
Abstract
Aims
Establishing a vegetation-soil model in Mediterranean saltmarshes based on the relationships between the plant communities and the abiotic factors, considering temporary variation.
Methods
Relationships between perennial plant species abundances and plant communities were analysed by DCAs. A CCA was performed to study the relationships between floristic composition and edaphic variables. Sixteen soil variables and Pearson correlations between them were considered. Marginal and conditional effects were supported by mixed ANOVA. Statistical analyses were performed to check temporary variation.
Results
DCAs results showed eight vegetation types. CCA showed E.C. as the main gradient, with the succulent halophyte communities growing in high E.C. soils. SAR and percentage of sand were considered as secondary gradients. Finally, the highest values of the edaphic variables were observed, in general, during the cold period.
Conclusions
The main gradient of salinity, together with sodicity and texture gradients, would markedly influence the plant distribution in Mediterranean saltmarshes. Two principal plant zones were observed: succulent zone vs. non-succulent zone, with a specific edaphic distribution for each plant community and for the proposed Limonium morphotypes treatment. A plant-soil model based on these three gradients is here proposed. Our results would complement the previous knowledge about plant-soil relationships in Mediterranean saltmarshes.
Similar content being viewed by others


Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
Saltmarshes are saline environments characterised by high saline soils (> 4 mS/cm) (Chapman 1974; Álvarez-Rogel 1997; Alonso 2000). Most of these habitats appear under arid, semiarid and dry conditions, where high evaporation allows for the accumulation of gypsum, sodium and calcium carbonate in their soils (FAO-UNESCO-ISRIC 1988; Deckers et al. 1998). These habitats are widely distributed worldwide, including Arctic zones, Northern and Southeastern Europe, Mediterranean areas, Australia, South and North America, Asia and Africa, although they usually appear under Temperate and Mediterranean climates with scarce precipitations (Chapman 1974). Under these conditions, a low diversity is observed because a relatively small number of plant species can tolerate the salinity levels that occur (Alonso 2000; Lendínez 2010). Salt induces high stress in vegetation and only stress-tolerant plant species, such as halophytes, can survive (Pennings et al. 2005; Flowers and Colmer 2008; Teege et al. 2011). Halophytes are plant species, which are able to complete the life cycle in a NaCl concentration of at least 200 mM (Flowers and Colmer 2008). Throughout the saltmarsh, the halophytic vegetation is distributed in a marked zonation with a well-delimited spatial structure (Chapman 1974; Álvarez-Rogel 1997), due to various abiotic and biotic processes which lead to the distribution of the halophilous plants (Gray 1994; Cantero et al. 1998).
Plant zonation in saltmarshes has been studied in relation to the variations in edaphic gradients in different geographical areas (Davis et al. 1996; Cantero et al. 1998; Zedler et al. 1999; Piernik 2003; Álvarez-Rogel et al. 2007; González-Alcaraz et al. 2014). In addition, the interaction between soil moisture and salinity has also been recognised as the most important factor in the distribution of halophytes (Waisel 1972; Flowers and Colmer 2008). The alternation between rainy periods, during which salts are leached to the deepest soil horizons, and drought periods, when they are brought to the soil surface, have an important effect on the quantity and type of ions (Chapman 1974; Álvarez-Rogel 1997). Salinity and moisture gradients markedly influence vegetation patterns and saline habitat structure (Pielou and Routledge 1976). However, topographical variations and climatic seasonal changes in these habitats would act on soil factors, thus affecting abiotic and biotic conditions (Chapman 1974; Bertness and Ellison 1987; Álvarez-Rogel et al. 2000; Engels and Jensen 2010; Engels et al. 2011; Baumberger et al. 2012).
