
Abstract
Aims
We examined the importance of litter quality and microclimate on early-stage litter mass loss, analysed the importance of interactions among environmental factors in determining key decomposition parameters and compared the variation in decomposition rates in vegetation types and sites with similar climate.
Methods
Following the Tea-Bag Index approach, 464 tea-bags were incubated in the soil in 79 sites, distributed across Italy, which included six vegetation types and a broad range of microclimatic conditions.
Results
Litter type exerted a stronger control on mass loss compared to climatic factors. The effects of soil moisture were not the same for high and lower quality litter. In addition, the effects of temperature on the decomposition rate depended on soil moisture. The stabilization factor was strongly temperature-dependent, but the influence of temperature differed among vegetation types: those dominated by small-size plants showed a strong decrease in the potential amount of plant material entering into the soil stock under warmer temperatures. The lowest variation in decomposition rate was found in sites characterised by low temperatures, and, among the vegetation types, in alpine snowbeds.
Conclusions
The role of litter quality and of the interactions among environmental conditions can potentially determine significant shifts in the expected patterns of ecosystem carbon fluxes.
Similar content being viewed by others


Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
Decomposition processes play a key role in linking organic and inorganic components of natural ecosystems and are equally significant in the completion of the carbon cycle. Litter decomposition represents a fundamental component in such processes since plants, as primary producers, are the main source of organic carbon in terrestrial ecosystems.
Decomposition has been widely studied for over a century, but it was the introduction of the litter bags technique (Bocock and Gilbert 1957) that triggered the proliferation of a wide range of systematic studies, which were also motivated by the growing concern about global climate change (e.g. Cornwell et al. 2008). Research undertaken in recent decades has tried to identify the main drivers of litter decomposition as well as highlighting the importance of such factors across a wide range of environmental conditions at different spatial scales (e.g. Gholz et al. 2000; Cornelissen et al. 2007; García-Palacios et al. 2013, 2016; Parton et al. 2007; Zhang et al. 2008). This research identifies climatic factors and litter quality as key drivers of litter decomposition on both a regional and global scale (Aerts 1997; Austin and Vitousek 2000; Cornwell et al. 2008; García-Palacios et al. 2013; Zhang et al. 2008).
Climate can affect litter decomposition directly, by regulating the activity of decomposers, and indirectly through changes in plant species composition and abundance and, as a result, in variation in litter quality and quantity. The decomposability of the litter depends also on the specific physical and chemical properties of plant tissues (i.e. their quality as a resource for decomposers) due to their responsiveness to microbial mineralization (Jagadamma et al. 2014). However, the lack of a widespread implementation of an easy-to-use and well-standardized method for estimating litter decomposition has hampered investigations aimed at understanding the relative importance of climatic factors and litter quality. In order to overcome the conceptual and practical setbacks involved in the use of local litter, Keuskamp et al. (2013) have recently proposed the use of the Tea-Bag Index (TBI), a method based on commercially available tea as a standard plant material. The TBI can be a useful tool to increase understanding of decomposition processes through an assessment of the relative importance of different drivers and through pinpointing the role of the interactions among environmental factors.
The function of the principal climatic factors (i.e. temperature and precipitation) and litter quality has been investigated independently in the past. The potential interactions among these agents is poorly known, despite their potential importance in the carbon balance of terrestrial ecosystems (Wu et al. 2011), in particular under changing climatic conditions and vegetation dynamics. Although the importance of the interactions between temperature and soil moisture on soil respiration rates has recently been demonstrated (Sierra et al. 2017; Tucker and Reed 2016; Wang et al. 2016), research on the role of the interactions among drivers of litter decomposition processes (i.e. litter quality, vegetation properties and climatic features) remains limited. The few available studies on the influence of interactions among environmental factors on litter decomposition are based on the interaction between mean annual air temperature (MAT) and mean annual cumulative precipitation (MAP) (Taylor et al. 2017) and on the interactions between litter quality and MAT, MAP, land-use and biome (Djukic et al. 2018). Furthermore, in most studies on litter decomposition, the main climatic variables tested were treated as constant across large areas and within biomes (e.g. Cornelissen et al. 2007; Cornwell et al. 2008). Topographic variability and vegetation cover, however, should be taken into consideration as these factors can potentially influence local microclimatic conditions (e.g. Wundram et al. 2010; Graham et al. 2012) and their spatial variation occurs at finer scales compared to the ones provided by high-resolution gridded climatic datasets. Variations in soil slope and aspect coupled with plant shading and transpiration are able to determine detailed patterns of soil temperature and moisture conditions on a very small spatial scale. In addition, historical land-use coupled with the occurrence of azonal vegetation types such as those associated with specific site conditions (e.g. waterlogging, high salt content, long-lasting snow-cover, frequent disturbance) contribute to a substantial plant cover differentiation of the landscape. It is, therefore, unknown how much variation in litter decomposition occurs at a local level, such as in sites characterised by similar climate but with different vegetation types.
