
Abstract
Aims
How landscape configuration, dispersal agents and diaspore features interact to determine the input of seeds into the soil, which is the first part of the formation of a soil seed bank, is a major challenge. This study explored the role of diaspore traits and sand surface configuration in determining the number of seeds that become buried during wind dispersal.
Methods
We investigated seed burial probability of 36 species with different diaspore traits under eight wind speeds and three sand surface configurations during wind dispersal by using a wind tunnel.
Results
Wind speed rather than diaspore traits and sand surface configuration was the most important factor affecting the burial of seeds. Effects of slope direction and surface barrier on seed burial were regulated by wind speed. Effects of diaspore traits on seed burial were only significant at high wind speeds, and seed burial was more likely to be formed for small or flat elongated diaspores than for large or spherical ones.
Conclusions
The effect of diaspore traits on seed burial is modified by the shifts in wind speed and sand surface configuration.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Soil seed bank is generally defined as the living seeds present in the soil and the litter layer on the soil surface (Simpson et al. 1989), which provides diaspores for population maintenance and community assembly. It is relevant for all vegetation processes and of practical significance in ecological restoration of degraded ecosystems (Harper 1977; Thompson and Grime 1979). The formation of soil seed bank consists of seed dispersal, settlement and burial (Chang et al. 2008; Rotundo et al. 2015). Regardless of seed bank types, i.e. transient and persistent seed bank (Thompson and Grime 1979), the first step in the formation of a soil seed bank is that the seeds need to be buried (Baskin and Baskin 2014). In addition, burial is believed to be a way to protect seeds from germination and predation (Fenner and Thompson 2005).
A lot of species are maintained by possessing a soil seed bank, and population recruitment and community assembly are dependent on the formation of a soil seed bank in many cases (Sletvold and Rydgren 2007; Helsen et al. 2015). To understand the characteristics and functions of the soil seed bank, we need to study the size, type, composition, and spatiotemporal pattern of soil seed banks. On the other hand, we need to study soil seed bank formation and its relevance to various biotic and abiotic factors. In particular, we need to know how seeds are buried. For restoration and biodiversity conservation, understanding how seeds are buried is more important than knowing seed bank size and pattern, because the former can help us understand why seeds of the target species are not available and how we can increase seed availability.
In general, landscape configuration, dispersal agents and diaspore features are associated with seed bank formation (López 2003; Wang et al. 2005; Yan et al. 2005). The factors of landscape configuration affecting seed bank formation can include matrix type, slope direction, slope angle, vegetation cover, and surface barriers (Redbo-Torstensson and Telenius 1995; Li 2008). They may affect seed deposition pattern by reducing wind speed, changing wind turbulence and intercepting seeds (Chambers 2000; Nathan and Muller-Landau 2000). The dispersal agents like animals, water and wind play their roles in the formation of seed bank. The predation preference of animals, water flow direction and wind condition can affect the dispersal process (Bakker et al. 1996; Navie et al. 1996; Obeso et al. 2011), leading to variation in seed bank formation. Diaspore features like morphological traits, appendage types and aerodynamic attributes can separately or together control the burial of seeds (Du et al. 2007; Symes 2012). Diaspore morphological traits may have dual conflicting roles in seed burial: promoting or impeding (Maddox and Carlquist 1985).
How landscape configuration, dispersal agents and species features interact to determine seed burial has been a research focus for plant ecologists, conservationists and ecological engineers (Baskin and Baskin 2014). Seed burial is a complex process, affected by factors such as soil features, seed characters, natural dispersal forces and tillage practices (Chambers 1991; Benvenuti 2007; Egawa et al. 2009). However, the process and mechanism of seed burial are not been well known. Here, we concentrate on how diaspore traits, wind speed and sand surface configuration interact to determine how many seeds are buried by wind during dispersal.
Anemochores, accounting for an average of 10% to 30%, and up to 70% of the species in the temperate vegetation (Willson et al. 1990; Oudtshoorn and Rooyen 1999), appear in all vegetation types in the world, and they are especially common in open habitats such as steppe, wasteland, desert, sand dune and road edge (Collins and Uno 1985). The dispersal of diaspores by wind includes primary and secondary dispersal (Chambers and Macmahon 1994; Oudtshoorn and Rooyen 1999). At present, systematic and empirical studies on how soil seed banks are formed, especially on how diaspore traits, sand surface configuration and wind speed interact to determine seed burial at a site have rarely been conducted. Lack of the information on seed burial impedes judging seed fate and determining plant adaptive strategy in windy environments and screening vegetation restoration measures for degraded open habitats.
Proper experimental methods are essential for quantitatively describing seed dispersal processes. Due to the high variability of wind speed and direction, field observations make it difficult to meet the demands for a quantitative description of seed dispersal and seed bank formation (Maler et al. 1999). Similarly, long-term monitoring is of little help in establishing the quantitative relations between diaspore traits, sand surface configuration and wind speeds due to unpredictable features of the field condition (Driscoll et al. 2014). Controlled experiments, taking into account the combination of various elements and their gradients to determine and compare processes and interactive relations of different experimental designs, are necessary for the research on seed dispersal and burial (Nathan et al. 2008). Thus, wind tunnels are being used to conduct controlled experiments on seed dispersal by wind and some interesting results have been obtained (Maler et al. 1999; Baker and Beck 2008; Pinceel et al. 2016).
