
Abstract
Aims
Corn and soybean crops are often grown in rotation, requiring lower nitrogen (N) inputs than continuous corn. However, soil organic carbon (C) may be declining in corn-soybean systems despite sustained residue inputs. We asked whether corn-soybean rotations increase decomposition of litter and soil C as compared with continuous corn.
Methods
We incubated soils from a long-term field experiment including continuous corn and both phases of the corn-soybean rotation. Soils were amended with corn litter, soybean litter, or no litter. We measured natural abundance C stable isotopes (δ13C values) in respiration and microbial biomass to partition C sources.
Results
Addition of soybean litter increased microbial biomass while corn did not. However, corn litter addition consistently increased (i.e., primed) soil C decomposition while soybean litter did not. Soils most recently planted to corn following soybeans had the greatest soil C decomposition and N mineralization irrespective of litter addition, and they decomposed corn litter faster and had a faster priming response than the other treatments.
Conclusions
Our data support the hypothesis that alternating inputs of N-rich soybean litter and relatively N-poor corn litter could enhance litter and SOC decomposition by driving microbial growth following the soybean phase and stimulating priming following the corn phase. Increased decomposition and N mineralization from litter and SOC in corn-soybean rotations may contribute to the soybean N credit but could also contribute to longer-term soil C and N declines, consistent with field data.
Similar content being viewed by others


Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
Corn (Zea mays L.) and soybean (Glycine max (L.) Merr.) crops grown in rotation are the dominant land cover and source of primary productivity in the Corn Belt of the midwestern United States, where a smaller area is under continuous corn cultivation (Prince et al. 2001; Yu et al. 2018). Impacts of these cropping systems on soil carbon (C) and nitrogen (N) cycling have received intensive study. Major soil organic C losses followed the conversion of historical grasslands to croplands, spurring interest in management to promote C accrual (Huggins et al. 1998; Russell et al. 2005; Chambers et al. 2016). Corn Belt cropping systems produce ongoing losses of reactive N to water and the atmosphere which are derived from both fertilizer/manure inputs and extant soil organic matter, prompting a focus on ecosystem-scale N balances (Jaynes and Karlen 2008; David et al. 2010; Martinez-Feria et al. 2018). Nevertheless, ongoing trends in soil C and N under corn-soybean cultivation remain uncertain. Changes in soil C stocks are primarily determined by the balance of crop residue inputs and decomposition of residues and extant C. Greater residue inputs contribute to greater soil C under continuous corn than corn-soybean rotations (Russell et al. 2009; Poffenbarger et al. 2017). However, potential differences in decomposition of litter and soil C between these cropping systems have received less attention and are the focus of our present study.
In several sites throughout the Corn Belt, soil C and N appeared to have stabilized after decades of corn-soybean cultivation, or may be increasing under continuous corn (Russell et al. 2005; David et al. 2009; Poffenbarger et al. 2017). Other studies suggested ongoing declines in soil C under corn-soybean rotations (Liebig et al. 2002; Huggins et al. 2007), as well as a decreased efficiency by which crop residues are retained as soil C in corn-soybean rotations as compared to continuous corn (Poffenbarger et al. 2017). Given that changes over annual to decadal periods are notoriously difficult to detect in large and heterogeneous pools, it is not surprising that the effects of corn-soybean cropping systems on soil C and N stocks remain unclear. However, a long-term synthesis of measurements from across the Corn Belt also demonstrated apparent ongoing losses of soil N (Tomer et al. 2017), along with stable or increasing crop productivity (Yu et al. 2018). Observations of decreasing soil N are also consistent with net negative balances of N inputs vs. outputs measured at the scale of agricultural catchments (Jaynes and Karlen 2008; Gentry et al. 2009; David et al. 2010). In this context, we sought to assess mechanisms that could potentially contribute to ongoing soil C and N losses in these systems. We pose the question: could corn-soybean rotations increase the decomposition of existing soil C and mineralization of associated N due to microbial dynamics associated with fluctuating litter quality?
Corn and soybean rotations involve a temporal fluctuation in the quantity and chemical composition of crop residue production, whereby corn and soybean decompose at different rates and alter the availability of mineral N (Green and Blackmer 1995). Their impacts on decomposition of extant soil C remain unclear, however. Corn typically produces a more than two-fold greater mass of total residue inputs (above- plus belowground) with a higher C:N ratio than soybean (Russell et al. 2009; Jarchow et al. 2015). Decomposition of corn residue in soil thus leads to a significantly longer period of net N immobilization than observed during soybean residue decomposition, which decreases the availability of mineral N for plants (Green and Blackmer 1995). This difference in N mineralization dynamics between corn and soybean litter is a major contributor to the “soybean N credit,” whereby corn grown following soybean typically requires lower inputs of mineral N to achieve optimum yields than corn following corn (Green and Blackmer 1995; Gentry et al. 2001).
Beyond their well-known differences in residue decomposition rates, corn and soybean grown in rotation may also impact the decomposition of extant soil C and subsequent litter inputs. Positive priming of soil C decomposition, defined as an increase in the decomposition of extant soil C in response to new residue inputs, has been demonstrated in many soils and is frequently linked to microbial nutrient demand (Kuzyakov 2010). That is, when a microbial community is confronted with substrates with high C:N relative to their biomass, microbes produce enzymes to degrade soil C to acquire associated N. Conversely, a recent study demonstrated that addition of synthetic N fertilizer to soils under continuous corn cultivation directly suppressed decomposition, presumably by satiating microbial N demand (Mahal et al. 2019). Priming of soil C decomposition induced by corn or soybean root growth has been demonstrated during short-term lab experiments (Helal and Sauerbeck 1984; Fu and Cheng 2002), as well as in field experiments where corn was grown on soils previously planted to pasture or other crops (Mazzilli et al. 2014; Kumar et al. 2016). However, we are unaware of studies that have explicitly examined impacts of priming and its temporal dynamics in the context of corn-soybean rotations. Previous syntheses indicated that crop rotations often increase microbial biomass and decomposition rates relative to monocultures (McDaniel et al. 2014; Tiemann et al. 2015), that microbial growth is a key driver of priming (Kuzyakov 2010), and that microbes grow more efficiently on litter with low C:N ratios (Manzoni et al. 2012). Thus, we propose that corn-soybean rotations could provide ideal conditions for priming: alternating substrates with low and high C:N would fuel microbial growth on the low C:N substrate (soybean) and enhance subsequent decomposition of litter and soil C following addition of the high C:N substrate (corn).