We measured early-stage mass loss and decomposition parameters following the TBI approach (Keuskamp et al. 2013) in order to investigate the effects of litter quality, microclimatic conditions, vegetation types and their interactions on litter decomposition and also to estimate the variation occurring in sites with similar climate and among different vegetation types. This was achieved by calculating the decomposition rate constant k, representing the turnover time of the labile fraction of material (i.e. the short-term dynamics of new inputs) and the stabilization factor S, indicating the amount of labile material that becomes recalcitrant (i.e. the potential carbon storage).
In particular, the aims of the present study were: (i) to assess the relative importance of litter quality and soil temperature and moisture in determining early-stage mass loss of litter; (ii) to disentangle the effects of vegetation type and soil temperature and moisture on decomposition parameters; and (iii) to compare the variation in decomposition rate among types of vegetation and among group of sites characterised by different climatic conditions.
Materials and methods
Study sites and environmental data
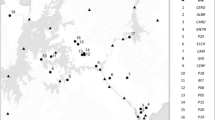
The study was carried out in 79 sites distributed all over Italy (Fig. 1). Given the position of the Italian peninsula, laying in the temperate zone of the Northern Hemisphere but located in the middle of the Mediterranean basin, together with its orographic features, Italy provides a wide range of contrasting climatic regimes within relatively small areas. Moreover, the historical influence of human land-use offers a variety of natural and semi-natural habitats within short distances and, consequently, experiencing the same climatic regime. The study sites were located at an elevation ranging from 0 to 2681 m a.s.l. and comprised different zones, ranging from the coastal areas of the southern peninsular regions and Sardinia, characterised by a typical Mediterranean climate, to the alpine tundra environment in the Rhaetian Alps (Table 1).

Location of the study sites, with vegetation type and soil moisture class. Tea-bag silhouette has the sole purpose of graphically representing the position and the number of the study sites occurring in neighbouring locations (small size <6, medium size = 6 to 10, large size >10)
Each site was assigned one of six vegetation types - snowbed, wetland, grassland, shrubland, broad-leaved forest, coniferous forest – on the basis of the composition and structure of its plant communities.
Soil temperature was recorded hourly at each site by a Pendant sensor (Onset, Cape Cod, MA, USA) placed in the soil at a depth of 8 cm, while soil water content was estimated through direct observations of the physical features of the soil - i.e. friability, ductility, waterlogging - carried out during the burying of the tea-bags in the soil and their subsequent retrieval. An estimate of the percentage of gravimetric water content (GWC) of the soil was undertaken in order to assign a categorical class of soil moisture to each site. Three categories of soil moisture were identified: dry (roughly <20% GWC), moist (20–80% GWC) and wet (GWC > 80%).
Standard material and sampling design
Tea-bags were incubated and processed following the Keuskamp et al. (2013) protocol. In each site, 3 to 5 sets of tea-bags, each set consisting of one green and one rooibos tea type, were buried in the soil at 8 cm depth at the beginning of the summer 2016 or 2017, depending on the site, and retrieved after approx. 3 months (Table 1). Lipton green tea (EAN: 87 22,700 05552 5) and Lipton rooibos tea (EAN: 87 22,700 18,843 8) were used, composed of nylon bags with a mesh size of 0.25 mm. Each bag contained approx. 2 g of tea. The two varieties of tea differ in the type of plant material used, C:N ratio and percentage of water soluble fraction. Whereas green tea consists of leaves, has a C:N ratio of approx. 12 and a high water soluble fraction (ca. 50%), rooibos tea consists of a mixture of mainly needle-like litter and stem tissue, with a C:N ratio of approx. 43 and contains half the amount of soluble compounds of green tea (Keuskamp et al. 2013). Because of their chemical properties, the two types of tea were used as surrogates of higher and lower quality litter, respectively. After the exclusion of damaged material, the data set included a total of 464 tea-bags, for each of which the initial air-dried mass and the final oven-dry mass was measured. An independent set of tea-bags, consisting of 26 green and 26 rooibos tea-bags, was used to estimate the initial oven-dry mass by calculating the ratio between air- and oven-dry masses.
Estimation of litter mass loss and decomposition parameters (k and S)
Litter mass loss was calculated for both green and rooibos tea as:
where M0 and M1 are the initial and final oven-dry mass of the tea, respectively.
In addition, two decomposition parameters were estimated following the TBI approach (Keuskamp et al. 2013): the decomposition rate constant of the labile fraction (k) and the stabilization factor (S). The TBI approach is based on the assumption that the mass loss of the recalcitrant fraction of plant material is negligible during short incubation periods (i.e. ca. 3 months). Therefore, the double exponential model was expressed as:
where X is the fraction of remaining mass at time t, a is the labile fraction, (1 – a) is the recalcitrant fraction of the material and k is the decomposition rate constant of the labile fraction. The latter was estimated as:
where Xr is the fraction of remaining rooibos tea (i.e. M1 / M0), ar is the predicted labile fraction of rooibos tea and t is the incubation time, expressed in days. The rooibos tea labile fraction was calculated as:
where Hr is the hydrolysable fraction of rooibos tea and S is the stabilization factor, which is assumed to be the same for both tea types and can be interpreted as the inhibiting effect of environment on the decomposition of the labile fraction of litter (Keuskamp et al. 2013). The stabilization factor, which indicates the amount of labile materials that tends to stabilize becoming recalcitrant, was calculated as:
where ag and Hg are respectively the decomposable fraction and hydrolysable fraction of green tea.