Trichomes, pappus, plumes, and other appendages on diaspores provide them with wind dispersal capacity (Andersen 1993; Maler et al. 1999; Vittoz and Engler 2007). Many studies indicate that the capacity for wind dispersal is affected not only by type of appendage but also by other attributes such as diaspore mass, terminal velocity, wing loading, projected area and shape index (Nathan and Muller-Landau 2000; Aavik and Helm 2017). Chambers (1991) demonstrated that small seeds were more likely to be trapped by small soil particles than large ones. Peart (1981) found that awns serve as a mechanism for orientation and interposition into soil after diaspore dispersal. However, few reports have been found on the functions of seed attributes in seed anchoring and burial.
In our study, we aimed to determine the role of diaspore traits, sand surface configuration and wind speed in the number of seeds that become buried during the dispersal by wind. Thus, a field wind tunnel experiment was conducted, and various diaspore traits, different sand surface configuration and wind speeds were considered.
Materials and methods
Definition of seed distribution pattern
The seed distribution pattern after secondary dispersal by wind consists of two parts: horizontal distribution and vertical distribution. Horizontal distribution has three cases: unshifted (diaspores stayed in situ), distantly-dispersed (diaspores dispersed a certain distance) and infinitely-dispersed (diaspores dispersed to infinity). Vertical distribution includes three situations: aboveground distribution (diaspores caught on barriers, and not contacting the ground), on the ground distribution (diaspores left on the ground), and underground distribution (diaspores buried). In this study, we considered diaspores burial by sand to be soil seed bank formation.
Diaspore selection and traits measurement
Ignoring family characteristics of the species, each diaspore was characterized by its morphological traits. Different gradients were set according to the diaspore size and shape. Air dried diaspores of 36 species, with (wing, thorn or hair) or without appendages, a mass range of 1.12–231.46 mg and a shape index range of 0.001–0.194 (Table 1), were selected as the actual species models to conduct the wind tunnel experiment. Nine of the species did not have an appendage (mass ranged from 1.115 mg to 66.6 mg, shape index ranged from 0.01 to 0.101), seven species had hairs (mass ranged from 1.12 mg to 11.625 mg, shape index ranged from 0.001 to 0.194), six species had thorns (mass ranged from 1.065 mg to 74.66 mg, shape index ranged from 0.004 to 0.067), and fourteen species had wings (mass ranged from 4.77 mg to 231.46 mg, shape index ranged from 0.002 to 0.173).
To facilitate soil seed bank investigation, diaspores with thorns, wings and without appendage were lightly sprayed with red aerosol paint, while diaspores with hairs were colored using red water-based markers. The dyed diaspores were naturally air-dried and placed in plastic boxes to ensure the integrity of morphological structure.
Diaspore traits like mass, shape index, wing loading, projected area and terminal velocity are important indicators for measuring the process of seed wind dispersal (Casper and Grant 1988; Matlack 1992; Casseau et al. 2015) and seed bank formation (Peart 1984; Thompson et al. 1993; Funes et al. 1999). In our study, traits of dyed-diaspores were measured to avoid the influence of dyes on the attribute value. Twenty intact diaspores of each species were selected for the measurements. Length, width and thickness of diaspore were measured with Vernier caliper (0.01 mm accuracy). The diaspore shape index was calculated as: \( \mathrm{Shape}\ \mathrm{index}=\mathrm{variance}\left(\frac{\mathrm{Length}}{\mathrm{Length}},\frac{\mathrm{Width}}{\mathrm{Length}},\frac{\mathrm{Height}}{\mathrm{Length}}\right) \) (Thompson et al. 1993).
Diaspore mass was measured by an electronic balance (0.1 mg accuracy). After scanning with a digital scanner, the projected area of each diaspore was determined with the image analysis software Motic Image Plus 2.0 (Motic China Group Co., Ltd., USA). The wing loading was calculated as the ratio of seed mass to projected area (Matlack 1987; Howlett 1995; Greene and Johnson 1997). Terminal velocity was defined as a steady and maximum speed when the air resistance is equal to the pull of gravity during a free fall of diaspore in the motionless air (Green 1980). The terminal velocity was measured with an apparatus described by Zotz et al. (2016).