Here, we asked how litter quality (corn vs. soybean) and cropping history (corn after soybean, soybean after corn, and continuous corn with partial residue removal) affected the decomposition of litter and soil C, N mineralization, and microbial biomass production. To isolate these drivers and partition multiple sources of C loss, we used laboratory incubations of soils from a long-term (10 y) no-till field experiment in central Iowa, USA (Jarchow et al. 2015). We used natural-abundance C stable isotope (δ13C) measurements to partition soil respiration and microbial biomass between corn (C4) and soybean (C3) sources. We hypothesized that 1) soybean litter decomposes more rapidly and produces more microbial biomass than does corn litter; 2) priming of soil C decomposition is greater following addition of corn litter than soybean litter; 3) soils planted to corn following soybean have greater rates of litter and soil C decomposition and N mineralization than soils planted to soybean following corn or continuous corn.
Methods
Field experiment
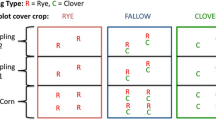
We collected soil samples from the Comparison of Biofuels Systems (COBS) experiment, which is located near Iowa State University in north-central Iowa, USA (41.9212 °N, −93.7499 °W). The predominant soil series at this site are Webster silty clay loam (fine-loamy, mixed, superactive, mesic Typic Endoaquoll) and Nicollet loam (fine-loamy, mixed, superactive, mesic Aquic Hapludoll). For this study, we compared soils that had been managed under continuous corn cultivation or corn-soybean rotations for 10 years (2008–2017). Both phases of the corn-soybean rotation were grown in separate plots during every year, such that three field treatments were examined: continuous corn, corn grown after soybean (corn after soy), and soybean grown after corn (soy after corn). Each treatment had four replicates (0.15 ha plots) arranged in a randomized complete block design. All treatments were managed without tillage and residue was partially removed (52% removal on average; 3.2 Mg ha−1) from the continuous corn treatment. Because residue production from soybean was less than half that of corn in the corn-soybean rotations (Jarchow et al. 2015), partial residue removal from the continuous corn treatment served to decrease differences in above-ground residue inputs between the corn-soybean rotations and the continuous corn treatments. Rates of N fertilization varied based on soil nitrate measurements and averaged 158 kg N ha−1 y−1 for continuous corn and 136 kg N ha−1 y−1 for corn grown in rotation with soybean (no N was applied during the soybean phase). Further management details are provided by Jarchow et al. (2015).
Soil sampling and incubation
Soils were sampled in October 2017 (prior to harvest) from six locations in each field plot from 0 to 15 cm depth using a stainless-steel corer (2.5 cm diameter) and were combined to form a composite sample from each plot (any surface residue was pushed aside prior to sampling the mineral soil). Samples were stored in sealed plastic bags at 4 °C for one week prior to the incubation, and representative subsamples were oven dried to determine moisture content. Soils from each field plot were incubated under each of three litter amendment treatments: added corn litter, added soybean litter, and no added litter. Thus, our experiment had a split-plot design: three cropping system treatments with four field replicates each, and three litter amendment treatments applied to soil from each field plot, for a total of 36 experimental units. Litter (corn and soybean leaf and petiole tissues) was collected from senesced standing biomass at the time of soil sampling, dried at 65 °C, and ground to pass a 1 mm mesh. We used this approach to focus on the most rapidly decomposing components of corn and soybean litter and their contrasting C:N ratios (Russell et al. 2009), given that corn and soybean stalks have greater lignin content and more similar C:N (Córdova et al. 2018). For each experimental unit, we combined 5 g soil (dry mass equivalent) and 0.05 g of litter (where applicable) in an open 50 mL centrifuge tube, mixed the contents by shaking side to side (bulk density was similar to 1 g cm−3), and placed each tube in a glass jar (0.95 L). These ratios of soil:litter are similar in magnitude to what surface soils receive at the field site, given above-ground residue inputs of 2–9 Mg ha−1 y−1 (Jarchow et al. 2015). To maintain microaggregate structure, soils were not sieved, but coarse roots and macrofauna (earthworms) were removed where present. Moisture was adjusted to 65% water-filled pore space by adding deionized water as necessary.
Soil-litter mixtures were then frozen for one week to simulate natural winter conditions at the field site and thawed at 4 °C for 24 h. Jars with samples were fitted with gas-tight lids and two butyl septa for gas sampling and flushing and were purged with humidified CO2-free air to remove ambient CO2 from the jar headspace by venting to the atmosphere. Mole fractions and δ13C values of soil-respired CO2 were measured periodically throughout the 193-day incubation by sampling the headspace of each jar using a syringe with stopcock and injecting a 5 mL gas sample into a tunable diode laser spectrometer (TGA200A, Campbell Scientific, Logan, UT) as described previously (Hall et al. 2017). All δ13C values were calibrated against reference gases traceable to World Meteorological Association primary standards and are reported in ‰ notation relative to Vienna Pee Dee Belemnite. Following each headspace sampling, jars were purged with humidified CO2-free air and remained sealed until the subsequent sampling date, such that we measured the complete cumulative CO2 production over the duration of the incubation. Headspace CO2 was measured at two- to four-day intervals during the first three weeks of the experiment, and progressively at one-, two-, or three-week intervals thereafter as fluxes stabilized (Fig. 1). Deionized water was added following headspace sampling to replenish minor soil moisture losses (due to water vapor loss during flushing) at approximately monthly intervals.

Trends in CO2 production a and δ13C of CO2b over time plotted by cropping and litter addition treatments. Symbols denote cropping treatments (continuous corn, corn after soybean, and soybean after corn, respectively). Yellow, green, and brown colors denote litter addition treatments (corn, soybean, and no litter, respectively)
At the beginning and end of the experiment, soil subsamples (2 g dry mass equivalent) were extracted in a 1:5 mass ratio with 2 M potassium chloride (KCl) solution for colorimetric measurement of ammonium and nitrate + nitrite using phenol-hypochlorite and sulfanilamide-NED colorimetric methods, respectively. Net N mineralization was calculated as the difference between total KCl-extractable mineral N measured at the end and beginning of the incubation. Soil organic C and N concentrations in the pre-incubation soils and litter were measured by combustion on an elemental analyzer coupled to an isotope ratio mass spectrometer (Thermo Finnigan Delta Plus XL, Waltham, MA). These soil subsamples were fumigated with hydrochloric acid vapor prior to analysis to remove any trace carbonates that may have been present.