In this context, disentangling the role of abiotic factors in vegetation patterns has been a particularly important goal to understand the assemblage of halophytic plant communities over the decades (Montasir 1943; Adams 1963; Bertness and Ellison 1987; Pennings and Callaway 1992; Álvarez-Rogel 1997; Pennings et al. 2005; Piernik 2012). Some of the first relevant approaches to the ecology of saline environments were the comparative studies conducted by Chapman (1939, 1974), which contributed to a global and uniting view. Subsequent floristic studies in different saltmarshes have shown patterns of plant zonation among edaphic gradients (Pignatti 1952; Tadros 1953; Wolff 1968; Costa and Boira 1981; Peinado et al. 1995; Alonso 2000; Sari-Ali et al. 2012), and numerous plant-soil ecological studies have proved the existence of vegetation patterns (Neiring and Warren 1980; Snow and Vince 1984; Bertness and Ellison 1987; Ortiz et al. 1995; Álvarez-Rogel 1997; Baumberger et al. 2012; González-Alcaraz et al. 2014). Many of these studies have focused on different geographical regions with a Mediterranean climate (Ortiz et al. 1995; Peinado et al. 1995; Álvarez-Rogel 1997; González-Alcaraz et al. 2014), where saltmarshes cover extensive areas. Mediterranean saltmarshes are well represented in many regions around the southeastern Iberian Peninsula, the Greek Islands, northern Africa or western North America (Chapman 1974; Peinado et al. 1995), and appear under Thermomediterranean (Itc = 350–450) and Mesomediterranean (Itc = 210–350) thermotypes and semiarid (200–350 mm) and dry (350–600 mm) ombrotypes (Rivas-Martínez 2007). Certain authors have quantified edaphic variables, using these data to explain the observed plant zonation according to soil gradients (García et al. 1993; Cantero et al. 1998; Álvarez-Rogel et al. 2000; González-Alcaraz et al. 2014; Koull and Chehma 2016). However, most of these studies are based on few and local saltmarshes with a semiarid climate, hence a broader approach is not only feasible but necessary to identify soil features associated with halophyte assemblages (Hackney et al. 1996; Álvarez-Rogel et al. 2000). The present study has been carried out on a wider regional scale, including different halophytic communities in twenty well-distributed saltmarshes throughout the southeast of the Iberian Peninsula. Some previous studies were based on saltmarshes from the southeast of the Iberian Peninsula (Ortiz et al. 1995; Álvarez-Rogel et al. 2000, 2001; González-Alcaraz et al. 2014), although a limited sampling was considered, since they included a reduced number of saltmarshes (from one to six). Similarly, studies from other Mediterranean zones also contained a scarce number of analysed locations (Koull and Chehma 2016). Consequently, our study would represent the most comprehensive analysis to assess the plant-soil relationships in saltmarshes, since a whole regional area is here analysed in detail.
Not only the sampling size would be a relevant factor for this type of studies, but also the selected statistical analyses, since the obtained results would be affected as well. Unconstrained ordination methods, such as Principal Components Analysis (PCA) or Detrended Correspondence Analysis (DCA), are commonly used to describe vegetation patterns or main gradients in community structure. However, these analyses do not include environmental factors in their algorithm and are more appropriate to explain the variability in species composition rather than its relationship with environmental variables (Lepš and Šmilauer 2014), although they can be integrated as supplementary variables or even they can be included in the role of plant species (ter Braak and Šmilauer 1999). Conversely, constrained ordination methods, such as Canonical Correspondence Analysis (CCA), would consider environmental variables in their algorithms and they would directly relate vegetal composition to environmental factors (Lepš and Šmilauer 2014). Besides, constrained ordination methods are able to test the variability on vegetation composition explained by the environmental variables (Lepš and Šmilauer 2014). In our study, both types of ordination analyses have been considered since the two approaches are complementary to assess plant-soil relationships (Lepš and Šmilauer 2014), although in earlier studies usually only either constrained or unconstrained ordination analyses are used (Cantero et al. 1998; Álvarez-Rogel et al. 2000, 2001; González-Alcaraz et al. 2014; Koull and Chehma 2016). In addition, analyses of marginal and conditional effects and Pearson correlations between edaphic variables are also appropriate, as they provide relevant information to support any plant-soil model proposed. For the western Mediterranean area, neither González-Alcaraz et al. (2014) nor Álvarez-Rogel et al. (2000, 2001) analysed the correlations between the studied edaphic variables.
This study represents a broader approach, since twenty saltmarshes distributed throughout the southeast of the Iberian Peninsula were taken into account in our research, using constrained ordination methods for their analyses. The aims of the present study were: (1) to identify the halophytic plant communities in saline environments based on ordination methods; (2) to contribute to the knowledge of the relationships between edaphic factors and vegetation in saline habitats of the studied zone; (3) to analyse the temporal changes of soil moisture and salinity in saline habitats of the studied zone; and (4) to establish a vegetation-soil model to relate the spatial plants distribution patterns in these peculiar environments.
Materials and methods
Study area
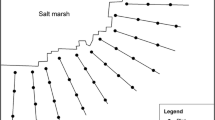
The study area was located at the southeast of the Iberian Peninsula, under Thermomediterranean and Mesomediterranean thermotypes, and semiarid, arid and dry ombrotypes (Rivas-Martínez 2007). Study sites were selected after several visits to different saltmarshes spread throughout an extension of approximately 13,000 km2. Twenty saltmarshes were selected, completely covering the southeast of the Iberian Peninsula (Fig. 1 and Supplementary material Appendix A), according to the following criteria: (1) they should be well-separated and independent from each other, (2) they should have a representative halophytic vegetation and not be excessively degraded, and (3) they should have a zonation of vegetation with at least two different plant formation structures (identified by the dominant plant species), thus indicating the existence of ecological gradients.