The wind tunnel used
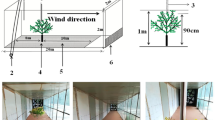
Controlled experiments with wind tunnels have become a new trend in the study of seed wind dispersal processes (Baker and Beck 2008; Lemke et al. 2009; Johnson and Fryer 1992; Pinceel et al. 2016). The use of a large-scaled portable field wind tunnel conforms to the physical similarity of horizontal wind and can accurately simulate the wind dispersal of diaspores. The wind tunnel used was 2 m high, 2 m wide, with a 20 m-long test section, and the sections could be adjusted so that the wind tunnel could be installed on an undulating terrain. Wind speed was monitored with a Magnesense II Differential Pressure Transmitter (MS2-W102-LCD, Dwyer Instruments Inc., Indiana, USA) out of the wind tunnel and a Pitot tube (160–96, Dwyer Instruments Inc., Indiana, USA) within the wind tunnel. A Pitot tube (a tubular device that measures the total pressure and the static pressure of the airflow to determine the velocity of the wind), located 10 m away from the power section, inserted through the roof, with the pitot hole 1 m from the ground, was used to measure the wind speed at 1 m above the sand surface (Fig. 1).

The sketch graph of seed burial experiment by wind tunnel. 1-Starting point section, 2-Power section, 3 and 4-Diversion section, 5-Rectifying section, 6-Transitional section, 7-Draft gear, 8-Road wheel, 9-Electric landing gear, 10-Experimental section, 11-Schematic plan of barriers pavement, 12-Starting point of the experiment, 13-Straw check boards, 14-Diagram of the straw check board dimensions, 15-The experimental section of wind tunnel, 16-Pitot tube, 17-Diaspore, 18-Pullable iron cover, 19-Differential pressure transmitter, 20-wire net. The wind tunnel used is 2 m in height, 2 m in width, and 20 m in length. Three sand surface configurations: upwind slope (6–8°), with the power section of wind tunnel at the lower part of the slope (a), downwind slope (6–8°), with the power section at the upper part of the slope (b), upwind slope with barriers, using straw check boards, 1 m × 1 m in size and 0.1 m in height, as barriers (c). The straw check board is 2 m away from the starting point of the experiment and the barrier section is 8 m in length, 2 m in width. One and two halves straw check boards transversely is designed to ensure one entire straw check board at the central of transverse section. Ten diaspores of each species are landed on the surface 10 m away from the power section, and covered with an iron cover. The iron cover is pulled up to release diaspores when the experiment starts. A wire net is set up at the outlet of wind tunnel to recollect the diaspores (d)
Preparation of sand surface configuration
We eliminated the effects of substratum attributes like roughness, moisture content and non-substrate impurities on the final deposition of seeds (Johnson and Fryer 1992; Schurr et al. 2005; Sun et al. 2012), and only considered the influence of the slope direction and obstacles on seed burial. Thus, the relatively ideal eolian sand from moving sand dune was chosen as the sand surface matrix. A moving sand dune (106°35′E, 40°17’N) in the Ulanbuh sand dune field in northwestern China, with the gradient about 6 degrees and length about 50 m, was selected to conduct the wind tunnel experiment.
To remove the impact of non-substrate impurities (such as stones, irregular plants, solid refuses, etc.) on experiment results, vegetation on the sand surface was manually removed, and the matrix to the depth of 30 cm was sieved with a mesh size of 5 mm. The sand used in this experiment was naturally air-dried to avoid influence of matrix moisture on the results.
Before the experiment started, boards (1.5 m in length, 2 m in width) with their surfaces covered with sand grains were placed over the transitional section of the wind tunnel to prevent the sand from moving and burying the experimental section and to ensure that the land surface of the transitional section was the same as the experimental section.
To avoid impacts of footprints on underlying surface, boards (1.5 m in length, 0.3 m in width, and 0.01 m in thickness) were used to cover the sand surfaces near the walls of the wind tunnel when investigating the number of seeds buried in the wind tunnel. The boards were put upright against the walls on both sides of the wind tunnel after finishing each investigation.
Controlled experiment on seed burial
Thirty-six species with different diaspore traits (appendage type, diaspore mass, projected area, shape index, wing loading and terminal velocity) were used to conduct the experiments under three sand surface configurations: upwind slope, downwind slope, and upwind slope with barriers. Eight wind speeds were applied. Ten diaspores of each species were employed each time and five replicas were adopted for each treatment. In total, 43,200 diaspores (times) (36 species×3 sand surface configurations×8 wind speeds×10 diaspores×5 replicas = 43,200) were involved.
When the experiment was conducted on the upwind slope, the power section of the wind tunnel was at the lower part of the slope, i.e. the wind blew towards the slope (Fig. 1a). While on the downwind slope, the power section was at the upper part of the slope, i.e. the wind blew downward the slope (Fig. 1b).
Barriers can affect seed dispersal by regulating the wind speed and surface roughness (Nathan and Muller-Landau 2000). To explore the presence or absence of obstacles on seed burial by sand, a standardized, controllable, and commonly-used sand-fixing straw check boards were selected as the experimental barrier. The straw barriers were located only on the upwind slope, and they consisted of straw placed upright in the sand around the edges of a 1 m × 1 m square; height above the surface was 0.1 m. Straw check boards were set at 2 m away from the starting point of the experiment to ensure that the diaspores were dispersing before they encountered the barriers. The barrier section was 8 m long (parallel to the wind tunnel), 2 m wide (perpendicular to the wind tunnel), i.e. there were eight straw check boards (8 barriers) lengthwise, one and two halves straw check boards (2 barriers) breadthwise. The reason for designing one and two halves straw check boards transversely was to ensure an entire straw check board at the central part of transverse section of the wind tunnel (Fig. 1c).