Microbial biomass C and δ13C values of pre-treatment and post-treatment soil samples were measured using chloroform fumigation-incubation, whereby soil respiration is measured following treatment with chloroform vapor, which lyses cells and increases C availability to re-establishing microbes (Jenkinson and Powlson 1976). Briefly, samples were fumigated with chloroform under vacuum for 24 h, brought to 65% water-filled pore space, and incubated in jars following purging of the headspace with CO2-free air as described above. Accumulated CO2 was measured after 7, 14, and 21 days, by which time CO2 production from fumigated samples was comparable with controls. Microbial biomass C was calculated as the difference in CO2-C production between fumigated and non-fumigated samples over 14 days. The δ13C values of microbial biomass were calculated using the chloroform fumigated and non-fumigated controls as follows, where the subscripts fum, control, and mic denote the fumigated samples, controls, or calculated microbial contributions, respectively:
Data analysis
In the samples that received added corn or soybean litter, we quantified contributions to soil respiration from litter C vs. extant soil C in a given sample using δ13C measurements of respired CO2 and two-end-member mixing models:
Here, flitter_t denotes the calculated fraction of CO2 derived from litter at each measurement period (time t), δ13Cobs denotes the observed δ13C value of respired CO2 at time t, δ13Clitter is the end-member value for litter-derived C (constant over time), and δ13Ccontrol is the observed δ13C value of soil respiration at each time t from control soils that did not receive added litter. Corn and soybean litters had bulk δ13C values of −13.2‰ and−29.1‰, respectively. We assigned δ13Clitter values in the mixing model that were 2‰ greater (−11.2‰ and−27.1‰ for corn and soybean, respectively) than measured bulk litter δ13C (Hall et al. 2017), assuming that respiration of added litter C would be dominantly derived from carbohydrates during the 193-day experiment, and that carbohydrates are 2‰ enriched in δ13C relative to bulk litter C, on average (Bowling et al. 2008). This assumption was necessary to account for high δ13C values observed early in the experiment for samples receiving corn litter (Fig. 1b), which were often greater than corn bulk litter δ13C values. In several limited cases early in the experiment where observed δ13C values were even greater than the defined C4 end-member, these values were corrected by subtracting observed analytical uncertainty of the δ13C measurements (2σ=0.6‰). After calculating flitter_t, the fraction of respiration derived from soil (fsoil_t) was calculated by difference:
From the control soils (no litter added), we further partitioned soil respiration between C4 (dominantly corn) and C3 (dominantly soybean) sources using a two-end-member mixing model:
Here, fC3 is the fraction of soil respiration derived from C3-C, and δ13CC4 and δ13CC3 are the end-member values for corn and soybean described above. The fraction of respiration derived from C4-C was calculated by difference:
We note that legacy C4 and C3 inputs from other plant species growing at the site prior to the present corn and soybean treatments may have also contributed to soil respiration. However, the long-term (decades) management of the study site under corn and soybean cultivation prior to the 10-year experiment implied the dominance of corn- and soybean-derived C as respiration sources. Therefore, we refer to C4- and C3-derived C as corn- and soybean-derived C, respectively. Given that C pools with sub-annual to decadal turnover times dominate soil respiration, impacts of changing atmospheric δ13C values and other long-term fractionating processes in soil, such as microbial carboxylation of soil CO2 (Ehleringer et al. 2000), should have relatively minor impacts on the mixing model results.
Fluxes of CO2 derived from litter and soil C were calculated by multiplying total CO2 fluxes by their fractional contributions. Similarly, C3- and C4-C contributions to respiration were calculated from the control soils. We calculated priming at each time point t as the difference in soil C losses between samples from the same cropping system that received litter and those that did not:
Patterns in C loss from litter and soil over time were quantified by fitting multiple-pool first-order decay models to observed trends in proportion C mass remaining (Fig. 2) for the combined replicates from each treatment using the nls function for non-linear regression in R version 3.5.1. The optimum number of pools for each respiration component (litter and soil) in each cropping system was selected by comparing model AIC values and trends in model residuals among nested models with one, two, or three C pools. The AIC model selection approach does not involve direct comparison of the fitted parameters among models, but rather balances model goodness of fit with parsimony when comparing models of increasing complexity. For example, a three-pool model took the form:

Production of CO2 from added litter C a and soil C b, and corresponding trends in cumulative mass loss of litter c and soil C d over time. Note that panels c and d have different y-axis scales for clarity. Symbols denote cropping treatments (continuous corn, corn after soybean, and soybean after corn, respectively). Yellow, green, and brown colors denote litter addition reatments (corn, soybean, and no litter, respectively)
Where Pr(t) is the proportion of C mass remaining at time t, Pvery_fastis the initial proportion of C in a “very-fast” pool, Pfastis the initial proportion of C in a “fast” pool, (1−Pfast−Pvery_fast) is the initial proportion of C in a “slow” pool, and kfast and kslow are decomposition rate constants (y−1). Mean turnover times were expressed as the inverse of the k values (y). Rate constants and pool sizes calculated from multiple-pool first-order models of CO2 and δ13C time series are widely used to provide inference into decomposition processes among treatments (Collins et al. 2000). Indeed, Reichstein et al. (2000) suggested that this method helps differentiate treatment effects that are consistent over time from transient differences in C mineralization. Differences in a given decomposition model term compared among either the cropping system or litter addition treatments were considered significant with non-overlapping 95% confidence intervals with the Tukey correction for multiple comparisons. Based on the cumulative final C flux data, we assessed effects of cropping treatments, litter addition treatments, and their interactions on decomposition of litter C, soil C, priming, and N mineralization using ANOVA and Tukey post-hoc comparisons.