Map of the study area, showing the position of the twenty studied Mediterranean saltmarshes
Sampling design
Sampling was performed on a seasonal basis, i.e. every three months for one year (July 2013–April 2014). Samples were grouped in two temperature periods, based on meteorological data collected during the sampling year. Temperature periods were considered instead of precipitation periods due to the variations in precipitation patterns among the studied zones. The warm period lasted from mid-April to mid-October and the cold period lasted from mid-October to mid-April. Daily precipitation and temperature data for each site from the nearest meteorological station were used to calculate accumulated precipitation and minimum, mean and maximum temperatures (Supplementary material Appendix B). Climatic variables were provided by AEMET (M° de Agricultura, Alimentación y Medio Ambiente, Spain).
In each sampling station, different plant formation structures were recognised on the basis of the dominant plant species observed in the field. A 100 m2 plot was established randomly in each studied plant formation structure (63 plots in total), and georeferenced using GPS Garmin© Oregon 300. The number of plots in each saltmarsh ranged from two to five according to the presence of different plant formation structures (Supplementary material Appendix A). Floristic composition and plant cover were recorded in each plot using the Braun-Blanquet scale with seven levels (r, +, 1, 2, 3, 4, 5), according to the sigmatist methodology (Braun-Blanquet 1979). Plant species were identified according to regional keys for plant identification (Castroviejo 1986–2015; Mateo and Crespo 2009; Blanca et al. 2011) and specialised research papers (Piirainen et al. 2017; Moreno et al. 2016, 2018). A total of 252 plant inventories (63 plots × 4 times) were obtained and a total of sixty-two plant species were recorded in the twenty studied Mediterranean saltmarshes. Plant inventories were averaged for each studied plot because of the scarce vegetation variation across the seasonal periods (Supplementary material Appendix C). Subsurface soil samples were collected at 20 cm depth to avoid the superficial salt crust. Three soil subsamples were collected randomly in each plot and were mixed in order to obtain a representative soil sample. A total of 252 soil samples (63 plots × 4 times) were obtained in the twenty Mediterranean saltmarshes studied, which were averaged for each studied plot (Supplementary material Appendix D).
Soil analyses
Soil moisture was estimated immediately after collection using the water retention method of 12 h in an oven at 110 °C (Burt 2004). The remaining sample was air-dried, crushed and 2-mm-sieved to remove coarse fragments before laboratory analyses (Burt 2004). Saturation extracts were obtained from saturated pastes through vacuum filtering. Electrical conductivity (E.C.) and pH were measured in saturation extracts using a conductivity meter Crison© CM 35+ and pH meter Crison© 25, respectively. Saturated pastes were dried for 12 h at 110 °C and the saturation percentage was calculated to estimate available water capacity (Burt 2004), hereafter named Plant Available Water Capacity (PAWC). For the calculation of soil moisture and PAWC, no correction for the structural water of gypsum was considered. In addition, specific concentrations of sodium (Na+), potassium (K+), calcium (Ca2+), magnesium (Mg2+) and sulphur (S) [used as an indicator of sulphate (SO42−)] in the saturation extracts were measured through Inductively Coupled Plasma Atomic Emission Spectroscopy (ICP-AES) (Perkin Elmer 7300 DV). The wavelengths used were 589 nm for Na+, 766 nm for K+, 317 nm for Ca2+, 279 nm for Mg2+ and 180.7 and 182.0 nm for S. Cation concentrations were used to calculate the Ca2+/Mg2+, Ca2+/Na+, and K+/Na+ ratios, and the Sodium Adsorption Ratio (SAR) (Burt 2004). Chloride (Cl−) concentration in the saturation extracts was measured using argentometry with silver nitrate (AgNO3) (Harris 2003). Finally, soil texture was determined using a Bouyoucos densitometer (Juárez et al. 2004), and soil colour was recorded for wet and dry samples in each study site according to the Munsell colour chart (Munsell® Corporation 1994). Soil texture and colour of the twenty studied sites were included in Supplementary material Appendix E.