The wind speeds (measuring 1 m height from the ground) were set at 3, 4.3, 5.6, 6.9, 8.2, 9.5, 10.8 and 12.1 m·s−1, increased by 1.3 m·s−1 between two adjacent wind speeds, and at a height of 10 m from the ground they were 4.1, 6.0, 7.7, 9.5, 11.3, 13.1, 14.9 and 16.7 m·s−1, respectively (Touma 1977), corresponding to Beaufort wind scale from level 4 to 7 (Mather 1987). Ten diaspores of each species were released from 10 cm above the sand surface near the starting point of the experiment section to make diaspores randomly lay on the surface. After landing, diaspores were covered with a specially designed iron cover. The iron cover was pulled up to release the diaspores when the wind reached a constant speed. Two minutes later, the wind tunnel was turned off. A wire net was set up at the outlet of wind tunnel to recollect the diaspores (Fig. 1d). Each wind speed was tested following the same procedures until all wind speeds were completed. Five replicas were applied.
Seed burial investigation
Seed burial was determined after the wind had been stopped. Since the diaspores involved might be on the ground, dispersed to infinity or buried underground, we first found the diaspores on the ground and captured by wire net and straw check boards; thus, we could calculate how many diaspores were buried belowground. Then, we used the shovels and brushes to retrieve the diaspores buried within the entire experimental section. The search was not stopped until all 10 diaspores were found. Since 10 diaspores were released each time and 5 replicas were used, we calculated the probability of seed burial, as the proportion of 50 diaspores that were buried in sand.
Data analysis
Ordination analysis was conducted to assess variation of seed burial probability corresponding to shape index, diaspore mass, projected area, wing loading, terminal velocity, appendage type, sand surface configuration and wind speed. Canonical correspondence analyses (CCAs) based on correlation matrixes of seed burial probability and explanatory factors were conducted using Canoco 5.0 (version 5.0, Microcomputer Power, Ithaca, NY, USA) (Tackenberg 2003). The contribution of each explanatory variable to the variation in seed burial probabilities was tested following standardized interactive-forward-selection procedures. The variation of seed burial under different wind speeds was analyzed, the polynomial regression on wind speed and average seed burial probability was performed, and the Pearson correlation analysis on diaspore attributes and seed burial probability was conducted. The stepwise regression analysis comparing the explanations of diaspore traits (mass, projected area, shape index, wing loading and terminal velocity) on the variation of seed burial probability under different wind speeds was applied using IBM SPSS Statistics 22.0 (IBM Corporation 1989–2013, USA). Plots were drawn using AutoCAD 2007 (Autodesk Incorporation 1982–2006, USA) and Origin Pro 8.5 (OriginLab Corporation 1991–2010, USA).
Results
Effect of wind speed, diaspore traits and sand surface configuration on seed burial
All factors combined explained 39.1% of the total variation in probability of seed burial. Wind speed, positively related to seed burial probability, was the most important factor affecting the burial number of seeds, and explained 30.4% of the variation. Diaspore traits, explained 8.6% of variation in seed burial probability. However, sand surface configuration explained only 0.1% of the variation (p > 0.01), and had no significant effect on seed burial. The contribution of wind speed, diaspore traits and sand surface configuration to the variation in seed burial probability were 77.8%, 21.9% and 0.3%, respectively (Table 2).
Seed burial probability in relation to sand surface configuration
The total probabilities of seed burial under the three sand surface configurations were ordered as downwind slope > upwind slope > upwind slope with barriers (Fig. 2a). The wind speed threshold for seed burial on both the upwind and downwind slopes was 6.9 m·s−1, while that on the upwind slope with barriers was 8.2 m·s−1 (Fig. 2a). The straw check boards increased the wind speed threshold for seed burial. The differences in seed burial probability between upwind slope and downwind slope became obvious when the wind speed was higher than 4.9 m·s−1. When wind speed was 4.9–13.2 m·s−1, seed burial probability on the downwind slope was higher than that on the upwind slope. When wind speed was higher than 13.2 m·s−1, seed burial probability on the downwind slope was less than that on the upwind slope (Fig. 2b). Compared with the upwind slope without straw check boards, the straw check boards reduced probability of the seed burial when the wind speed was 3.4–10.7 m·s−1. However, when the wind speed was higher than 10.7 m·s−1, the straw check boards facilitated seed burial (Fig. 2b).