Results
Soil carbon, respiration, and decomposition modeling
Soil organic C and total N averaged 23±1 mg C g−1 and 1.8±0.1 mg N g−1, respectively, and neither parameter (nor the C:N ratios) differed significantly among cropping treatments (Table 1). Total CO2 production from soils that received added litter was initially 6- to 18-fold greater than unamended soils, declining to a 1.2- to 1.4-fold difference after 190 days (Fig. 1a, Supplementary Table 1). In litter-amended soils, δ13C values of soil-respired CO2 were initially similar to end-member values of corn and soybean litter and gradually converged towards the soils with no added litter, which had intermediate δ13C values (Fig. 1b). In soils without added litter, the continuous corn treatment consistently had the greatest δ13C values, followed by soy after corn and soy after corn. The latter two treatments converged after approximately 96 days (Fig. 1b).
Trends in respired C were best fit by exponential decay functions that included either two or three pools each for litter and soil (Fig. 2, Table 2). The mean R2 values for litter and soil C decomposition models were 0.994 and 0.999, respectively, and all models had R2>0.98. Several decomposition rate constants were estimated as zero, indicating a constant decomposition flux independent of mass remaining. The inverse of the rate constant is the mean turnover time: decomposing litter was best described as the combination of a fast pool with turnover times between 1 and 5 weeks, and a slow pool with turnover times between 11 months and 11 years (varying between litter types and among cropping treatments). The fast pools comprised 45–61% of total litter C among treatments, with the balance allocated to the slow pools (Table 2). Soybean litter consistently decomposed faster than corn litter (significantly greater k-values for the fast pool; Fig. 2, Table 2). However, cropping treatments altered decomposition of corn litter: the proportion of corn litter C in the fast pool tended to be greatest, and the slow pool the was significantly smaller in corn after soy soil than the other cropping treatments.
Decomposition of soil C was best approximated with three-pool models in most treatments, with the exception of continuous corn and soy after corn soils amended with soybean litter, which were best fit with two-pool models. For three-pool models, the very fast pool had turnover times of one to three days and comprised 0.3–1.2% of soil C; the fast pool had turnover times of one to eight months and comprised 2–7% of soil C; and, the slow pool had turnover times of 17–24 years and comprised 93–99% of soil C (Table 2). Addition of corn litter, but not soybean litter, increased the proportion of soil C in the very-fast soil pool relative to the no-litter controls (P<0.05; Table 2). Soils from the corn after soy cropping treatment had a larger proportion of soil C in the fast pool than the others (P<0.05; Table 2).
Cumulative decomposition
Differences in modeled decomposition rates and pool distributions among treatments translated to differences in cumulative decomposition of litter and soil C and net N mineralization. Cumulative decomposition of added litter C (Fig. 3a) differed between corn and soybean litters (P=0.0002) and among cropping treatments (P=0.023), with a marginally significant interaction (P=0.05). Pairwise contrasts revealed that cumulative soybean litter decomposition was significantly greater than corn litter decomposition only in the continuous corn soils. Soybean litter decomposition was highest in continuous corn soil (29% higher than soy after corn; P=0.03) and intermediate in corn after soy soil. In contrast, corn litter decomposition was highest in corn after soy soil, where C loss was 29% and 25% higher than continuous corn and soy after corn, respectively (P<0.05).

Cumulative decomposition of added litter C a and soil C b and net soil N mineralization c plotted by cropping treatments (indicated by labeled rectangles on the upper x axis) and type of added litter (indicated by labels on the lower x axis and colored bars). Error bars indicate standard errors (n=4). Cropping treatments, litter treatments, and their interaction significantly affected litter C decomposition a, and both main treatments (but no interaction) significantly affected soil C decomposition b and net N mineralization c
Cumulative decomposition of soil C (Fig. 3b) also differed among litter treatments (P<0.0001) and cropping treatments (P=0.0006), with no significant interaction (P=0.47). Cumulative soil C decomposition in soils receiving corn litter was 32% greater than soybean litter and 42% greater than the no-litter samples (P<0.0001), averaged across cropping treatments. Cumulative soil C decomposition from corn after soy soil was 20% greater than soy after corn (P=0.008) and 28% greater than continuous corn (P<0.001).
In the control soils (no litter added), the contributions of corn- vs. soybean-derived C to CO2 differed significantly among cropping treatments (Fig. 4; P<0.01). Decomposition of corn-derived C was significantly greater (P=0.005) in corn after soy soils (840±45 μg C g−1) than soy after corn (564±63 μg C g−1), with intermediate values in continuous corn (729±15 μg C g−1). However, decomposition of soybean-derived C was equivalent between corn after soy (413±17 μg C g−1) and soy after corn (368±47 μg C g−1) soils, with significantly lower values (P=0.01) in continuous corn (210±11 μg C g−1; Fig. 4).

Cumulative decomposition of soil C from control soils (no litter added) plotted by cropping treatment; shading in the bars indicate contributions of corn- and soybean-derived C to decomposition as partitioned with δ13C values. Error bars indicate standard errors (n=4). Treatments with different lower-case and upper-case letters differed significantly in corn- and soybean-derived C decomposition, respectively
Prior to the incubation, mineral N was significantly greater (P=0.03) in the soy after corn soils (11±2 μg N g−1) than corn after soy (7±2 μg N g−1) or continuous corn (6±1 μg N g−1). Net N mineralization differed significantly between litter treatments (P<0.0001) and cropping treatments (P=0.0004) with no significant interaction (Fig. 3c, Supplementary Table 2). Net N mineralization was more than two-fold greater in soils amended with soybean litter. However, net N mineralization was 25–28% greater in the corn after soy soils than the other two cropping treatments, irrespective of the litter addition treatment applied.
Microbial biomass
At the beginning of the experiment, microbial biomass C averaged 88 μg C g−1 and tended to be greater in the corn after soy soils (102±15 μg C g−1) than continuous corn (90±9 μg C g−1) and soy after corn (70±28 μg C g−1), although differences were not significant. Corn was the dominant C source for initial microbial biomass, comprising approximately 69%, 65%, and 86% in the corn after soy, soybean-corn, and continuous corn soils. At the end of the experiment, total microbial biomass C significantly differed among litter addition treatments (P=0.0001) and was significantly greater in soils amended with soybean than corn litter or no litter (P<0.05; Fig. 5, Supplementary Table 3). Microbial biomass C in soils amended with soybean litter was approximately equally derived from corn (53±3%) and soybean (47±3%) C, and it was primarily derived from corn in soils amended with corn litter (91±3%) and the control (88±4%).