Statistical analyses
A linear mixed-effect model (LMM) was performed to assess the effects of litter quality and soil temperature and moisture on initial (3-month period) mass loss. The proportion of mass loss was set as the response variable, whereas the tea type (2-level factor: green and rooibos tea, for higher and lower quality litter, respectively), the average soil temperature during the incubation period (continuous variable, in °C), the soil moisture class (3-level factor: dry, moist and wet) and their 2-way interactions were considered as predictors. Since we buried multiple sets of teabags in each site, site identity was included in the model as a random factor. For this and the following analyses, minimal adequate models were obtained by means of model selection following Crawley (2013), and model assumptions were checked through visual inspection of residual patterns (Zuur et al. 2009).
LMMs were then fitted to k and S, considered as response variables, to investigate whether the decomposition parameters differ among vegetation types and how these are influenced by soil temperature and moisture. In both models, the vegetation type (6-level factor: snowbed, wetland, grassland, shrubland, broad-leaved forest and coniferous forest), soil temperature and moisture class and their 2-way interactions were set as the fixed effects, whereas the site was set as the random effect. To meet linear model assumptions, k values were square root transformed prior to analysis.
Finally, differences in the coefficient of variation (CV) of k among climatic clusters and among vegetation types were analysed using the asymptotic test of Feltz and Miller (1996). Climatic clusters were defined through a cluster analysis on mean air temperature and cumulative precipitation data of the warmest quarter of the year during the period 1970–2000; climatic variables were extracted from WorldClim version 2 (Fick and Hijmans 2017) at a 30 s (~1 km2) spatial resolution. Cluster analysis was performed with the Ward (minimum variance) clustering method and the Euclidean dissimilarity index. Finally, the optimal number of clusters was identified following the silhouette width criterion (Rousseeuw 1987).
Statistical analyses were carried out in R version 3.4.3 (R Core Team 2017) with the following packages: lme4 (Bates et al. 2015) for model fitting, car (Fox and Weisberg 2011) for model selection, multcomp (Hothorn et al. 2008) for post-hoc comparisons, vegan (Oksanen et al. 2017) for cluster analysis, cvequality (Marwick and Krishnamoorthy 2018) for comparison of CVs and visreg (Breheny and Burchett 2017) for graphs of regressions.
Results
Mass loss
Results illustrated the following hierarchy regarding the drivers which determine mass loss: litter quality (F1,382 = 6110.30, P < 0.001) >> > soil moisture (F1,75 = 10.49, P < 0.001) > soil temperature (F1,74 = 7.54, P = 0.008). Thus, variation due to different litter quality was significantly greater than that due to soil temperature (Fig. 2a) and moisture (Fig. 2b) variation. The interaction between litter type and soil moisture was also significant (F2,382 = 4.75, P = 0.009), with wetter soil conditions promoting the mass loss of higher litter quality to a greater extent compared to the mass loss of lower litter quality. Overall, mass loss was higher in wet and moist soils compared to dry ones (Z = 4.445, P < 0.001 and Z = 3.183, P < 0.001, respectively), while the difference between wet and moist soils was marginally significant (Z = 2.204, P = 0.069).

Mass loss of higher and lower quality litter in relation to (a) temperature variation and (b) soil moisture classes
Decomposition parameters
The analysis did not evidence an overall effect of soil temperature on the decomposition constant k (F1,72 = 1.93, P = 0.169), which, on the other hand, exhibited a significant response to both soil moisture (F2,73 = 6.36, P = 0.003) and the interaction between soil moisture and temperature (F2,73 = 8.39, P < 0.001) (Fig. 3a). Hence, the effects of soil temperature and moisture were not additive and increasing temperatures were associated to higher k values only in dry and wet soils. On average, k values were higher in wet soils than in dry (Z = 4.338, P < 0.001) and moist soils (Z = 3.914, P < 0.001), whereas dry and moist soils did not differ between each other (Z = 1.122, P = 0.497). The vegetation type did not exhibit a significant influence on k (variable excluded from the model).

