutralizing epitope fusion protein (Sim et al. 2009), ribosome-inactivating proteins (Thorup et al. 1994), human acetylcholinesterase (Woods et al. 2008), hepatitis B surface antigen (Unni and Soniya 2010), and human granulocyte-macrophage colony stimulating factor (Wang et al. 2009). These studies demonstrate that hairy roots and transgenic plants can be a better alternative to mammalian cell cultures and microbial fermentation for the production of recombinant proteins.
Plant-based production of therapeutic proteins is appealing due to its low cost, ease of management, and presence of post-translational modification machinery that can engineer more human-like modifications as compared to mammalian cell cultures and microbial fermentation. However, restrictions and regulations are imposed on the transgenic plant approach because of the potential for gene transfer to wild species through cross-pollination with closely-related species (Streatfield 2005). In this context, production of therapeutic proteins using plant cell or organ cultures such as hairy root cultures may prove to be advantageous as compared to the other systems. While several plant cell lines are available for use, more organized hairy root cultures may present additional benefits, e.g., genetic and biochemical stability and faster growth rates (Sivakumar 2006; Srivastava and Srivastava 2007).
Oriental melons (Cucumis melo), which originated in Africa and Southwest Asia (Silberstein et al. 1999), are grown in a variety of climates worldwide. Melons (C. melo), rich in nutrients (carbohydrate, organic acids, minerals, and vitamins) is widely cultivated as a commercially profitable crop in Korea. We previously reported the development of a highly efficient system for the Agrobacterium rhizogenes-mediated transformation of Oriental melons by overexpression of the gfp-gus fusion gene (Pak et al. 2009). These results strongly showed the possibility of producing therapeutic proteins in the hairy roots of Oriental melons.
In this study, we have demonstrated the feasibility of producing the human t-PA in transgenic hairy roots derived from Oriental melon (C. melo) plants wherein the t-PA gene is driven by the rolD promoter following A. rhizogenes-mediated transformation. Hairy root cultures were screened for the t-PA expression and accumulation levels and its biochemical properties were subsequently studied.
Materials and Methods
Construction of Expression Vectors
Four expression cassettes were generated containing t-PA gene (1,584 bp; GenBank accession no. A07197) driven under the transcriptional control of single (1×), dual (2×), triple (3×), and quadruple (4×) roD promoter fragments. The rolD promoter fragment (450 bp) was generated by polymerase chain reaction (PCR) amplification using flanking primers containing EcoR I/Stu I and Hpa I/Mlu I sites and cloned in to pBluescript vector (Stratagene, USA). The rolD fragment was isolated by EcoR I/Mlu I digestion and inserted into the same sites upstream of the t-PA gene in the binary p221a vector (Hahn et al. 2009) and named as p221a-rolD1×. The dual rolD (2×) expression vector was generated by ligating the rolD fragment digested with EcoR I/Hpa I in the pBluescript vector and rolD PCR product digested with Stu I/Mlu I and inserted into the same sites of p221a vector and named as p221a-rolD2×. Expression cassettes with triple (3×) and quadruple (4×) rolD fragments were cloned into p221a vector separately, and were named as p221a-rolD3× and p221a-rolD4×, respectively. A Kpn I/Hind III t-PA gene fragment containing the signal sequence of the alfalfa glucose-regulated endoplasmic reticular protein was cloned in to plant binary plasmid p221a, which included a tobacco etch virus leader sequence. The vector (p221a) had npt II and bar as two selectable markers driven under the control of CaMV 35S promoter and 35S terminator and Ti7 terminator. Finally, the transfer of the subcloned t-PA and rolD promoter fragments into the vector was confirmed by DNA sequencing.
Induction of Hairy Roots
The Agrobacterium-mediated transformation of melon cotyledons was performed as follows. Oriental melon seeds from (C. melo L. cv. Geumssaragi-euncheon) were sterilized, germinated and grown on Murashige and Skoog (MS) medium (Murashige and Skoog 1962). The cotyledons of 1-week-old seedlings (30–40 cotyledons) infected with and without A. rhizogenes (K599) for 30 min. After removing the excessive bacteria culture with filter paper, the transformed explants were transferred to MS medium and kept in the dark for 2 days. The infected cotyledons were washed with sterile water three times. After removing the excess water with filter paper, the putative transformed explants were transferred to MS medium containing kanamycin (300 μg/ml) and carbenicilin (500 μg/ml) in order to induce and select putative transformed hairy roots. The induced hairy roots were subcultured to the same MS medium at biweekly intervals for 4 weeks and were sequentially transferred to MS medium containing kanamycin (300 μg/ml) and carbenicilin (100 μg/ml) for maintenance and further analysis.