Microbial biomass C measured at the end of the experiment, plotted by litter addition treatment (cropping treatments did not differ significantly); shading in the bars indicate contributions of corn and soybean C to microbial biomass C partitioned using δ13C values. The dashed line indicates initial microbial biomass C, which did not differ significantly among cropping treatments. Error bars indicate standard errors (n=12). Treatments with different lower-case and upper-case letters differed significantly in corn- and soybean-derived microbial biomass C, respectively
Priming of soil C loss
Priming of soil C loss in the corn and soybean litter amendment treatments peaked on the first sampling date and declined rapidly thereafter, punctuated by a brief period of negative priming for both soybean and corn litters at days 4 and 12, respectively (Fig. 6a). After that, positive priming continued to occur in all soils that received corn litter until at least day 96. However, cropping treatments differed in temporal dynamics of priming, and positive priming tended to be more rapid in corn after soy soils than the other treatments between days 20 and 73 (P<0.10 for pairwise comparisons with the others on all of these days; Fig. 6b). The rapid priming response observed in the corn after soy soils persisted even after accounting for differences in litter mass remaining among cropping treatments (Fig. 6c). However, by the end of the experiment, cumulative priming of soil C loss (Fig. 7) differed between litter treatments (P=0.001) but not among cropping treatments (P=0.44), with no significant interaction (P=0.49). Corn litter addition caused significant soil C loss across all soils (P<0.0001) while priming from soybean litter did not differ from zero (P=0.37). Cumulative priming from corn litter addition measured 434±56 μg C g−1 (Fig. 7). In comparison, decomposition of soil C from samples that received soybean litter or no litter measured 1076±40 μg C g−1 (Fig. 3b). Thus, on average, priming from corn litter addition increased SOC loss to 140% of the soils that received no litter or soybean litter.

Priming of soil C decomposition plotted over time for each measurement date a or as a cumulative flux over time b or vs. litter mass remaining c. Priming is defined as the additional loss of soil C resulting from litter addition relative to an unamended control. Symbols denote cropping treatments (continuous corn, corn after soybean, and soybean after corn, respectively). Yellow, green, and brown colors denote litter addition treatments (corn, soybean, and no litter, respectively)

Cumulative priming of soil C decomposition plotted by the type of added litter, averaged across cropping treatments because of no significant differences in total priming; error bars indicate standard errors (n=12)
Discussion
Our data indicated that the identity of the previous crop and any added litter had key impacts on the decomposition dynamics of soil organic matter in corn-soybean rotations, which differed from continuous corn. Supporting our first hypothesis, soybean litter decomposed faster than corn litter within a given soil (Table 2) and drove a net increase in microbial biomass derived from soybean C (Fig. 4). In contrast, corn litter addition did not significantly increase microbial biomass despite the fact that its cumulative decomposition was statistically equivalent to that of soybean in soils from both phases of the corn-soybean rotation (Fig. 3a, Fig. 4). Supporting our second hypothesis, corn (but not soybean) litter addition consistently primed the decomposition of soil C, with the most rapid priming effects and greatest corn litter decomposition in soils from the corn following soybean treatment (Table 2, Fig. 3a, Fig. 6). In contrast, addition of soybean litter had no significant impact on decomposition of extant C and did not cause net priming of soil C decomposition (Fig. 7). Supporting our third hypothesis, microbial biomass, decomposition of corn litter and soil C, and N mineralization were consistently greatest in soils planted to corn after soybean, even as these soils maintained high decomposition rates of soybean-derived C (equivalent to the soybean after corn treatment). We interpret this finding as a logical consequence of hypotheses 1 and 2: microbial biomass increased during the decomposition of soybean litter, and when confronted with N-poor corn litter, microbes depolymerized organic matter to release N, thus increasing rates of litter and soil C decomposition and N mineralization.
Together, our results suggest a mechanism whereby corn-soybean rotations could enhance mineral N availability by increasing the decomposition rates of both new corn litter inputs and extant soil organic matter. This mechanism may represent an unacknowledged contributor to the well-known phenomenon of decreased N fertilizer requirements for corn grown after soybean known as the soybean N credit (Green and Blackmer 1995; Gentry et al. 2001). It could also potentially contribute to long-term soil C and N losses if the observed increases in decomposition were not offset by other mechanisms. These include increased production of microbial necromass from low C:N litter and its subsequent accrual in soil C (Cotrufo et al. 2013), sorption of soluble litter decomposition products (Córdova et al. 2018) or changes in physico-chemical protection of soil C mediated by increased production of microbial detritus (Tiemann et al. 2015). Although these mechanisms have been demonstrated in longer and more diverse crop rotations, where soil C tends to increase relative to monocultures, corn-soybean rotations remain a notable exception where soil C gains are not typically observed (Russell et al. 2009; McDaniel et al. 2014; Tiemann et al. 2015).
The fact that we did not observe significant differences in soil C concentrations among the field plots sampled in this study (Table 1) is not surprising, given high spatial heterogeneity and large pool sizes that challenge the detection of even large changes in soil C over decadal scales (Goidts et al. 2009). We also note that soil C concentrations in these cultivated soils are much lower than under reference prairie conditions (David et al. 2009; Russell et al. 2005), and thus far from saturation. Finally, we emphasize that our findings from this study are empirical and not normative; there are substantial environmental costs associated with increased N fertilization requirements and potential changes in pest or disease pressure in continuous corn vs. corn-soybean rotations (Gentry et al. 2013), irrespective of any differences in soil C or N concentrations. These collective tradeoffs should also be considered in the context of other potential cropping systems, such as longer and more diverse crop rotations involving small grains and forage legumes, many of which appear effective in maintaining or accruing soil C and N in our region (Russell et al. 2009; Liebman et al. 2013).