Statistical analyses
A first Detrended Correspondence Analysis (DCA) was performed to observe the relationships between perennial plant species abundance and plant communities using CANOCO v. 5 (Microcomputer Power, Ithaca, NY, US). This analysis was carried out with the complete data set of the sixty-three plant inventories taken at the twenty saltmarshes. Besides, a second DCA was performed and focused on the samples dominated by Limonium sp. pl. or Lygeum spartum L. to discern between these plant communities. In this second analysis, different perennial Limonium species were joined according to different morphotypes based on their traits: (1) numerous sterile branches and absent leaves at the anthesis [L. caesium (Girard) Kuntze and L. insigne Kuntze] (LimA); (2) some sterile branches and withered leaves present at the anthesis [L. furfuraceum (Lag.) Kuntze, L. parvibracteatum Pignatti and L. supinum (Girard) Pignatti] (LimB); and (3) absence of sterile branches and green leaves, mostly coriaceous, at the anthesis [L. admirabile Terrones et al., L. angustebracteatum Erben, L. cossonianum Kuntze, L. delicatulum (Girard) Kuntze and L. tobarrense J. Moreno et al.] (LimC). In both analyses, plant species covers were converted to percentages according to the approximation of Braun-Blanquet (1946). All obtained percentages were log-transformed [log (y + 1)] and rare plant species were downweighted using the options of the software (Lepš and Šmilauer 2014). On the basis of the results of the DCA, plant communities were defined and assigned to each plot.
Pearson correlations between pairs of edaphic variables were carried out to establish relationships between them using the ‘rcorr’ function of the ‘Hmisc’ package with R software v. 3.3.0 (R Core Team 2016). Correlations were performed with the complete set of soil data, including seasonal data. To study the relationships between floristic composition and the edaphic variables, a Canonical Correspondence Analysis (CCA) was performed using CANOCO v. 5 (Microcomputer Power, Ithaca, NY, US). This analysis was carried out with the complete data set of the sixty-three plant inventories taken at the twenty saltmarshes and the sixteen soil variables averaged annually. Plant species covers were converted to percentages according to the approximation by Braun-Blanquet (1946). The percentages were log-transformed [log (y + 1)] and rare plant species were downweighted using the options of the software (Lepš and Šmilauer 2014). Sixteen edaphic variables were included in the environmental matrix: soil moisture; E.C.; pH; PAWC; Na+, K+, Ca2+, Mg2+, Cl− and SO42− concentrations; Ca2+/Mg2+, Ca2+/Na+ and K+/Na+ ratios; SAR and percentages of sand and clay. Ca2+/Mg2+ and Ca2+/Na+ ratios, SAR, pH, PAWC, percentage of sand and Mg2+ were log-transformed to fulfil the normality requirements, while square root transformation was used with Cl−, K+, Na+ and SO42− concentrations, E.C., soil moisture, K+/Na+ ratio and percentage of clay. Ca2+ was not transformed. The amount of variability in the floristic composition explained by the CCA was calculated for marginal and conditional effects. Marginal effects denoted the variability explained by the given set of environmental variables without considering other environmental factors, whereas conditional effects denoted the variability explained by the given environmental set after removing the confounding effect of one or more other environmental variables (covariables) (Lepš and Šmilauer 2014). Conditional effects were performed by a stepwise selection. Marginal and conditional effects were tested for significance using Monte Carlo permutation tests with 9999 permutations. In addition, a similar CCA, but using E.C. as covariable, was performed to verify the effect of the studied edaphic variables on the floristic composition of plant communities, avoiding the effect of E.C.
Mixed analyses of variance (ANOVAs) were performed to detect significant differences (P ≤ 0.05) for the edaphic variables. Three different predictors were considered: (i) type of halophytic plant community, which was treated as an independent-measure factor, (ii) period, which was treated as a repeated-measure factor, and (iii) their interaction. Saltmarshes were included as a nested factor. Models were created with the ‘lme’ function and tested with the ‘anova’ function (‘nlme’ package); and type II sums of squares were used in order to avoid problems with the unequal sample sizes (Tabachnick and Fidell 2007). These analyses were conducted using R software v. 3.3.0 (R Core Team 2016). To check significant differences between periods in each plant community type, paired t-test analyses were performed using the ‘t.test’ function. ANOVAs were performed to check significant differences between plant community types in every period using the ‘lm’ and ‘anova’ functions. For significant differences in these analyses, Tukey tests were performed with the ‘glht’ function (‘multcomp’ package). Soil texture variables (percentages of sand and clay) and PAWC were considered only in comparisons between different halophytic plant communities and not in comparisons comprising time because they are constant during the year. These analyses were conducted using R software v. 3.3.0 (R Core Team 2016). Graphs were plotted with the ‘GrapheR’ package using R software v. 3.3.0 (R Core Team 2016) to observe significant differences.