Relationships between (a) the decomposition constant and (b) the stabilization factor with temperatures and moisture classes of the soil
The stabilization factor S was significantly affected by both soil temperature (F1,65 = 8.16, P = 0.006) and moisture (F1,67 = 4.73, P = 0.012) (Fig. 3b), while differences among vegetation types were marginally significant (F1,65 = 2.07, P = 0.080). Overall, dry sites exhibited a higher S than moist ones (Z = −2.894, P = 0.010). Moreover, the effects of temperature and moisture were additive (interaction excluded from the model), unlike those of temperature and vegetation type (F1,67 = 5.22, P < 0.001) (Fig. 4). In particular, a significant decrease of S occurred with increasing temperatures in snowbeds, wetlands and grasslands, whereas in vegetation dominated by shrubs or trees S did not vary at different temperatures.

Effects of soil temperatures on the stabilisation factor in different vegetation types
Variation in decomposition rate
The coefficient of variation (CV) showed significant differences both among the 17 climatic clusters and among the six vegetation types (D’AD = 57.5, P < 0.001 and D’AD = 23.8, P < 0.001, respectively). Despite substantial variation occurring in almost all the clusters (Fig. S1 in Supplementary Material), the CV seemed to follow a trend towards lower values at colder temperatures (Fig. 5a) and higher values at high temperatures and low precipitation. Finally, among the vegetation types investigated, the lowest CV of k was found in snowbed communities (Fig. 5b), whereas grasslands and coniferous forests showed the highest variation in the decomposition rate.

Values of the coefficient of variation in relation to site-specific temperature and precipitation extracted from the WorldClim dataset (Fick and Hijmans 2017) and in the six vegetation types
Discussion
Studies on decomposition processes based on standard litter allow the investigation of environmental drivers without being conditioned by marginal effects such as the “home-field advantage” (Gholz et al. 2000). Based on incubations performed in a wide range of environmental conditions, the present study demonstrated that litter quality exerts a stronger effect on early-stage mass loss than variation of soil temperature and moisture, supporting the conclusions of previous studies (Carbognani et al. 2014; Cornwell et al. 2008; Djukic et al. 2018; Shaw and Harte 2001; Zhang et al. 2008). Furthermore, the results suggested that the differences in chemical and physical properties of litter are more important in determining mass loss compared to the variation in biological processes, such as microbial respiration induced by increasing temperature (Fig. 2a) and the variation of abiotic processes, such as leaching induced by soil water availability (Fig. 2b).
The analysis of the interplay of controlling factors revealed the occurrence of three significant interactions: (i) between litter quality and soil moisture, affecting mass loss (Fig. 2b); (ii) between soil temperature and moisture, influencing the decomposition constant k (Fig. 3a); and (iii) between soil temperature and vegetation type, acting on the stabilization factor S (Fig. 4).
With regard to the first interaction, although responses of mass loss to temperature in green and rooibos tea were similar, the two litter types did not decompose equally in soils with different moisture content: the difference in mass loss of high quality litter in sites with different soil moisture (i.e. dry vs moist and wet sites) was higher compared to the mass loss of low quality litter (Fig. 2b). This result is consistent with the conclusions of Yajun et al. (2016), indicating that the magnitude of synergistic interactions between soil water content and litter type increases with increasing water availability. Liu et al. (2005) also showed that water addition favours mass loss in high quality litter. The greater sensitivity of high quality litter in the leaching phase could explain the higher responsiveness of mass loss to increasing soil water content in this type of litter compared to low quality litter. Indeed, a substantial part of litter mass loss during the early stages of litter decomposition occurs during the leaching phase, when both inorganic elements and simple organic compounds are removed (MacLean and Wein 1978). Given the importance of the water-driven phase on litter decomposition and its dependence on both the quantity and quality of water-soluble compounds (Ibrahima et al. 2008), it could be asserted that high soil water content can potentially determine a stronger mass loss in litter types rich in carbohydrates, these being easily leached during the first decomposition stages (Cotrufo et al. 2015; Mansfield and Bärlocher 2005). Liu et al. (2005) suggested that the different responses of the two litter types to soil water content could also be related to their different physical structures: while, in the case of rooibos tea coriaceous and lignified leaves are present, green tea is composed of softer and more fragile leaves, which could be more prone to physical fragmentation and leaching.
The litter decomposition rate, driven mainly by microbial activity, is largely temperature-dependent (Davidson and Janssens 2006; Kirschbaum 2006). The results revealed, however, that a significant increment in k was determined by warmer soil temperatures only in dry and wet sites, whereas in moist soils the decomposition rate did not exhibit any significant response to temperature variation (Fig. 3a). The impact of temperature on k is, therefore, not consistent in soils with different moisture levels. Results from both green and rooibos tea (Fig. 2b) showed that dry soils are characterized by a lower mass loss than moist and wet soils. It seems, therefore, that the reduction of the decomposition rate associated with drier conditions counteracts the enhancing effect of temperature on microbial activity (Fig. 2a), resulting in no significant increase of decomposition rates with increasing temperatures in moist conditions (Fig. 3a). A possible explanation of these results could be that, although a temperature increase enhances the activity of decomposers, it also reduces the moisture of the soil. It is likely that in intermediate soil moisture conditions (i.e. GWC ranging from 20 to 80%) warmer temperatures may cause larger differences in soil moisture compared to those occurring in dry and wet soils. Similarly, Christiansen et al. (2016) reported a negative relation between litter decomposition rates and temperature increase in both xeric and wet tundra sites, due to evaporative drying associated with warmer temperatures, which counteracted the enhancing effect of temperature on microbial activity. Alternatively, the vegetation type could also explain the lack of response of k to temperature variation in soils with intermediate water content, since most of the sites with soil classified as moist were grasslands, a vegetation type that has already been found to exhibit unexpected responses to warming. In the study by Bontti et al. (2009) litter decomposition in grasslands was not shown to be affected by any of the climatic variables under consideration.