Comparison of seed burial probability on three sand surface configurations at different wind speeds. In the box plot (a), the thick line in the middle of the box indicates the median of each variable. The two ends of the box represent the higher and lower quartiles respectively. The upper and lower thin lines outside the box indicate the maximum and minimum values except the abnormal value. The “✳” indicates mean value, and the “★” indicates extreme value. In the polynomial regression diagram (b), the solid regression line corresponds to upwind slope (squares), short-dashed line to downwind slope (dots) and long-dashed line to upwind slope with barriers (triangles). yupwind = 0.313x2 − 1.939x + 2.897, R2 = 0.964, ydownwind = 0.257x2 − 0.925x − 0.744, R2 = 0.960, ybarrier = 0.485x2 − 4.364x + 9.110, R2 = 0.991
Seed burial probability in relation to diaspore traits
Of five diaspore traits, shape index, which was positively correlated with seed burial probability, and diaspore mass, which was negatively correlated with seed burial probability, were the two significant explanatory variables, explaining 6.1% (P < 0.01) and 0.7% (P < 0.05) variation, respectively (Tables 2 and 3). When wind speed was 3–9.5 m·s−1, the total contribution of diaspore traits to seed burial increased with an increase in wind speed but it decreased when the wind speed was ≥10.8 m·s−1 (Fig. 3, for details, see Supplementary Table S1). When wind speed was ≥6.9 m·s−1, the effect of diaspore traits on seed burial began to be significant. When wind speed was ≥6.9 m·s−1, shape index was positively correlated with seed burial probability. The correlation coefficient increased with an increase in wind speed, but it decreased when the wind speed was up to 12.1 m/s. When wind speed was 9.5 m·s−1, the probability of seed burial also had significant negative correlation with diaspore mass and projected area, but for the other wind speeds, no significant correlation existed (Table 3).

Explanations of impact factors to the total variation in seed burial probability under different wind speeds (For details, see Supplementary Table S1). MS: diaspore mass (mg), PA: projected area (mm2), SI: shape index, WL: wing loading (mg·mm−2), TV: terminal velocity (m·s−1)
Discussion
Although interactions of wind speed, land surface configuration and diaspore traits on seed burial during wind dispersal have been studied extensively, these data are the first empirical one resulting from a manipulative experiment involving multiple species by using a wind tunnel. Our results indicated that wind speed contributes the most to seed burial among the three factors. The influences of the slope direction on seed burial become more conspicuous and wind barriers facilitate seed burial only when the wind speed is high. Barriers can raise the wind speed threshold for seed burial. Effects of diaspore traits on seed burial are only significant at high wind speeds. Small or flat elongated diaspores are more likely to be buried than large or spherical diaspores.
Relative contributions of wind speed, diaspore traits, and sand surface configuration to seed burial
The wind speed, sand surface configuration, and diaspore traits are easily considered to be the affecting factors of soil seed bank formation (Thompson 1987; Horn et al. 2001; Li 2008). However, to determine which of these three aspects is the most important in controlling the burial of seeds, empirical and systematic studies are required. Based on a controlled wind tunnel experiment on 36 species with different diaspore traits, eight wind speeds, and three sand surface configurations, we found that wind speed is the most important for seed burial (Table 2). Previous studies indicated that wind conditions such as speed and type (advection or turbulence) can regulate primary seed dispersal (Horn et al. 2001). However, since our study took only wind speed into consideration, further study should take wind type into account.
Disperal modes reflect plant evolution selection. Anemochorous species have involved some morphological traits that facilitate wind dispersal (Hintze et al. 2013). In our study, we considered six wind-dispersal-related diaspore traits (quantitative or qualitative) in relation to diaspore traits in seed burial and found that diaspore traits are less important than wind speed in promoting burial. Thus, in general, environmental factors are more important in regulating soil seed bank formation than diaspore traits.
Presumably, the sand surface configuration should have a crucial influence on soil seed bank formation (Redbo-Torstensson and Telenius 1995). However, our results showed that in comparing with wind speed and diaspore traits, the sand surface configuration had no significant effect on seed burial. The possible reason for the result is that the three sand surface configurations, sand matrix, and gentle slopes are not sufficient for elucidating how sand surface configurations affect the burial of seeds. Further research is required to determine whether the results would change if other matrixes, steep slopes or other types of barriers were used.
Effects of sand surface configuration on seed burial
Although previous studies indicate that barriers can affect seed dispersal by regulating wind speed and surface roughness (Horvitz and Schemske 1986; Nathan and Muller-Landau 2000) and the difference in slope direction causes variance in seed deposition (Li 2008), there is a lack of exploration of the transient effects of slope direction and obstacles on seed burial.
Although sand surface configuration showed no significant effect on seed burial (Fig. 2a), the sand surface attributes such as the matrix, slope, barriers etc. as single factors have an impact on soil seed bank formation. Our observation indicates that the sand is more likely to be blown up on the downwind slope than on the upwind slope, and this might be the major cause of more diaspores being buried on the downwind slope than on the upwind slope. The barrier can weaken the near-surface wind speed (Zhang et al. 2006) and catch diaspores (unpublished observations); therefore, fewer seeds are able to be buried on the upwind slope with barriers.