Results
Plant communities in saltmarshes
Based on the DCA ordination diagram (Fig. 2a), some plant groups were observed and each studied plot was assigned to the most similar vegetation group. Several polygons were drawn to identify these vegetation groups on the basis of the dominant plant species in the samples. The first two axes of the analysis explained 21.83% of total variation (12.87% and 8.96% of the variance for first and second axes, respectively). According to this analysis, seven groups could be identified (Fig. 2a). Group 1 included plots dominated by Halocnemum strobilaceum (Pall.) M. Bieb. (No. 54 and 56). Group 2 was formed by those plots dominated by Arthrocaulon macrostachyum (Moric.) Piirainen & G. Kadereit (No. 1, 7, 20, 21, 25, 31, 35, 36, 41, 45, 48 and 52). Group 3 corresponded to those plots with Salicornia fruticosa (L.) L. as dominant species (No. 6, 13, 14, 17, 19, 24, 28, 37, 40, 44, 49, 51 and 60), although A. macrostachyum and Suaeda vera Forssk. ex J.F. Gmel. might appear in certain plots. These first three groups were dominated by different perennial succulent halophytes, forming generally monospecific plant formations. Group 4 was formed by plots dominated by Limonium caesium (No. 10, 29, 57 and 58), although the halonitrophilous shrub S. vera also appeared. Group 5 included plots dominated by Tamarix species (No. 9, 18, 23, 30, 59 and 63). Group 6 corresponded to those plots characterised by having Lygeum spartum as dominant species (No. 3, 5, 11, 12, 16, 27, 32, 33, 43, 47 and 55), which might be accompanied by certain Limonium species. Finally, Group 7 included the plots with a clear dominance of different Limonium species such as L. angustebracteatum, L. cossonianum, L. delicatulum, L. insigne and L. parvibracteatum (No. 2, 4, 8, 15, 22, 26, 34, 38, 39, 42, 46, 50, 53, 61 and 62). Conversely to Groups 1, 2 and 3, these latter groups were more floristically diverse, which could be easily observed from the major dispersion of the plots in the diagram (Fig. 2a).

a Detrented Correspondence Analysis (DCA) carried out with the data of the sixty-three plant inventories taken at the twenty studied Mediterranean saltmarshes. The ordination diagram represents only the fifteen plant species best predicted by the explanatory variables. Species abbreviations: ArtMac, Arthrocaulon macrostachyum; HalPor, Halimione portulacoides; HalStr, Halocnemum strobilaceum; LimAng, Limonium angustebracteatum; LimCae, Limonium caesium; LimCos, Limonium cossonianum; LimDel, Limonium delicatulum; LimIns, Limonium insigne; LimPar, Limonium parvibracteatum; LimSup; Limonium supinum; LygSpa, Lygeum spartum; SalFru, Salicornia fruticosa; SuaVer, Suaeda vera; TamBov, Tamarix boveana; TamGal, Tamarix gallica. b Detrented Correspondence Analysis (DCA) carried out with the samples of Limonium group at the twenty studied Mediterranean saltmarshes. The ordination diagram represents only the five plant species best predicted by the explanatory variables. Species abbreviations: Lim A, Limonium caesium and L. insigne; Lim B, Limonium supinum type; Lim C, Limonium delicatulum group; LygSpa, Lygeum spartum; SuaVer, Suaeda vera
The second DCA, focused on Limonium sp. pl. and L. spartum communities, showed the identification of four groups on the basis of their different traits (Fig. 2b). The first two axes of the analysis explained 32.69% of the total variation. First axis explained 21.24% of variance and the second one explained 10.45% of variance. Those plots dominated by L. spartum appeared clearly isolated from the remaining groups, and morphologically close Limonium species were grouped together. The Limonium plots were differentiated in three different groups instead of the two initial groups (Fig. 2a, b). The first group, Group 7A, was formed by plots dominated by Limonium morphotype A (No. 10, 29, 39, 57 and 58), with the species L. caesium and L. insigne. In the first DCA, these plots mostly belonged to Group 4 (dominated by L. caesium), but plot No. 39, characterised by the dominance of L. insigne, was initially added to Group 7. Group 7B was formed by those plots dominated by species of Limonium morphotype B (No. 8, 15, 34 and 53), such as L. supinum and L. parvibracteatum. Finally, Group 7C included those close plots dominated by Limonium morphotype C (No. 2, 4, 22, 26, 38, 42, 46, 50, 61 and 62), with the species L. delicatulum, L. cossonianum or L. angustebracteatum. In the first DCA (Fig. 2a), both groups of Limonium formed Group 7, though those plots dominated by Limonium morphotype B appeared rather close to Group 6 (see Fig. 2a, plots separated by a dashed line).