PCR and RT-PCR Analysis
Genomic DNA was isolated from putative transgenic hairy roots and the PCR amplification was carried out using the gene specific primers (5′-TCTTACCAAGT-GATCTGCAGAG-3′) and (5′-TCACGGTCGCAT-GTTGTCAC-3′). PCR was performed in an HP thermacycler under the following conditions: 95°C for 30 s, 60°C for 30 s, and 72°C for 2 min for a total of 30 cycles. PCR products were analyzed on 1% gel. Total RNA was isolated from 150 mg of hairy roots using the TRIzol reagent according to the manufacturer instructions (Invitrogen). Reverse transcription polymerase chain reaction (RT-PCR) analysis was performed with a First-Strand cDNA synthesis kit (Takara) following the instruction manual in the kit. Specific primers were used for the amplification of t-PA (forward: 5′-TCTTACCAAGT-GATCTGCAGAG-3′ and reverse: 5′-TCACGGTCGCAT-GTTGTCAC-3′). The amplified PCR products were electrophoresed on a 1% agarose gel containing ethidium bromide.
Enzyme-Linked Immunosorbent Assay
The amount of recombinant t-PA in total soluble proteins of hairy root extracts was detected using a double-sandwich enzyme-linked immunosorbent assay (ELISA). Briefly, a 96-well ELISA plate was coated with polyclonal antibody (raised in sheep) to human t-PA. The microwell strips were washed twice with approximately 300 μl of wash buffer per well.
Hairy root samples (100 mg) were homogenized in 500 μl of buffer containing 1× phosphate-buffered saline (PBS)-0.05% Tween 20 and 1× protease inhibitor cocktail (Sigma). The insoluble material was removed by centrifugation at 12,000 rpm for 20 min. The protein concentrations were determined by using a Bradford Protein Assay Reagent kit (Bio-Rad). Five hundred nanograms of total soluble protein was added and incubated for 2 h at 37°C. The wells were washed three times with PBS and then horseradish peroxidase (HRP) conjugated anti t-PA monoclonal antiserum (1:100 dilution) was then added to each well and incubated for 2 h at 37°C. After washing the wells as before, 100 μl of tetramethylbenzidine was added, and the plate was incubated for 15 min. The enzyme reaction was stopped by quickly pipetting 100 μl of 1 M phosphoric acid in to each well. The absorbance was read by a microplate reader at 450 nm. The t-PA standard was used as a positive control.
Western Blot Analysis
Hairy root samples (100 mg) were homogenized in 500 μl of a buffer solution containing 50 mM Tris–HCl (pH 7.5), 400 mM NaCl, 1 mM EDTA, 20 mM DTT, 5% glycerol, 1% Triton X-100, and 1× protease inhibitor cocktail (Sigma). The insoluble material was removed by centrifugation at 12,000 rpm for 20 min. The protein concentrations were determined by using Bradford Protein Assay Reagent kit (Bio-Rad). The proteins (100 μg) were separated in 8% SDS polyacrylamide gels and then electrophoretically transferred to Hybond-enhanced chemiluminescence (ECL) nitrocellulose membrane (GE Healthcare). Transferred membrane was blocked with 5% nonfat milk powder in Tween-/Tris-buffered saline to prevent the nonspecific antibody binding. The membrane was then probed with polyclonal t-PA antibody at a dilution of 1:1,000. After being washed three times, the blotted membrane was probed with HRP-conjugated secondary antibody (1:5,000) for 2 h. Proteins were detected using the ECL chemiluminescence reagent as recommended by the manufacturer’s protocol.