In contrast to the decomposition constant k, the stabilization factor S was significantly reduced by both increasing soil temperature and moisture, with the effects of the two variables resulting as additive - i.e. temperature has the same effect in soils belonging to different moisture classes. These results are consistent with those reported by Mueller et al. (2018) for tidal wetlands, indicating that the stabilization of organic material is higher in colder and drier soils. In the present research, the vegetation type did not seem to exhibit any direct control on early decomposition rates, confirming the results of Djukic et al. (2018) on land use categories. The absence of a direct effect with regard to the vegetation type on litter decomposition dynamics could be explained by the greater influence of climatic factors, with values of temperature and moisture varying greatly among vegetation types. However, the effect of the interaction between vegetation type and temperature on the stabilization factor S was highly significant (Fig. 4). This suggests that warming-induced effects on the potential storage of organic carbon in the soil could differ among vegetation types: although S consistently decreased with increasing temperature in all soil moisture categories (Fig. 3b), consistent patterns across all the considered vegetation types were not observed (Fig. 4). Whereas soil temperature in shrublands and forests did not affect S, the stabilization factor was significantly reduced by warmer temperatures in snowbeds, wetlands and grasslands, which may suggest a lower stability of the soil carbon stocks of these vegetation types under warmer climatic conditions. More specifically, the lack of temperature sensitivity of S in shrublands and forests is not consistent with the well-known warming-induced increase in decomposition of soil organic carbon (SOC) that can strongly affect the ecosystem carbon storage (e.g. Ding et al. 2014; Melillo et al. 2011; Trumbore et al. 1996). Furthermore, in a study on the drivers of SOC stability in temperate forests, Tian et al. (2016) reported that MAT only influences the labile carbon pool size but does not affect the SOC stability. Another study by Crowther et al. (2016) showed that the sensitivity of soil carbon stock to warming strictly depends on its initial size. Our results seems consistent with those reported by this last study since wetlands, snowbeds and mountain grasslands generally hold a large amount of SOC (e.g. Garcia-Pausas et al. 2017). Our findings could also help in predicting in which ecosystems the SOC might be more sensitive to the current warming trend: in particular, peatlands (i.e. wetlands) and alpine tundra (i.e. snowbeds) communities seem to be the most sensitive to warming-induced changes in carbon fluxes, confirming the long-held concern about possible positive feedbacks on climate warming (Conant et al. 2011; Davidson and Janssens 2006; Kirschbaum 2006). Considering that significant influences of warmer temperatures were found in vegetation types characterised by small-size plants (i.e. snowbeds, wetlands and grasslands), it is likely that the effects of climate warming on decomposition could be especially pronounced in sites with reduced vegetation cover or small plant size, where solar radiation can warm the soil without being screened by the canopy. The results lead to the conclusion that the vegetation type could effectively modulate the impact of temperature increase on ecosystem carbon stocks and should, therefore, be taken into consideration when modelling future scenarios of carbon cycle responses to climate change.
It is also important to note that local microclimatic conditions are also influenced by further environmental factors other than temperature and precipitation characterising regional climate regimes. Topography and evapotranspiration can, in fact, determine a substantial variation in the decomposition rate k, as suggested by the analysis along the climatic gradients investigated in the present study (Fig. 5a). The variation in k was generally high both among sites characterised by similar climatic conditions and among similar vegetation types. Low variation of k was found in the coldest sites, characterised by alpine tundra vegetation (i.e. snowbeds) and in vegetation types associated with high level of soil moisture (i.e. wetlands) (Fig. 5b). Among the investigated vegetation types, grasslands and coniferous forests had the highest variation in decomposition rate, probably due to the high range of climatic conditions where these types of vegetation can develop, with grasslands being present along a wide elevation gradient (from 0 to 2654 m a.s.l.) and coniferous forests covering a broad latitudinal range (from 39.2° to 46.3° N).