According to DCAs results, eight vegetation types were finally considered from then on (Fig. 2a, b): Halocnemum strobilaceum communities (Group 1); Arthrocaulon macrostachyum communities (Group 2); Salicornia fruticosa communities (Group 3); Limonium A zones (Group 7A, including Group 4); Limonium zones B (Group 7B); Limonium zones C (Group 7C); Lygeum spartum steppes (Group 6); and saltcedar woodlands (Group 5).
Relationships between halophytic plant communities and edaphic properties
Regarding the results of Pearson correlations, many edaphic variables showed strong positive correlations between them (Supplementary material Appendix F). E.C. was highly correlated with Na+, K+, Ca2+, Mg2+, Cl− and SO42− concentrations and SAR (r ≥ 0.50), and many of these variables were also highly correlated between them (r ≥ 0.52), except for Mg2+ and SO42− concentrations with Ca2+, Cl− concentrations and SAR; and K+ concentration with Ca2+ concentration. Ca2+/Na+ and K+/Na+ ratios were correlated between them and with E.C., Na+ concentration and SAR (|r| ≥ 0.50). Besides, soil moisture was strongly correlated with PAWC (r = 0.71), and Ca2+ concentration was negatively correlated with pH (r = −0.55).
The CCA showed the relationships between the halophytic plant communities identified here and edaphic variables (Fig. 3). The first two axes of the analysis explained 25.07% of total variation and 66.70% of fitted variance. The first axis explained 9.54% of the variance (25.38% of the fitted variation), and the second one explained 15.53% of the variance (41.32% of the fitted variation). The studied edaphic variables had a high significant effect upon the halophytic plant community composition (P ≤ 0.001). Marginal effects showed that all edaphic variables were significant to explain the variation in the vegetation patterns (P ≤ 0.05), but for Ca2+/Mg2+ ratio and percentage of clay (Table 1). The largest part of the variance was explained by E.C., Na+, Cl− and SAR (Table 1). Similar results could be observed in the ANOVA, all edaphic variables had significant differences between vegetation types (P ≤ 0.05), except the percentages of clay and sand (Table 2). However, conditional effects were significant for only seven variables to explain the variation in vegetation patterns (P ≤ 0.05): E.C. (7.7%), SAR (3.9%), the percentage of sand (3.2%), K+ (2.4%), Mg2+ (2.4%) and pH (2.4%, Table 1). Nonetheless, E.C., SAR and the percentage of sand explained most of the variability. Besides, no difference between marginal and conditional effects for the percentage of sand showed that the variance explained by this variable was not explained by E.C. nor SAR. Conversely, the difference of values for SAR indicated that part of its variance was explained by E.C., and hence E.C. can mask the effect of SAR (Table 1). These results are congruent with Pearson correlations, as E.C. is strongly correlated with all ion concentrations and SAR (see above). These relationships between edaphic variables were also revealed in the CCA ordination diagram (Fig. 3). The CCA results showed the importance of E.C. as the main variable to explain the vegetation patterns in saline habitats. Succulent halophyte communities would appear in areas with high E.C., whereas Limonium A zones and saltcedar woodlands would appear in the least saline areas, leaving the intermediate zones for Limonium B and C zones and L. spartum steppes (Fig. 3). Related to these three latter groups, Limonium C communities would appear in zones with higher E.C. than both Limonium B communities and L. spartum steppes, and Limonium B zones and L. spartum steppes would appear together in the diagram (Fig. 3). However, no significant differences could be observed between these three vegetation types for E.C. in ANOVA (Table 3). Therefore, the E.C. gradient was particularly relevant to separate halophytic communities in saltmarshes (Table 3 and Fig. 3). Accordingly, cation concentrations and SAR were, in general, higher in succulent halophyte communities than in the remaining vegetation types, as well as soil moisture (Table 3). These variables tended to have higher values in H. strobilaceum communities than in S. fruticosa communities and A. macrostachyum communities, but this pattern changed for Ca2+ concentration and soil moisture, for which no strong differences were observed (Table 3). However, these succulent plant communities, in general, did not appear clearly differentiated between them in the CCA diagram (Fig. 3). Nevertheless, some significant differences could be observed for Mg2+, K+ and SO42− concentrations and Ca2+/Mg2+ ratio (Table 3), but only Mg2+ and K+ concentrations were significant in conditional effect analyses (Table 1). Therefore, SO42− concentration and Ca2+/Mg2+ ratio could not be directly responsible for the changes in succulent vegetation. Moreover, it should also be pointed out that soil texture showed wide variability in the different studied vegetation types, but no significant differences were observed in the ANOVA (Tables 2 and 3). However, the highest percentage of sand was observed in the H. strobilaceum and Limonium A communities (Table 3), and both plant communities were related to high percentages of sand (Fig. 3).