Contrasting biogeochemical impacts of corn vs. soybean litters
As expected, added soybean litter decomposed faster than corn litter in accordance with its lower C:N ratio, leading to greater net N mineralization as microbial N demand was satisfied by the litter rather than exogenous sources (Green and Blackmer 1995; Parton et al. 2007). In our study, the absence of priming induced by soybean litter suggests that litter N was largely sufficient to sustain microbial N demand during its decomposition (Kuzyakov 2002). In contrast, the significant priming and lack of net microbial biomass growth observed following corn litter addition was consistent with microbial N limitation (Kuzyakov 2002), a lower microbial C-use efficiency imposed by N limitation (Manzoni et al. 2012), and/or increased turnover of extant microbial biomass driven by priming (Kuzyakov 2010). The burst of soil-derived CO2 observed immediately after corn litter addition (Fig. 6) likely reflected this latter mechanism. Conversely, the greater initial mineral N availability observed in the soybean after corn soils could have explained the slower decomposition of added soybean litter in that treatment, e.g. by downregulating microbial production of non-specific decomposition enzymes as often observed in response to mineral N enrichment (Janssens et al. 2010). We used leaf residue amendments in this study given their importance for N supply in corn and soybean cropping systems (Green and Blackmer 1995) and their high abundance relative to roots (mean root:shoot ratios measured 0.05 at this site; Jarchow et al. 2015) but the potential impacts of adding leaves vs. roots deserve further exploration.
Faster priming and greater litter and SOC decomposition in soils where corn followed soybean
All soils exhibited significant priming when amended with corn litter, but the timescale of the response differed among cropping treatments and was fastest in soils from the corn after soybean treatment (Fig. 6). One plausible explanation for faster priming and the consistent and significant increase in corn litter decomposition, soil C decomposition, and N mineralization in soils from corn after soybean involves the legacy of increased microbial growth on the previous year’s soybean litter. This litter was likely to have decomposed during the most recent growing season (corn phase) just prior to soil sample collection. At the beginning of our laboratory incubation, this soybean-fed microbial community was confronted with detritus from the most recently grown corn roots, which had senesced just before soils were collected. Decomposing these residues likely required additional N, thus stimulating enzymatic attack on soil organic matter, as well as leading to greater decomposition of added corn litter, as we observed (Fig. 3a). This would also explain the fact that soils from this treatment (corn after soy) sustained high rates of decomposition of older soybean-derived C (Fig. 4) despite being most recently planted to corn.
Other explanations for these results are possible but less likely. Differences in residue inputs, especially roots, are key drivers of soil C dynamics in continuous corn and corn-soybean rotations under conventional management (Russell et al. 2009; Brown et al. 2014). However, in our study, these differences cannot explain faster decomposition of added corn litter (Fig. 3a) nor the sustained decomposition of soybean-derived C in the corn after soy treatment (Fig. 4). Furthermore, in the control soils (no litter added), the mean mass ratio of C decomposed to CO2 relative to net N mineralization was similar among all three cropping treatments (32–33) despite their differences in total soil C decomposition (Fig. 3b). This consistent coupling between mineralization of C and N would not be expected if differences in recent residue inputs were responsible for differences in soil C decomposition among cropping treatments, as corn and soybean vary substantially in their C:N ratios (Green and Blackmer 1995). In addition, the partial (~52%) above-ground residue removal from the continuous corn treatment in this 10-year experiment served to minimize long-term differences in above-ground residue inputs between continuous corn and the corn-soybean rotation soils by compensating for decreased residue inputs during the soybean phase (Jarchow et al. 2015). To minimize differences in the most recent above-ground corn residue inputs between the continuous corn and corn after soy treatments, soils were sampled prior to harvest, when partial residue removal occurred. Finally, shallow fine roots are arguably the most important sources of C inputs to these systems (Russell et al. 2009), and their biomass (0–30 cm) was statistically similar among soy after corn, soy after corn, and continuous corn treatments for most years of this study (Jarchow et al. 2015).
Implications for the fertility of corn-soybean rotations
Our findings have intriguing implications for soil C and N cycling in corn-soybean cropping systems. Previous work suggested that the soybean N credit results from a combination of increased net N mineralization relative to that in continuous corn production and an increase in residual soil N due to symbiotic N fixation in the soybean plant (Green and Blackmer 1995; Gentry et al. 2001). Conversely, decreased net N mineralization explains the “yield penalty” often observed under continuous corn (Gentry et al. 2013). Expanding on this work, our data suggest that increased N mineralization linked to accelerated decomposition of extant soil organic matter and recent corn litter inputs—not simply lower soil N immobilization and faster N release from decaying soybean residue—may also contribute to the soybean N credit. Empirical estimates of the soybean N credit vary tremendously, e.g. from −22 to 210 kg N ha y−1 among Wisconsin cropping systems (Bundy et al. 1993), with enormous implications for N fertilizer use in the Corn Belt. Some long-term corn-soybean cropping systems appear to have net negative N balances (Jaynes and Karlen 2008; Gentry et al. 2009; David et al. 2010), which may contribute to a regional trend of lower soil N stocks (Tomer et al. 2017). Our data suggest that better prediction of the magnitude of the soybean N credit, and its long-term sustainability, may require an explicit consideration of the dynamics of decomposition priming under field conditions.
Our findings are also consistent with observations of decreased soil C retention in corn-soybean rotations even after accounting for lower C inputs from soybean residues. Poffenbarger et al. (2017) observed that corn-soybean rotations required more than 30% greater C input to maintain equivalent soil C stocks as compared with a continuous corn system (4.2 vs 3.2 Mg residue C ha−1 y−1) over a 14–16 y field trial. Similarly, Russell et al. (2009) observed that soil C at one long-term experimental site in Iowa (Nashua) was 12% lower under long-term corn-soybean rotations than continuous corn, despite the fact that mean residue C inputs were only 3% lower. Conversely, a continuous corn-oats-alfalfa rotation had slightly greater soil C than continuous corn despite lower residue C inputs in that study. Huggins et al. (2007) found similar stocks of C3-derived C in surface soils under continuous corn and corn-soybean rotations, despite the absence of contemporary inputs of C3-C in the former treatment—implying increased decomposition of legacy soybean C inputs under corn-soybean rotations than continuous corn.