Conclusions
The Tea-Bag Index, allowing the testing of litter decomposition with a replicable standard over a wide range of environments, has proved to be an effective method for studying the drivers of litter decomposition. The present study provides evidence that litter quality not only exerts the strongest influence on early litter decomposition dynamics, but also modulates the effect of soil moisture on mass loss. Moreover, while the temperature effect on the decomposition constant k depends on soil moisture, the warming-induced decrease of the amount of organic material accumulated in the soil is constant at varying levels of moisture. The temperature effect appears, however, to be related to the vegetation type, with the stabilization factor in colder and wetter ecosystems, such as snowbeds and wetlands, being potentially more sensitive to current climate change. In addition, when gridded climatic factors were used as predictors, substantial variation in the decomposition rate was revealed, as well as in types of vegetation occurring over wide environmental gradients. In the light of these results, the variability in the response of carbon stock to climatic drivers as a function of climatic conditions and vegetation types should be taken into consideration when modelling future scenarios of carbon fluxes across terrestrial ecosystems.
Abbreviations
- GWC:
-
Gravimetric water content
- LMM:
-
Linear mixed-effect model
- MAP:
-
Mean annual cumulative precipitation
- MAT:
-
Mean annual air temperature
- SOC:
-
Soil Organic Carbon
- TBI:
-
Tea-Bag Index
References
Aerts R (1997) Climate, leaf litter chemistry and leaf litter decomposition in terrestrial ecosystems: a triangular relationship. Oikos 79:439–449. https://doi.org/10.2307/3546886
Austin AT, Vitousek PM (2000) Precipitation, decomposition and litter decomposability of Metrosideros polymorpha in native forests on Hawai'i. J Ecol 88:129–138. https://doi.org/10.1046/j.1365-2745.2000.00437.x
Bates D, Maechler M, Bolker B, Walker S (2015) Fitting linear mixed-effects models using lme4. J Stat Softw 67:1–48. https://doi.org/10.18637/jss.v067.i01
Bocock KL, Gilbert OJW (1957) The disappearance of leaf litter under different woodland conditions. Plant Soil 9:179–185. https://doi.org/10.1007/BF01398924
Bontti EE, Decant JP, Munson SM, Gathany MA, Przeszlowska A, Haddix ML, Owens S, Burke IC, Parton WJ, Harmons ME (2009) Litter decomposition in grasslands of Central North America (US Great Plains). Glob Change Biol 15:1356–1363. https://doi.org/10.1111/j.1365-2486.2008.01815.x
Breheny P, Burchett W (2017) Visualization of regression models using visreg. The R Journal 9:56–71
Carbognani M, Petraglia A, Tomaselli M (2014) Warming effects and plant trait control on the early-decomposition in alpine snowbeds. Plant Soil 376:277–290. https://doi.org/10.1007/s11104-013-1982-8
Christiansen CT, Haugwitz MS, Priem A, Nielsen CS, Elberling B, Michelsen A, Grogan P, Blok D (2016) Enhanced summer warming reduces fungal decomposer diversity and litter mass loss more strongly in dry than in wet tundra. Glob Change Biol 23:406–420. https://doi.org/10.1111/gcb.13362
Conant RT, Ryan MG, Agren GI et al (2011) Temperature and soil organic matter decomposition rates – synthesis of current knowledge and a way forward. Glob Change Biol 17:3392–3404. https://doi.org/10.1111/j.1365-2486.2011.02496.x
Cornelissen JHC, van Bodegom PM, Aerts R et al (2007) Global negative vegetation feedback to climate warming responses of leaf litter decomposition rates in cold biomes. Ecol Lett 10:619–627. https://doi.org/10.1111/j.1461-0248.2007.01051.x
Cornwell WK, Cornelissen JHC, Amatangelo K, Dorrepaal E, Eviner VT, Godoy O, Hobbie SE (2008) Plant species traits are the predominant control on litter decomposition rates within biomes worldwide. Ecol Lett 11:1065–1071. https://doi.org/10.1111/j.1461-0248.2008.01219.x
Cotrufo MF, Soong JL, Horton AJ, Campbell EE, Haddix ML, Wall DH, Parton WJ (2015) Formation of soil organic matter via biochemical and physical pathways of litter mass loss. Nat Geosci 8:776–779. https://doi.org/10.1038/ngeo2520
Crawley MJ (2013) The R book. Wiley, Chichester. https://doi.org/10.1002/9781118448908
Crowther TW, Todd-Brown KEO, Rowe CW et al (2016) Quantifying global soil carbon losses in response to warming. Nature 540:104–110. https://doi.org/10.1038/nature20150
Davidson EA, Janssens IA (2006) Temperature sensitivity of soil carbon decomposition and feedbacks to climate change. Nature 440:165–173. https://doi.org/10.1038/nature04514