Fibrinolytic Activity and Inhibition of the Fibrinolytic Activity of Recombinant t-PA Using PAI-1
Fibrinolytic activity was performed with modified method of Astrup and Mullertz (1952). The fibrinogen solution (10 ml of 0.8% bovine fibrinogen in 0.2 M borate buffer, pH 7.8) was poured into a Petri dish (2 × 9 cm) and then clotted by the addition of 0.5 ml of 10 NIH U/ml thrombin in 0.9% saline. The clot was allowed to stand for 30 min at room temperature. Ten micrograms of crude hairy root extracts were spotted carefully on the fibrin-coated plate and the plates were incubated for 17 h at 37°C. The fibrinolytic activity was carried out by measuring two perpendicular diameters of the lysed zone. For the measurement of inhibition of the fibrinolytic activity of recombinant t-PA using plasminogen activator inhibitor (PAI-1), 5 μg of crude hairy root extracts with/without a mixture of PAI-1 (1 μg) was added to the fibrin-coated plates and incubated at room temperature for 20 min. The samples were then spotted on the fibrin-coated plate and were incubated for 20 h at 37°C. In this fibrin plate method, a clear transparent region is observed where fibrin is hydrolyzed.
Results
Construction of Expression Vector and Induction of Transgenic Hairy Roots
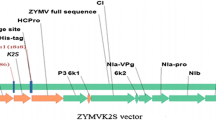
To study the comparative expression of t-PA gene under the control of rolD (1×), (2×), (3×), and (4×) promoters, four expression cassettes were used as shown in Fig. 1a. The subcloning into the binary vector (p221a) and the size of the rolD promoters was confirmed by EcoR I/Mlu I digestion which yielded expected rolD (1×; 450 bp), (2×; 900 bp), (3×; 1,350 bp), and (4×; 1,800 bp) fragments, respectively (Fig. 1b). Thirty putative transgenic hairy roots were induced by A. rhizogenes K599 harboring plant expression vectors and 20 transgenic hairy roots were selected for subsequent assays.

Schematic representation of the expression cassettes and electrophoretic analysis of binary vectors p221a-rolD1×/2×/3×/4×-t-PA. a The t-PA gene fusion constructs in p221a plasmid is under the control of rolD promoters (rolD 1–4×). Tobacco etch virus (TEV), signal sequence of the alfalfa glucose-regulated endoplasmic reticular protein (SS), cauliflower mosaic virus promoter (CaMV 35S), nopalin synthase gene (npt II); bar gene for selection with PPT (phosphinothricin), terminator of nopalin synthase gene (Tnos). b Restriction fragments of p221a-rolD1×/2×/3×/4×: 2–3 EcoR I and Mlu I digest of p221a-rolD1× plasmid; 4–5 EcoR I and Mlu I digest of p221a-rolD2×; 6–7 EcoR I and Mlu I digest of p221a-rolD3×; 8–9 EcoR I and Mlu I digest of p221a-rolD4×, respectively
Molecular Analysis of the t-PA Gene in Transgenic Hairy Roots
The presence of t-PA gene in the genomic DNA of different transgenic hairy roots was confirmed by PCR amplification. The expected amplification product of 1,584 bp was detected from five putative transgenic hairy roots expressing rolD (1×), (2×), (3×), and (4×) promoters, respectively (Fig. 2a). The plasmid p221a-t-PA chosen as a positive control was also found to be positive for PCR, while no amplification was observed in the untransformed control (Fig. 2a, lane 3).

Molecular analysis for the detection of the t-PA gene. a PCR analysis of the integration of the t-PA gene into transgenic hairy roots. 4–13 and 14–23 transformed hairy root genomic DNAs exhibiting amplified fragment (1,584 bp). 1 DNA-size markers; 2 p221a-rolD1x-t-PA binary vector; 3 the genomic DNA of the hairy root induced by K599 harboring no binary vector as a negative control. b RT-PCR from selected transgenic hairy roots showing t-PA gene expression (4–11). 1 DNA-size markers; 2 p221a-rolD1×-t-PA plasmid served as the positive control; 3 RNA of the hairy root induced by K599 harboring no binary vector as a negative control
The mRNA expression of t-PA gene in transgenic hairy roots was confirmed by RT-PCR analysis. The amplification products correspond to specific transcript of the expected size of the transgene was observed. No amplification was observed in the untransformed controls used as a negative control. The transgenic hairy roots expressing rolD (1×; Fig. 2b, lanes 4 and 5) and rolD (2×; Fig. 2b, lanes 6 and 7) promoters showed elevated t-PA expression levels when compared to the transgenic hairy roots expressing rolD (3×; Fig. 2b, lanes 8 and 9), and rolD (4×; Fig. 2b, lanes 10 and 11) promoters which showed reduced expression levels (Fig. 2b).