Our laboratory study demonstrates the plausibility of enhanced litter and SOC decomposition under soil from corn-soybean rotations vs. continuous corn, controlling for climate—a hypothesis that bears further testing under field conditions. This is an important finding in that previous work has often focused on decreased net primary productivity of soybean as a primary driver of observed differences in SOC stocks between these systems (e.g. Russell et al. 2009). Other factors may also contribute to shifts in SOC dynamics under corn-soybean rotations as compared with continuous corn, such as greater early-season soil temperature following soybean due to lower residue cover (Huggins et al. 2007), or differences in the physical and chemical partitioning of residues during decomposition (Tiemann et al. 2015). Mahal et al. (2019) recently demonstrated that N fertilizer addition could directly suppress SOC decomposition in continuous corn, presumably by inhibiting microbial decomposition activity related to N acquisition. Further evaluation of these mechanisms is critical, as even small changes in decomposition rates within and among cropping systems have key implications for soil C and N cycling and related environmental impacts in one of the dominant agroecosystems of North America.
References
Bowling DR, Pataki DE, Randerson JT (2008) Carbon isotopes in terrestrial ecosystem pools and CO2 fluxes. New Phytol 178:24–40. https://doi.org/10.1111/j.1469-8137.2007.02342.x
Brown KH, Bach EM, Drijber RA, et al (2014) A long-term nitrogen fertilizer gradient has little effect on soil organic matter in a high-intensity maize production system. Glob Change Biol 20:1339–1350. https://doi.org/10.1111/gcb.12519
Bundy LG, Andraski TW, Wolkowski RP (1993) Nitrogen credits in soybean-corn crop sequences on three soils. Agron J 85:1061. https://doi.org/10.2134/agronj1993.00021962008500050020x
Chambers A, Lal R, Paustian K (2016) Soil carbon sequestration potential of US croplands and grasslands: implementing the 4 per thousand initiative. J Soil Water Conserv 71:68A–74A. https://doi.org/10.2489/jswc.71.3.68A
Collins HP, Elliott ET, Paustian K et al (2000) Soil carbon pools and fluxes in long-term corn belt agroecosystems. Soil Biol Biochem 32:157–168. https://doi.org/10.1016/S0038-0717(99)00136-4
Córdova SC, Olk DC, Dietzel RN, Mueller KE, Archontouilis SV, Castellano MJ (2018) Plant litter quality affects the accumulation rate, composition, and stability of mineral-associated soil organic matter. Soil Biol Biochem 125:115–124. https://doi.org/10.1016/j.soilbio.2018.07.010
Cotrufo MF, Wallenstein MD, Boot CM, et al (2013) The Microbial Efficiency-Matrix Stabilization (MEMS) framework integrates plant litter decomposition with soil organic matter stabilization: do labile plant inputs form stable soil organic matter? Glob Change Biol 19:988–995. https://doi.org/10.1111/gcb.12113
David MB, McIsaac GF, Darmody RG, Omonode RA (2009) Long-term changes in Mollisol organic carbon and nitrogen. J Environ Qual 38:200. https://doi.org/10.2134/jeq2008.0132
David MB, Drinkwater LE, McIsaac GF (2010) Sources of nitrate yields in the Mississippi River basin. J Environ Qual 39:1657. https://doi.org/10.2134/jeq2010.0115
Ehleringer JR, Buchmann N, Flanagan LB (2000) Carbon isotope ratios in belowground carbon cycle processes. Ecol Appl 10:412–422. https://doi.org/10.1890/1051-0761(2000)010[0412:CIRIBC]2.0.CO;2
Fu SL, Cheng WX (2002) Rhizosphere priming effects on the decomposition of soil organic matter in C-4 and C-3 grassland soils. Plant Soil 238:289–294. https://doi.org/10.1023/A:1014488128054
Gentry LE, Below FE, David MB, Bergerou JA (2001) Source of the soybean N credit in maize production. Plant Soil 236:175–184. https://doi.org/10.1023/A:1012707617126
Gentry LE, David MB, Below FE, Royer TV, McIsaac GF (2009) Nitrogen mass balance of a tile-drained agricultural watershed in east-Central Illinois. J Environ Qual 38:1841. https://doi.org/10.2134/jeq2008.0406
Gentry LF, Ruffo ML, Below FE (2013) Identifying factors controlling the continuous corn yield penalty. Agron J 105:295–303. https://doi.org/10.2134/agronj2012.0246
Goidts E, van Wesemael B, Crucifix M (2009) Magnitude and sources of uncertainties in soil organic carbon (SOC) stock assessments at various scales. Eur J Soil Sci 60:723–739. https://doi.org/10.1111/j.1365-2389.2009.01157.x
Green CJ, Blackmer AM (1995) Residue decomposition effects on nitrogen availability to corn following corn or soybean. Soil Sci Soc Am J 59:1065. https://doi.org/10.2136/sssaj1995.03615995005900040016x
Hall SJ, Huang W, Hammel KE (2017) An optical method for carbon dioxide isotopes and mole fractions in small gas samples: tracing microbial respiration from soil, litter, and lignin. Rapid Commun Mass Spectrom 31:1938–1946. https://doi.org/10.1002/rcm.7973
Helal HM, Sauerbeck DR (1984) Influence of plant roots on C and P metabolism in soil. Plant Soil 76:175–182. https://doi.org/10.1007/BF02205578
Huggins DR, Buyanovsky GA, Wagner GH, Brown JR, Darmody RG, Peck TR, Lesoing GW, Vanotti MB, Bundy LG (1998) Soil organic C in the tallgrass prairie-derived region of the Corn Belt: effects of long-term crop management. Soil Till Res 47:219–234. https://doi.org/10.1016/S0167-1987(98)00108-1
Huggins DR, Allmaras RR, Clapp CE et al (2007) Corn-soybean sequence and tillage effects on soil carbon dynamics and storage. Soil Sci Soc Am J 71:145–154. https://doi.org/10.2136/sssaj2005.0231
Janssens IA, Dieleman W, Luyssaert S, et al (2010) Reduction of forest soil respiration in response to nitrogen deposition. Nat Geosci 3:315–322. https://doi.org/10.1038/ngeo844