Ding F, Huang Y, Sun W, Jiang G, Chen Y (2014) Decomposition of organic carbon in fine soil particles is likely more sensitive to warming than in coarse particles: an incubation study with temperate grassland and forest soils in northern China. PLoS One 9:e95348. https://doi.org/10.1371/journal.pone.0095348
Djukic I, Kepfer-Rojas S, Schmidt IK et al (2018) Early stage litter decomposition across biomes. Sci Total Environ 628-629:1369–1394. https://doi.org/10.1016/j.scitotenv.2018.01.012
Feltz CJ, Miller GE (1996) An asymptotic test for the equality of coefficients of variation from k populations. Stat Med 15:647–658. https://doi.org/10.1002/(SICI)1097-0258(19960330)15:6<647::AIDSIM184>3.0.CO;2-P
Fick SE, Hijmans RJ (2017) Worldclim 2: new 1-km spatial resolution climate surfaces for global land areas. Int J Climatol 37:4302–4315. https://doi.org/10.1002/joc.5086
Fox J, Weisberg S (2011) An {R} companion to applied regression. Sage, Thousand Oaks
García-Palacios P, Maestre FT, Kattge J, Wall DH (2013) Climate and litter quality differently modulate the effects of soil fauna on litter decomposition across biomes. Ecol Lett 16:1045–1053. https://doi.org/10.1111/ele.12137
García-Palacios P, McKie BG, Handa IT, Frainer A, Hättenschwiler S (2016) The importance of litter traits and decomposers for litter decomposition: a comparison of aquatic and terrestrial ecosystems within and across biomes. Funct Ecol 30:819–829. https://doi.org/10.1111/1365-2435.12589
Garcia-Pausas J, Montané F, Rios AI, Taull M, Rovira P, Casals P (2017) Are soil carbon stocks in mountain grasslands compromised by land-use changes? In: Catalan J, Ninot JM, Aniz M (eds) High mountain conservation in a changing world. Springer Nature, Cham, pp 207–230. https://doi.org/10.1007/978-3-319-55982-7_9
Gholz HL, Wedin DA, Smitherman SM, Harmon ME, Parton WJ (2000) Long-term dynamics of pine and hardwood litter in contrasting environments: toward a global model of decomposition. Glob Change Biol 6:751–765. https://doi.org/10.1046/j.1365-2486.2000.00349.x
Graham EA, Rundel PW, Kaiser W, Lam Y, Stealey M, Yuen EM (2012) Fine-scale patterns of soil and plant surface temperatures in an alpine fellfield habitat, White Mountains, California. Arct Antarct Alp Res 44:288–295. https://doi.org/10.1657/1938-4246-44.3.288
Hothorn T, Bretz F, Westfall P (2008) Simultaneous inference in general parametric models. Biom J 50:346–363. https://doi.org/10.1002/bimj.200810425
Ibrahima A, Biyanzi P, Halima M (2008) Changes in organic compounds during leaf litter leaching: laboratory experiment on eight plant species of the Sudano-Guinea savannas of Ngaoundere, Cameroon. iForest 1:27–33. https://doi.org/10.3832/ifor0450-0010027
Jagadamma S, Mayes MA, Steinweg JM, Schaeffer SM (2014) Substrate quality alters the microbial mineralization of added substrate and soil organic carbon. Biogeosciences 11:4665–4678. https://doi.org/10.5194/bg-11-4665-2014
Keuskamp JA, Dingemans BJJ, Lehtinen T, Sarneel JM, Hefting MM (2013) Tea bag index: a novel approach to collect uniform decomposition data across ecosystems. Methods Ecol Evol 4:1070–1075. https://doi.org/10.1111/2041-210X.12097
Kirschbaum MUF (2006) The temperature dependence of organic-matter decomposition – still a topic of debate. Soil Biol Biochem 38:2510–2518. https://doi.org/10.1016/j.soilbio.2006.01.030
Liu P, Huang J, Han X, Sun OJ, Zhou Z (2005) Differential responses of litter decomposition to increased soil nutrients and water between two contrasting grassland plant species of Inner Mongolia. China Appl Soil Ecol 34:266–275. https://doi.org/10.1016/j.apsoil.2005.12.009
MacLean DA, Wein RW (1978) Weight loss and nutrient changes in decomposing litter and forest floor material in New Brunswick forest stands. Can J Botany 56:2730–2749. https://doi.org/10.1139/b78-326
Mansfield SD, Bärlocher F (2005) Determination of soluble carbohydrates. In: Graça MAS, Bärlocher F, Gessner MO (eds) Methods to study litter decomposition. Springer, Dordrecht, pp 85–90. https://doi.org/10.1007/1-4020-3466-0_12
Marwick B, Krishnamoorthy K (2018) cvequality: tests for the equality of coefficients of variation from multiple groups. R software package version 01.3. https://github.com/benmarwick/cvequality
Melillo JM, Butler S, Johnson J et al (2011) Soil warming, carbon–nitrogen interactions, and forest carbon budgets. P Natl Acad Sci USA 108:9508–9512. https://doi.org/10.1073/pnas.1018189108
Mueller P, Schile-Beers LM, Mozdzer TJ et al (2018) Global-change effects on early-stage decomposition processes in tidal wetlands – implications from a global survey using standardized litter. Biogeosciences 15:3189–3202. https://doi.org/10.5194/bg-15-3189-2018