Fibrinolysis of Recombinant t-PA
Fibrinolysis assay performed to determine the fibrinolytic activity of the recombinant t-PA from the transgenic hairy roots. Fibrinolytic activity was apparent in all the transgenic hairy root samples expressing rolD promoters (Fig. 3). The transgenic hairy root samples expressing rolD (1×) and (2×) promoters showed 0.3–0.25 cm2 average fibrinolytic activity whereas, the hairy roots samples expressing rolD (3×) and (4×) promoters exhibited 0.1–0.07 cm2 average fibrinolytic activity of recombinant t-PA.

Fibrinolysis of recombinant t-PA in selected transgenic hairy roots. Ten micrograms of total root proteins from transgenic hairy roots was spotted onto fibrin plate and zone of clearance was measured. Vertical point plot fibrinolytic activity of crude extracts and the selected transgenic hairy roots. Triangle average fibrinolytic activity of t-PA followed by rolD promoter number (rolD1×-4×), respectively
Enzyme-Linked Immunosorbent Assay
The immunoactivity of the recombinant t-PA protein extracted from the transgenic hairy roots was analyzed by ELISA. The crude extracts of transgenic hairy roots expressing t-PA under the control of rolD promoters exhibited detectable amount of recombinant t-PA (Fig. 4a). The enhanced t-PA levels was exhibited by transgenic hairy roots expressing rolD (1×) and rolD (2×) promoters when compared to transformants expressing rolD (3×) and rolD (4×) promoters. The average expression level of recombinant t-PA from different transgenic hairy roots ranged from 0.03 to 0.17 μg/mg.

T-PA levels and western blot analysis in selected transgenic hairy roots. a ELISA sandwich assay: TSP from hairy roots was used for ELISA. K599 is a negative control, induced hairy root by K599 harboring no binary vector. Triangle average amount of t-PA followed by rolD promoter number (rolD1×–4×), respectively. b Western blot analysis: TSP from hairy roots was used for immune blotting. Sheep anti-t-PA serum and rabbit anti-sheep horseradish peroxidase conjugate were used as primary and secondary antibodies, respectively. 1 Recombinant protein from animal cells, 2 negative control, 3–6 selected transgenic hairy roots. Arrowhead points to the recombinant t-PA
Western Blot Analysis
Western blot analysis was performed to evaluate the expression of the recombinant t-PA in the transgenic hairy roots. The results revealed that the antibodies raised against the recombinant t-PA cross-reacted with total soluble protein (TSP) displaying a distinct band with the expected molecular weight of 59 kDa (Fig. 4b). The t-PA antibody did not recognize any proteins extracted from untransformed control hairy roots (Fig. 4b, lane 2). The protein expression levels was elevated in the transgenic hairy roots expressing rolD (1×) and (2×) (Fig. 4b, lanes 3 and 4) promoters as compared to the hairy roots expressing rolD (3×) and (4×; Fig. 4b, lanes 5 and 6) promoters which exhibited weak signals.
Inhibition of Fibrinolytic Activity of Recombinant t-PA Using PAI-1
In order to study the inhibition of the fibrinolytic activity of recombinant t-PA in the transgenic hairy roots, crudes extracts with PAI-1 were spotted on a fibrin plate for phenotype observation. The results revealed that, in the absence of PAI-1, crude transgenic hairy root extracts readily digested the fibrin clot (Fig. 5, lanes 3–6), whereas no clear zone was observed in untransformed hairy root extract (Fig. 5, lane 1). In the presence of PAI-1, none of the samples exhibited a clear lysed zone on the fibrin plates (Fig. 5, lanes 7–12).