Jarchow ME, Liebman M, Dhungel S, Dietzel R, Sundberg D, Anex RP, Thompson ML, Chua T (2015) Trade-offs among agronomic, energetic, and environmental performance characteristics of corn and prairie bioenergy cropping systems. GCB Bioenergy 7:57–71. https://doi.org/10.1111/gcbb.12096
Jaynes DB, Karlen DL (2008) Sustaining soil resources while managing nutrients. In: Final report: gulf hypoxia and local water quality concerns workshop. American Society of Agricultural and Biological Engineers, St. Joseph, pp 149–158
Jenkinson DS, Powlson DS (1976) The effects of biocidal treatments on metabolism in soil—I. Fumigation with chloroform. Soil Biol Biochem 8:167–177. https://doi.org/10.1016/0038-0717(76)90001-8
Kumar A, Kuzyakov Y, Pausch J (2016) Maize rhizosphere priming: field estimates using 13C natural abundance. Plant Soil 409:87–97. https://doi.org/10.1007/s11104-016-2958-2
Kuzyakov Y (2002) Review: Factors affecting rhizosphere priming effects. J Plant Nutr Soil Sci 165:382–396. https://doi.org/10.1002/1522-2624(200208)165:4<382::AID-JPLN382>3.0.CO;2-#
Kuzyakov Y (2010) Priming effects: interactions between living and dead organic matter. Soil Biol Biochem 42:1363–1371. https://doi.org/10.1016/j.soilbio.2010.04.003
Liebig MA, Varvel GE, Doran JW, Wienhold BJ (2002) Crop sequence and nitrogen fertilization effects on soil properties in the western corn belt. Soil Sci Soc Am J 66:596–601. https://doi.org/10.2136/sssaj2002.5960
Liebman M, Helmers MJ, Schulte LA, Chase CA (2013) Using biodiversity to link agricultural productivity with environmental quality: results from three field experiments in Iowa. Renewable Agric Food Syst 28:115–128. https://doi.org/10.1017/S1742170512000300
Mahal NK, Osterholz WR, Miguez FE, Poffenbarger HJ, Sawyer JE, Olk DC, Archontoulis SV, Castellano MJ (2019) Nitrogen fertilizer suppresses mineralization of soil organic matter in maize agroecosystems. Front Ecol Evol 7. https://doi.org/10.3389/fevo.2019.00059
Manzoni S, Taylor P, Richter A, et al (2012) Environmental and stoichiometric controls on microbial carbon-use efficiency in soils. New Phytol 79–91. https://doi.org/10.1111/j.1469-8137.2012.04225.x
Martinez-Feria RA, Castellano MJ, Dietzel RN, Helmers MJ, Liebman M, Huber I, Archontoulis SV (2018) Linking crop- and soil-based approaches to evaluate system nitrogen-use efficiency and tradeoffs. Agric Ecosyst Environ 256:131–143. https://doi.org/10.1016/j.agee.2018.01.002
Mazzilli SR, Kemanian AR, Ernst OR, Jackson RB, Piñeiro G (2014) Priming of soil organic carbon decomposition induced by corn compared to soybean crops. Soil Biol Biochem 75:273–281. https://doi.org/10.1016/j.soilbio.2014.04.005
McDaniel MD, Tiemann LK, Grandy AS (2014) Does agricultural crop diversity enhance soil microbial biomass and organic matter dynamics? A meta-analysis. Ecol Appl 24:560–570. https://doi.org/10.1890/13-0616.1
Parton W, Silver WL, Burke IC, et al (2007) Global-scale similarities in nitrogen release patterns during long-term decomposition. Science 315:361–364. https://doi.org/10.1126/science.1134853
Poffenbarger HJ, Barker DW, Helmers MJ, Miguez FE, Olk DC, Sawyer JE, Six J, Castellano MJ (2017) Maximum soil organic carbon storage in Midwest U.S. cropping systems when crops are optimally nitrogen-fertilized. PLoS One 12:e0172293. https://doi.org/10.1371/journal.pone.0172293
Prince SD, Haskett J, Steininger M, Strand H, Wright R (2001) Net primary production of U.S. Midwest croplands from agricultural harvest yield data. Ecol Appl 11:1194–1205. https://doi.org/10.2307/3061021
Reichstein M, Bednorz F, Broll G, Kätterer T (2000) Temperature dependence of carbon mineralisation: conclusions from a long-term incubation of subalpine soil samples. Soil Biol Biochem 32:947–958
Russell AE, Laird DA, Parkin TB, Mallarino AP (2005) Impact of nitrogen fertilization and cropping system on carbon sequestration in Midwestern Mollisols. Soil Sci Soc Am J 69:413–422
Russell AE, Cambardella CA, Laird DA, Jaynes DB, Meek DW (2009) Nitrogen fertilizer effects on soil carbon balances in Midwestern US agricultural systems. Ecol Appl 19:1102–1113
Tiemann LK, Grandy AS, Atkinson EE, Marin-Spiotta E, McDaniel MD (2015) Crop rotational diversity enhances belowground communities and functions in an agroecosystem. Ecol Lett 18:761–771. https://doi.org/10.1111/ele.12453
Tomer MD, James DE, Schipper LA, Wills SA (2017) Use of the USDA National Cooperative Soil Survey soil characterization data to detect soil change: a cautionary tale. Soil Sci Soc Am J 81:1463–1474. https://doi.org/10.2136/sssaj2017.06.0198
Yu Z, Lu C, Cao P, Tian H (2018) Long-term terrestrial carbon dynamics in the Midwestern United States during 1850–2015: roles of land use and cover change and agricultural management. Glob Chang Biol 24:2673–2690. https://doi.org/10.1111/gcb.14074
Acknowledgements
Funding was provided in part by the Center for Global and Regional Environmental Research at the University of Iowa. We thank Matt Liebman for thoughtful comments and for facilitating access to the field site, Carlos Tenesaca for assistance in the lab, the McNair Scholars program for providing research support for ARM, and two anonymous reviewers who provided thoughtful and useful comments.
Author information
Authors and Affiliations
Contributions
AER, ARM, and SJH conceived the research and designed and performed the experiment. SJH analyzed data and wrote the paper, with contributions from AER.
Corresponding author
Additional information
Responsible Editor: Elizabeth M Baggs.
Publisher’s note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
About this article

Cite this article
Hall, S.J., Russell, A.E. & Moore, A.R. Do corn-soybean rotations enhance decomposition of soil organic matter?. Plant Soil 444, 427–442 (2019). https://doi.org/10.1007/s11104-019-04292-7
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11104-019-04292-7