Effects of sand surface configuration on seed burial are affected by wind speeds. The difference in the seed burial between slopes became more conspicuous and the barriers strengthened seed burial only when the wind speed is high (Fig. 2b). Low wind speed (< 5.9 m·s−1 in this study) is not enough to carry the sand to bury diaspores, but when the wind speed is high (> 13.2 m·s−1 in this study), turbulent flow seems more easily to be formed on the downwind slope than on the upwind slope. Thus, diaspores on the downwind slope are less easily buried than those on the upwind slope at high wind speeds. Previous studies indicated that the barriers on the soil surface can accelerate the formation of soil seed banks (Redbo-Torstensson and Telenius 1995). However, our study revealed that the effect of barriers on seed burial is modified by wind speed. It seems that there is wind speed threshold (10.7 m·s−1 in our study), above which barriers will not fuction well to stablize sand and diaspores easily will be buried.
Effects of diaspore traits on seed burial
Our study features the selection of multiple species and multiple indicators, as well as quantitative descriptions of diaspore attribute and seed burial. Previous studies usually selected several species in a single family (Mandák and Pyšek 2001; Moody-Weis and Alexander 2007; Zhu et al. 2016) or fewer morphological indicators (Peart 1981, Moles et al. 2000, Funes et al. 2003, Farrell et al. 2012). The selection of multiple species allows the results to reflect the general trends of seed bank formation. Quantitative multi-indicator selection can better screen decisive influencing factors.
In our results, shape index is the most important morphological trait affecting the burial of seeds by wind-blown sand (Table 3). The higher the shape index is (the diaspore tended to be flat elongated) the higher the probability of seeds becoming buried in sand. Diaspore mass is the secondary explanatory factor of soil seed bank formation, small diaspores are more likely to be buried than the large ones (Table 3). Diaspores with thorns or without appendages are more likely to be buried than those with hairs or wings; diaspores with hairs are especially difficult to bury (Table 2).
Our study indicated that the effects of diaspore traits on seed burial are regulated by wind speed. The explanatory power of diaspore attributes on seed burial increases with the wind speed, but decreases when the wind speed is particularly high (≥10.8 m·s−1 in this study) (Fig. 3). From the overall perspective, the soil seed bank is much easier to be formed from small and flat elongated diaspores than large and spherical ones (Table 3).
Conclusions
Our study suggests that the effect of diaspore traits on seed burial is modified by shifts in wind speed and sand surface configuration. This information has important implications on biodiversity conservation and ecological restoration. Seed dispersal and seed bank formation are closely linked to vegetation development and population maintenance (Jalili et al. 2003), and a seed bank can greatly facilitate restoration (Major and Pyott 1966). A reference for judging the effects of wind speed, diaspore traits, and sand surface configuration is provided. Although we cannot alter the wind speed in nature, we can modify the sand surface configuration and therefore increase soil seed bank formation, which could accelerate ecological restoration processes.
References
Aavik T, Helm A (2017) Restoration of plant species and genetic diversity depends on landscape-scale dispersal. Restor Ecol S2:S92–S102
Andersen MC (1993) Diaspore morphology and seed dispersal in several wind-dispersed Asteraceae. Am J Bot 80:487–492
Baker DV, Beck KG (2008) The weed tunnel: building an experimental wind tunnel. Weed Technol 22:549–552
Bakker JP, Poschlod P, Strykstra RJ, Bekker RM, Thompson K (1996) Seed banks and seed dispersal: important topics in restoration ecology. Plant Biol 45:461–490
Baskin CC and Baskin JM (2014) Chapter 7- germination ecology of seeds in the persistent seed bank. Seed ecology, biogeography, and evolution of dormancy and germination 187-276
Benvenuti S (2007) Natural weed seed burial: effect of soil texture, rain and seed characteristics. Seed Sci Res 17:211–219
Casper BB, Grant BW (1988) Morphology and dispersal of one and two-seeded diaspores of Cryptantha flava. Am J Bot 75:859–863
Casseau V, De Croon G, Izzo D, Pandolfi C (2015) Morphologic and aerodynamic considerations regarding the plumed seeds of Tragopogon pratensis and their implications for seed dispersal. PLoS One 10:e0125040
Chambers JC (1991) Seed entrapment in alpine ecosystems: effects of soil particle size and diaspore morphology. Ecology 72:1668–1677
Chambers JC (2000) Seed movements and seedling fates in disturbed sagebrush steppe ecosystems: implications for restoration. Ecol Appl 10:1400–1413