Inhibition of fibrinolytic activity of recombinant t-PA using PAI-1. Total root proteins from transgenic hairy roots was spotted onto a plate containing fibrin clot and incubated at 37°C overnight. In the absence of PAI-1 (1 μg), 1 is the hairy root induced by K599 harboring no binary vector, 2 is the recombinant t-PA protein (0.2 ng) from animal cells. 3–6 Transformed hairy roots (rolD1×–4×). In the presence of PAI-1 (1 μg), 7 is the hairy root induced by K599 harboring no binary vector; 8 is the recombinant t-PA protein from animal cells; 9 and 12 transformed hairy roots (rolD1×–4×)
Discussion
Hairy root cultures, induced by A. rhizogenes infection are extensively used in the field of genetic engineering due to their ease of propagation and growth in confined environments. One of the aspects of transformed root cultures that have attracted intense commercial interest is their potential for protein production (Shanks and Morgan 1999). Therefore, we investigated the expression and production of t-PA in hairy roots using Agrobacterium-mediated transformation. Many therapeutic proteins have been produced using hairy roots cultures, such as monoclonal antibodies (Sharp and Doran 2001), ribosome-inactivating proteins (Thorup et al. 1994), human secreted alkaline phosphatase (Gaume et al. 2003), and human acetylcholinesterase (Woods et al. 2008). These studies demonstrate that hairy roots can be a better alternative to whole plants for producing pharmacologically functional animal proteins.
The promoters of the rooting loci (rol) genes found in the Ri (root-inducing) plasmid of A. rhizogenes are popular targets of study because of the root-directed transformation and expression of their gene products. Specifically, the rolD promoter has been analyzed by a series of promoter deletion: GUS fusion constructs and has been found useful. In this experiment, single (1×), dual (2×), triple (3×), and quadruple (4×) rolD promoters were analyzed for their ability to drive maximal expression of t-PA gene. The t-PA gene integration in to the genome, mRNA, and protein expression levels were confirmed by PCR, RT-PCR, and western blotting. The transcript and protein expression levels of recombinant t-PA were found to be higher in the transgenic hairy roots expressing rolD (1×) and rolD (2×) promoters when compared to hairy roots expressing rolD (3×) and rolD (4×) promoters. Further, the yield of recombinant t-PA from transgenic hairy roots expressing rolD (1×; 0.17 μg/mg) and rolD (2×; 0.15 μg/mg) promoters were increased when compared to hairy roots expressing rolD (3×; 0.07 μg/mg) and rolD (4×; 0.03 μg/mg) promoters. Similarly, the fibrinolytic activity exhibited by the recombinant t-PA was in correlation with the protein expression studies. Overall, our findings indicate that single and dual rolD promoter expression provided the highest transgene and protein expression in the transgenic hairy roots. Our results were consistent with the earlier reports with rolD (1×) promoter directing high levels of transgene expression five times higher than the CaMV 35S promoter (Leach and Aoyagi 1991) and 30-fold higher expression of GUS in roots of transgenic tobacco plants driven by rolD (2×) promoter (Elmayan and Tepfer 1995). However, the t-PA expression levels in the transgenic hairy roots expressing rolD (3×) and (4×) promoters were greatly reduced. The possible reason could be the gene silencing at the transcriptional level due to repeated use of the rolD promoter. It has been reported that homologue sequence, more than 90 bp within two promoters, often leads to gene silencing in transgenic plants (Parry et al. 1994).
In conclusion, previously, we reported the generation of transgenic tobacco plants that accumulate recombinant human t-PA driven by CaMV 35S promoter (Hahn et al. 2009). Here, we demonstrate that the t-PA can be efficiently produced in hairy roots derived from Oriental melon driven under rolD promoters (1–4×) which was 12 times higher than the previous levels in the transgenic tobacco plants (Hahn et al. 2009). The transgenic hairy roots expressing rolD (1×) and rolD (2×) constructs exhibited elevated t-PA levels when compared to hairy roots expressing rolD (3×) and rolD (4×) constructs. Therefore, it is desirable to minimize the repeated use of a single promoter that may cause transcriptional gene silencing leading to the reduced accumulation of recombinant proteins.