Oksanen J, Blanchet FG, Friendly M, Kindt R, Legendre P, McGlinn D, Minchin PR, O'Hara RB, Simpson GL, Solymos P, Stevens MHH, Szoecs E, Wagner H (2017) Vegan: community ecology package. R package version 2.4–4. https://CRAN.R-project.org/package=vegan
Parton W, Silver WL, Burke IC et al (2007) Global-scale similarities in nitrogen release patterns during long-term decomposition. Science 315:361–364. https://doi.org/10.1126/science.1134853
R Core Team (2017) R: a language and environment for statistical computing. The R Foundation for Statistical Computing, Vienna. http://www.R-project.org/
Rousseeuw PJ (1987) Silhouettes: a graphical aid to the interpretation and validation of cluster analysis. J Comput Appl Math 20:53–65. https://doi.org/10.1016/0377-0427(87)90125-7
Shaw MR, Harte J (2001) Control of litter decomposition in a subalpine meadow–sagebrush steppe ecotone under climate change. Ecol Appl 11:1206–1223. https://doi.org/10.2307/3061022
Sierra CA, Malghani S, Loescher HW (2017) Interactions among temperature, moisture, and oxygen concentrations in controlling decomposition rates in a boreal forest soil. Biogeosciences 14:703–710. https://doi.org/10.5194/bg-14-703-2017
Taylor PG, Cleveland CC, Wieder WR, Sullivan BW, Doughty CE, Doughty SZ, Townsend AR (2017) Temperature and rainfall interact to control carbon cycling in tropical forests. Ecol Lett 20:779–788. https://doi.org/10.1111/ele.12765
Tian Q, He H, Cheng W, Bai Z, Wang Y, Zhang X (2016) Factors controlling soil organic carbon stability along a temperate forest altitudinal gradient. Sci Rep-UK 6:18783. https://doi.org/10.1038/srep18783
Trumbore SE, Chadwick OA, Amundson R (1996) Rapid exchange between soil carbon and atmospheric carbon dioxide driven by temperature change. Science 272:393–396. https://doi.org/10.1126/science.272.5260.393
Tucker C, Reed SC (2016) Low soil moisture during hot periods drives apparent negative temperature sensitivity of soil respiration in a dryland ecosystem: a multi-model comparison. Biogeochemistry 128:155–169. https://doi.org/10.1007/s10533-016-0200-1
Wang D, He N, Wang Q, Lü Y, Wang Q, Xu Z, Zhu J (2016) Effects of temperature and moisture on soil organic matter decomposition along elevation gradients on the Changbai Mountains, Northeast China. Pedosphere 26:399–407. https://doi.org/10.1016/S1002-0160(15)60052-2
Wu Z, Dijkstra P, Koch GW Peñuelas J, Hungate BA (2011) Responses of terrestrial ecosystems to temperature and precipitation change: a meta-analysis of experimental manipulation. Glob Change Biol 17:927–942. https://doi.org/10.1111/j.1365-2486.2010.02302.x
Wundram D, Pape R, Löffer J (2010) Alpine soil temperature variability at multiple scales. Arct Antarct Alp Res 42:117–128. https://doi.org/10.1657/1938-4246-42.1.117
Yajun X, Yonghong X, Xinsheng C, Feng L, Zhiyong H, Xu L (2016) Non-additive effects of water availability and litter quality on decomposition of litter mixtures. J Freshw Ecol 31:153–168. https://doi.org/10.1080/02705060.2015.1079559
Zhang D, Hui D, Luo Y, Zhou G (2008) Rates of litter decomposition in terrestrial ecosystems: global patterns and controlling factors. J Plant Ecol 1:85–93. https://doi.org/10.1093/jpe/rtn002
Zuur AF, Ieno EN, Walker NJ, Saveliev AA, Smith GM (2009) Mixed effects models and extensions in ecology with R. Springer, New York. https://doi.org/10.1007/978-0-387-87458-6
Acknowledgements
We thank T’ai Gladys Whittingham Forte for proof reading, and the Handling Editor and three anonymous reviewers for their comments on an earlier version of the manuscript. Fieldwork of DG was financially supported by Ente Parco Nazionale del Pollino as part of the research program “Un laboratorio di campo permanente nel Parco Nazionale del Pollino”. SC was supported by Nature State Reserve “Montagna di Torricchio” (LTER_EU_IT_033) and by Sentina Nature Reserve.
Author information
Authors and Affiliations
Corresponding author
Additional information
Responsible Editor: Feike A. Dijkstra.
Alessandro Petraglia and Cecilia Cacciatori equally contributed to this work and should be considered co-first authors
Electronic supplementary material
ESM 1
(DOCX 142 kb)
Rights and permissions
About this article

Cite this article
Petraglia, A., Cacciatori, C., Chelli, S. et al. Litter decomposition: effects of temperature driven by soil moisture and vegetation type. Plant Soil 435, 187–200 (2019). https://doi.org/10.1007/s11104-018-3889-x
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11104-018-3889-x