Chambers JC, Macmahon JA (1994) A day in the life of a seed: movements and fates of seeds and their implications for natural and managed systems. Annu Rev Ecol Syst 25:263–292
Chang ER, Veeneklaas RM, Buitenwerf R, Baker JP, Bouma TJ (2008) To move or not to move: determinants of seed retention in a tidal marsh. Functional ecology 22:720–727
Collins SL, Uno GE (1985) Seed predation, seed dispersal, and disturbance in grasslands: a comment. Am Nat 125:866–872
Driscoll DA, Banks SC, Barton PS, Karen I, Pia L, Lindenmayer DB et al (2014) The trajectory of dispersal research in conservation biology. Systematic review. PLoS ONE 9:e95053
Du X, Guo Q, Gao X, Ma KP (2007) Seed rain, soil seed bank, seed loss and regeneration of Castanopsis fargesii, (Fagaceae) in a subtropical evergreen broad-leaved forest. For Ecol Manag 238:212–219
Egawa C, Koyama A, Tsuyuzaki S (2009) Relationships between the developments of seedbank, standing vegetation and litter in a post-mined peatland. Plant Ecol 203:217–228
Farrell C, Hobbs RJ, Colmer TD (2012) Microsite and litter cover effects on seed banks vary with seed size and dispersal mechanisms: implications for revegetation of degraded saline land. Plant Ecol 213:1145–1155
Fenner M, Thompson K (2005) The ecology of seeds. Cambridge University Press, Cambridge
Funes G, Basconcelo S, Diaz S, Cabido M (1999) Seed size and shape are good predictors of seed persistence in soil in temperate mountain grasslands of Argentina. Seed Sci Res 9:341–345
Funes G, Basconcelo S, Sandra D, Cabido M (2003) Seed bank dynamics in tall-tussock grasslands along an altitudinal gradient. J Veg Sci 14:253–258
Green DS (1980) The terminal velocity and dispersal of spinning samaras. Am J Bot 67:1218–1224
Greene DF, Johnson EA (1997) Secondary dispersal of tree seeds on snow. J Ecol 85:329–340
Harper JL (1977) Population biology of plant. Academic Press, London
Helsen K, Hermy M, Honnay O (2015) Changes in the species and functional trait composition of the seed bank during semi-natural grassland assembly: seed bank disassembly or ecological palimpsest? J Veg Sci 26:58–67
Hintze C, Heydel F, Hoppe C, Cunze S, König A, Tackenberg O (2013) D3: the dispersal and diaspore database - baseline data and statistics on seed dispersal. Perspect Plant Ecol 15:180–192
Horn HS, Nathan R, Kaplan SR (2001) Long-distance dispersal of tree seeds by wind. Ecol Res 16:877–885
Horvitz CC, Schemske DW (1986) Seed dispersal and environmental heterogeneity in a neotropical herb: a model of population and patch dynamics. Frugivores and seed dispersal, Springer Netherlands
Howlett R (1995) Life in moving fluids - the physical biology of flow, 2nd edition - vogel,s. Nature 373:114–114
Jalili A, Hamzeh'Ee B, Asri Y et al (2003) Soil seed banks in the Arasbaran protected area of Iran and their significance for conservation management. Biol Conserv 109:425–431
Johnson EA, Fryer JI (1992) Physical characterization of seed microsites -- movement on the ground. J Ecol 80:823–836
Lemke A, Lippe MVD, Kowarik I (2009) New opportunities for an old method: using fluorescent colours to measure seed dispersal. J Appl Ecol 46:1122–1128
Li FR (2008) Presence of shrubs influences the spatial pattern of soil seed banks in desert herbaceous vegetation. J Veg Sci 19:537–548
López RP (2003) Soil seed banks in the semi-arid Prepuna of Bolivia. Plant Ecol 168:85–92
Maddox JC, Carlquist S (1985) Wind dispersal in Californian desert plants: experimental studies and conceptual considerations. Aliso 11:77–96
Major J, Pyott WT (1966) Buried viable seeds in temperate forest. In: White JD (ed) The population structure of vegetation. Junle Publishers, Dordrecht, pp 253–282
Maler A, Emig W, Leins P (1999) Dispersal patterns of some Phyteuma species (Campanulaceae). Plant Biol 1:408–417
Mandák B, Pyšek P (2001) Fruit dispersal and seed banks in Aatriplex sagittata: the role of heterocarpy. J Ecol 89:159–165
Mather JR (1987) Beaufort wind scale. In: Climatology. Encyclopedia of Earth Science. Springer, Boston, MA
Matlack GR (1987) Comparative demographies of four adjacent populations of the perennial herb Silene dioica (Caryophyllaceae). J Ecol 75:113–134
Matlack GR (1992) Influence of fruit size and weight on wind dispersal in Betula lenta, a gap-colonizing tree species. Am Midl Nat 128:30–39
Moles AT, Hodson DW, Webb CJ (2000) Seed size and shape and persistence in the soil in the New Zealand Flora. Oikos 89:541–545
Moody-Weis J, Alexander HM (2007) The mechanisms and consequences of seed bank formation in wild sunflowers (Helianthus annuus). J Ecol 95:14
Nathan R, Muller-Landau HC (2000) Spatial patterns of seed dispersal, their determinants and consequences for recruitment. Trends Ecol Evol 15:278–285
Nathan R, Schurr FM, Spiegel O, Steinitz O, Trakhtenbrot A, Tsoar A (2008) Mechanisms of long-distance seed dispersal. Trends Ecol Evol 23:0–647
Navie SC, Cowley RA, Rogers RW (1996) The relationship between distance from water and the soil seed bank in a grazed semi-arid subtropical rangeland. Aust J Bot 44:421–431