References
Astrup T, Mullertz S (1952) The fibrin plate method for estimating of fibrinolytic activity. Arch Biochem Biophys 40:346–351
Cartwright T (1992) Production of tPA from animal cell culture. In: Spier RE, Griffiths JB (eds) Anim cell biotechnol London. Academic, UK, pp 218–245
Collen D, Bounameaux H, DeCock F et al (1986) Analysis of coagulation and fibrinolysis during intravenous infusion of recombinant human tissue-type plasminogen activator in patients with acute myocardial infarction. Circulation 73:511–517
Cui L, Chen Y, Shen G, Zhao L, Tang K (2010) Expression of bioactive thymosin alpha 1 (Tα1) in marker-free transgenic lettuce (Lactuca sativa). Plant Mol Biol Rep. doi:10.1007/s11105-010-0246-5
Ebert KM, Selgrath JP, DiTullio P et al (1991) Transgenic production of a variant of human tissue-type plasminogen activator in goat milk: generation of transgenic goats and analysis of expression. Biotechnology 9:835–838
Elmayan T, Tepfer M (1995) Evaluation in tobacco of the organ specificity and strength of the rolD promoter, domain A of the 35S promoter and the 35S2 promoter. Trans Res 4:388–396
Gaume A, Komarnytsky S, Borisjuk N et al (2003) Rhizosecretion of recombinant proteins from plant hairy roots. Plant Cell Rep 21:1188–1193
Griffiths JB, Electricwala A (1987) Production of tissue plasminogen activators from animal cells. Adv Biochem Eng Biotechnol 34:147–166
Hahn BS, Sim JS, Kim HM et al (2009) Expression and characterization of human tissue-plasminogen activator in transgenic tobacco plants. Plant Mol Biol Rep 27:209–216
Jarvis DL, Summers MD (1989) Glycosylation and secretion of human tissue plasminogen activator in recombinant baculovirus-infected insect cells. Mol Cell Biol 9:214–223
Keivan MA, Vahid K, Davami F et al (2010) Cloning and expression of functional full-length human tissue plasminogen activator in Pichia pastoris. Appl Biochem Biotechnol 162:2037–2048
Kim JY, Fogarty EA, Lu FJ, Zhu H, Wheelock GD, Henderson LA et al (2005) Twin-arginine translocation of active human tissue plasminogen activator in Escherichia coli. Appl Environ Microbiol 71:8451–8459
Ko SM, Yoo BH, Lim JM, Oh KH, Liu JI, Kim SW, Liu JR, Choi KS, Yoon ES (2009) Production of fibrinolytic enzyme in plastid-transformed tobacco plants. Plant Mol Biol Rep 27:448–453
Leach F, Aoyagi K (1991) Promoter analysis of the highly expressed rolC and rolD root-inducing genes of Agrobacterium rhizogenes: enhancer and tissue-specific DNA determinants are dissociated. Plant Sci 79:69–76
Liang Y, Zhao S, Zhang X (2009) Antisense suppression of cycloartenol synthase results in elevated ginsenoside levels in Panax ginseng hairy roots. Plant Mol Biol Rep 27:298–304
Manosroi J, Tayapiwatana C, Gotz F, Werner RG, Manosroi A (2001) Secretion of active recombinant human tissue plasminogen activator derivatives in Escherichia coli. Appl Environ Microbiol 67:2657–2664
Murashige T, Skoog F (1962) A revised medium for rapid growth and bioassays with tobacco tissue cultures. Physiol Plant 15:473–497
Nandeshwar SB, Moghe S, Chakrabarty PK, Deshattiwar MK, Kranthi K, Anandkumar P, Mayee CD, Khadi BM (2009) Agrobacterium-mediated transformation of cry1Ac gene into shoot-tip meristem of diploid cotton Gossypium arboreum cv. RG8 and regeneration of transgenic plants. Plant Mol Biol Rep 27:549–557
Nazari R, Davoudi N (2010) Cloning and expression of truncated form of tissue plasminogen activator in Leishmania tarentolae. Biotechnol Lett. doi:10.1007/s10529-010-0470-y
Pak HK, Sim JS, Rhee Y, Ko HR, Ha SH, Yoon MS, Kang CH, Lee S, Kim YH, Hahn BS (2009) Hairy roots induction in Oriental melon (Cucumis melo) by A. rhizogenes and production of the root-knot nematode. Plant Cell Tiss Organ Cult 98:219–228