Canonical Correspondence Analysis (CCA) of sixty-three samples of halophytic communities from studied Mediterranean saltmarshes showing correlations between samples and edaphic variables. Arrows indicate the edaphic variables and their directions and length show their relationships to the ordination axes. Edaphic variables abbreviations: E.C., electrical conductivity; Moisture, soil moisture; PAWC, plant available water capacity; SAR, sodium adsorption ratio. Plant community abbreviations: A. macrostachyum, Arthrocaulon macrostachyum; H. strobilaceum, Halocnemum strobilaceum; L. spartum, Lygeum spartum; S. fruticosa, Salicornia fruticosa
Since E.C. was correlated with many other variables and it might be influencing part of the variability of other significant variables in conditional effects (i.e. part of the SAR effect is explained by E.C.), a second CCA was performed including E.C. as covariable (Supplementary material Fig. A1). According to this analysis, H. strobilaceum and Limonium A community samples appeared in zones with low SAR and high percentage of sand, whereas Tamarix community samples were located in the high SAR and the low percentages of sand (Supplementary material Fig. A1). However, soil texture factors were not significantly different for any vegetation types in the ANOVA (Table 2). In the case of SAR, this second CCA only shows the part of the variability that is not explained by E.C., and hence, the results of this analysis cannot be directly compared to the ANOVA because they refer to different variables.
Temporal differences in soil conditions and between halophytic plant communities
According to the mixed ANOVA, no interaction between period and vegetation type was significant for any soil variable (P > 0.05, Table 2). Temporal changes have been clearly identified in soil conditions, with the higher values of E.C., ionic concentrations, SAR and soil moisture during the cold period (Table 4). This seasonal pattern was well supported by significant variations between both periods for certain edaphic variables such as E.C., Na+, Mg2+, K+ and soil moisture (Tables 2 and 4). The Ca2+/Mg2+ ratio showed the highest values during the warm period; and the remaining soil variables showed similar data between both periods (Table 4).
Some temporal differences could be observed between soil conditions and different halophytic vegetation types. In general, for each studied plant community, the edaphic variables displayed higher values in the cold period than in the warm period (Fig. 4). These seasonal changes have been significantly different from certain soil variables for some vegetation types, so E.C. and Na+, Mg2+ and K+ concentrations were significantly different for A. macrostachyum and L. spartum communities (Fig. 4 and Supplementary material Appendix H). Besides, E.C. in H. strobilaceum communities and Na+ concentration in S. fruticosa communities also showed significant differences (Fig. 4 and Supplementary material Appendix H). In the case of soil moisture, this edaphic feature was only significantly different for both S. fruticosa communities and L. spartum steppes (Fig. 4 and Supplementary material Appendix H). Finally, Limonium zones and saltcedar woodlands did not have, in general, statistical support for any of the mentioned edaphic variables (Fig. 4 and Supplementary material Appendix H).

Variations in electrical conductivity (E.C.), Na+, K+ and Mg2+ concentrations, and soil moisture in the different halophytic communities depending on the period. Shared letters indicate no difference between halophytic plant communities for each period (Significance test P ≤ 0.05). Asterisks show significant differences between periods within the same vegetation type (Significance test P ≤ 0.05). Vegetation types abbreviations: Haloc, Halocnemum strobilaceum communities; Arth, Arthrocaulon macrostachyum communities; Salic, Salicornia fruticosa communities; LimC, Limonium groups C; LimB, Limonium group B; Lyg, Lygeum spartum steppes; LimA, Limonium group A; Tam, saltcedar woodlands
Some relevant differences for soil variables were observed between vegetal communities within each period (Fig. 4). In general, E.C. and Na+ were significantly different between succulent communities and non-succulent communities in both periods, showing succulent communities the highest average values (Fig. 4). Besides, soil moisture showed significant differences in the cold period, being higher in A. macrostachyum and S. fruticosa communities and lower in Limonium C zones and L. spartum steppes (Fig. 4); these variables exhibited similar significant differences in the warm period (Fig. 4).