Obeso JR, Martínez I, García D (2011) Seed size is heterogeneously distributed among destination habitats in animal dispersed plants. Basic Appl Ecol 12:134–140
Oudtshoorn KVRV, Rooyen MWV (1999) Dispersal biology of desert plants. Springer, Berlin
Peart HM (1981) Further experiments on the biological significance of the morphology of seed-dispersal units in grasses. J Ecol 69:425
Peart HM (1984) The effects of morphology, orientation and position of grass diaspores on seedling survival. J Ecol 72:437–453
Pinceel T, Brendonck L, Vanschoenwinkel B (2016) Propagule size and shape may promote local wind dispersal in freshwater zooplankton -- a wind tunnel experiment. Limnol Oceanogr 61:122–131
Redbo-Torstensson P, Telenius A (1995) Primary and secondary seed dispersal by wind and water in Spergularia salina. Ecography 18:230–237
Rotundo JL, Aguiar MR, Benech-Arnold R (2015) Understanding erratic seedling emergence in perennial grasses using physiological models and field experimentation. Plant Ecol 216:143–156
Schurr FM, Bond WJ, Midgley GF, Higgins SI (2005) A mechanistic model for secondary seed dispersal by wind and its experimental validation. J Ecol 93:1017–1028
Simpson RL, Lerck MA, Parker VT (1989) Ecology of soil seed Banks. Academic Press, San Diego
Sletvold N, Rydgren K (2007) Population dynamics in Digitalis purpurea: the interaction of disturbance and seed bank dynamics. J Ecol 95:1346–1359
Sun Y, Tan DY, Baskin CC, Baskin JM (2012) Role of mucilage in seed dispersal and germination of the annual ephemeral Alyssum minus (Brassicaceae). Aust J Bot 60:439
Symes CT (2012) Seed dispersal and seed banks in Aloe marlothii (Asphodelaceae). S Afr J Bot 78:276–280
Tackenberg O (2003) Modeling long-distance dispersal of plant diaspores by wind. Ecol Monogr 73:173–189
Thompson K (1987) Seeds and seed banks. New Phytol 106:23–34
Thompson K, Grime JP (1979) Seasonal variation in the seed banks of herbaceous species in ten contrasting habitats. J Ecol 67:893–921
Thompson K, Band SR, Hodgson JG (1993) Seed size and shape predict persistance in soil. Funct Ecol 7:236–241
Touma JS (1977) Dependence of the wind profile power law on stability for various locations. J Air Pollut Contr Assoc 27:863–866
Vittoz P, Engler R (2007) Seed dispersal distances: a typology based on dispersal modes and plant traits. Bot Helv 117:109–124
Wang SM, Zhang X, Li Y, Zhang L, Xiong YC, Wang G (2005) Spatial distribution patterns of the soil seed bank of Stipagrostis pennata (Trin.) de winter in the Gurbantonggut Desert of north-West China. J Arid Environ 63:203–222
Willson MF, Rice BL, Westoby M (1990) Seed dispersal spectra: a comparison of temperate plant communities. J Veg Sci 1:547–562
Yan QL, Liu ZM, Zhu JJ, Luo YM, Wang HM, Jiang DM (2005) Structure, pattern and mechanisms of formation of seed banks in sand dune systems in northeastern Inner Mongolia, China. Plant Soil 277:175–184
Zhang RL, Liu GH, Cui XP (2006) Effects of living Salix gordejevii barrier on wind-breaking and sand-fixation in Hunshandake Sandland. J Desert Res 26:717–721
Zhu ZC, Cozzoli F, Chu NY et al (2016) Interactive effects between physical forces and ecosystem engineers on seed burial: a case study using Spartina anglica. Oikos 125:98–106
Zotz G, Weichgrebe T, Happatz H, Einzmann HJR (2016) Measuring the terminal velocity of tiny diaspores. Seed Sci Res 26:222–230
Acknowledgements
We thank Qun Ma, Liang Tian, Wenkai Shou, and Zhiquan Wang for their assistance in seed collection and field experiment. We also thank the Experimental Center of Desert Forestry, Chinese Academy of Forestry and the First Forestry Station for providing accommodations for us during the experiment. This study was supported financially by National Natural Science Foundation of China (41571270).
Author information
Authors and Affiliations
Contributions
Z.L., W.L., M.L., Z.W. and Z.S. conceived the ideas and designed methodology, W.L., X.Q., Q.Z., Z.X., and X.L. collected the data, Z.L., M.L., and Y.L. provided financial support, W.L. analyzed the data and led the writing of the manuscript, Z.L. revised this paper. All authors contributed critically to the drafts and gave final approval for publication.
Corresponding author
Additional information
Responsible Editor: Jeffrey Walck.
Publisher’s note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Highlight
The effect of diaspore traits on seed burial during wind dispersal is modified by the shifts in wind speed and sand surface configuration
Electronic supplementary material
ESM 1
(DOCX 21 kb)
Rights and permissions
About this article

Cite this article
Liang, W., Liu, Z., Liu, M. et al. How do diaspore traits, wind speed and sand surface configuration interact to determine seed burial during wind dispersal?. Plant Soil 440, 357–368 (2019). https://doi.org/10.1007/s11104-019-04071-4
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11104-019-04071-4