Parry SK, Sharma SB, Terzaghi EA (1994) Construction of a bidirectional promoter probe vector and its use in analyzing Nod gene-expression in Rhizobium loti. Gene 150:105–109
Pennica D, Holmes WE, Kohr WJ, Harkins RN, Vehar GA, Ward CA et al (1983) Cloning and expression of human tissue-type plasminogen activator cDNA in E. coli. Nature 301:214–221
Qiu J, Swartz JR, Georgiou G (1998) Expression of active human tissue-type plasminogen activator in Escherichia coli. Appl Environ Microbiol 64:4891–4896
Shanks JV, Morgan J (1999) Plant “hairy root” culture. Curr Opin Biotechnol 10:151–155
Sharp JM, Doran PM (2001) Strategies for enhancing monoclonal antibody accumulation in plant cell and organ cultures. Biotechnol Prog 17:979–992
Silberstein L, Kovalski I, Huang R, Anagostou K, Kyle Jahn M, Perl-Treves R (1999) Molecular variation in melon (Cucumis melo L.) as revealed by RFLP and RAPD markers. Sci Hort 79:101–111
Sim JS, Pak HK, Kim DS, Lee SB, Kim YH, Hahn BS (2009) Expression and characterization of synthetic heat-labile enterotoxin B subunit and hemagglutinin–neuraminidase-neutralizing epitope fusion protein in Escherichia coli and tobacco chloroplasts. Plant Mol Biol Rep 27:388–399
Sivakumar G (2006) Bioreactor technology: a novel industrial tool for high-tech production of bioactive molecules and biopharmaceuticals from plant roots. Biotechnol J 1:1419–1427
Srivastava S, Srivastava A (2007) Hairy root culture for mass-production of high-value secondary metabolites. Crit Rev Biotech 27:29–43
Streatfield SJ (2005) Regulatory issues for plant-made pharmaceuticals and vaccines. Exp Rev Vacc 4:591–601
Thorup JE, McDonald KA, Jackman AP et al (1994) Ribosome-inactivating protein production from Trichosanthes kirilowii plant cell cultures. Biotechnol Prog 10:345–352
Unni SC, Soniya EV (2010) Transgenic Cucumis sativus expressing the hepatitis B surface antigen. Plant Mol Biol Rep 28:627–634
Upshall A, Kumar AA, Bailey MC, Parker MD, Favreau MA (1987) Secretion of active human tissue plasminogen activator from the filamentous fungus Aspergillus nidulans. Biotechnology 5:1301–1304
Van Zonneveld AJ, Chang GT, van den Berg J et al (1986) Quantification of tissue-type plasminogen activator (t-PA) mRNA in human endothelial-cell cultures by hybridization with a t-PA cDNA probe. Biochem J 235:385–390
Wang B, Jiang YN, Yao LM, Wu TL (2009) Improved production of the human granulocyte-macrophage colony stimulating factor in transgenic Arabidopsis thaliana seeds using a dual sorting signal peptide. Plant Mol Biol Rep 27:462–468
Woods RR, Geyer BC, Mor TS (2008) Hairy root organ cultures for the production of human acetylcholinesterase. BMC Biotechnol 8:95
Acknowledgments
We thank Ms. Kyung-Il Lee (NAAS) for technical assistance. This project (PJ006652) was supported by grant (to BSH) from the National Academy of Agricultural Science, RDA, Korea. This study was supported by 2011 Postdoctoral Fellowship Program (to HA) of National Academy of Agricultural Science, RDA, Korea.
Author information
Authors and Affiliations
Corresponding author
Additional information
Sol Kang and Hemavathi Ajjappala equally contributed to this work.
Rights and permissions
About this article
Cite this article
Kang, S., Ajjappala, H., Seo, HH. et al. Expression of the Human Tissue-Plasminogen Activator in Hairy Roots of Oriental Melon (Cucumis melo). Plant Mol Biol Rep 29, 919–926 (2011). https://doi.org/10.1007/s11105-011-0303-8
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11105-011-0303-8