Discussion
Main soil gradient in halophytic zonation
The obtained soil analyses would indicate the existence of a salinity gradient, defined mainly by E.C., but also correlated with Na+, K+, Mg2+, Ca2+, Cl− and SO42− concentrations, K+/Na+ and Ca2+/Na+ ratios and SAR, as the main edaphic gradient to lead vegetation distribution in saltmarshes. Our results agreed with other recent reports (González-Alcaraz et al. 2014; Koull and Chehma 2016), indicating that salinity would have a notable impact on plant distribution in saline Mediterranean habitats. In general, salinity, based on specifically the presence of high Na+, Mg2+, Cl− and SO42− concentrations, has a well-known negative effect on plant growth, involving osmotic effects, toxicity of certain ions, especially Na+ and Cl−, and nutrient deficiencies, particularly for K+ and Ca2+ (Bernstein 1975; Lambers et al. 1998). However, halophytes show a combination of several different morphological and physiological traits that allow them to support the negative effects of salinity (Flowers and Colmer 2008). Depending on the salt adaptation trait of the different halophyte species, several tolerances to salinity would be observed. According to our results, and similarly to Ortiz et al. (1995), perennial succulent halophytes, such as A. macrostachyum and S. fruticosa communities, would dominate in soils with high salinity. Arthrocaulon macrostachyum communities would appear in soils characterised by higher E.C. than in the case of S. fruticosa communities, even showing significant differences in Na+, Cl−, Ca2+ and K+ concentrations and pH probably causing that both succulent shrub communities are able to form well-separated vegetation zones within saline environments (Álvarez-Rogel 1997; Alonso 2000; González-Alcaraz et al. 2014). Although both plants show different germination syndromes (Muñoz-Rodríguez et al. 2017), the germination syndrome of both succulent species cannot explain by itself the observed spatial distribution. Arthrocaulon macrostachyum presents high levels of germination at low-medium salinities (up to 0.6 M NaCl) and its germination is notably reduced after high salinity exposure (Vicente et al. 2007; Muñoz-Rodríguez et al. 2017), though Pujol et al. (2000) exposed high recovery values of seed germination after osmotic stress. Conversely, the seeds of S. fruticosa easily germinate in a wider range of salinities, even at high salinity (up to 1 M NaCl) (Redondo et al. 2004; Muñoz-Rodríguez et al. 2017). Another feasible explanation would be the one proposed by González-Alcaraz et al. (2014), which is that A. macrostachyum appears in zones with high seasonal variation, and S. fruticosa appears in more stable zones. Our results are congruent with this model, since A. macrostachyum communities showed a higher seasonal variation in E.C. than S. fruticosa communities.
The highest salinity values have been obtained in H. strobilaceum communities in comparison to the remaining succulent plant communities, although these data must be taken with some caution due to the low number of analysed samples. Nevertheless, our results were similar to Álvarez-Rogel et al. (2000), where H. strobilaceum was principally related to the highest SAR values and lowest Mg2+ content in relation to Ca2+, considering that these variables are correlated with E.C. Besides, Koull and Chehma (2016) observed a high relation between H. strobilaceum communities and soil salinity and moisture; and Chigani et al. (2010) proved that salinity, K+ concentration, CaSO4 and clays have positive effects on H. strobilaceum formations. In addition, some phytosociological studies described the soils occupied by H. strobilaceum as more saline soils than A. macrostachyum community soils (Pignatti 1952; Wolff 1968), although certain previous studies were not able to find any differences between H. strobilaceum and A. macrostachyum communities (Kassas and Zahran 1967; Alcaraz et al. 1987).
Limonium zones and L. spartum steppes would appear in soils characterised by lower salinity, showing an intermediate position between succulent and Tamarix communities along the saline gradient and sharing ecological space on several occasions. These results are similar to many previous studies in which Limonium communities also occurred at an intermediate position along the saline gradient (Álvarez-Rogel et al. 2001; Baumberger et al. 2012; González-Alcaraz et al. 2014), although Ortiz et al. (1995) reported that Limonium bands occupied the same ecological position as the succulent species.
Secondary soil gradients in plant distribution
Our results suggest that the salinity gradient would be followed by two secondary soil gradients: a sodicity gradient and a texture gradient. The sodicity gradient was led by SAR, which is strongly correlated with E.C. and Na+. Consequently, SAR values did not only indicate sodicity but also salinity, hence these values can only be directly compared in similar values of E.C. High sodicity has been reported to have negative effects on plant growth, since it can involve nutrient deficiencies, especially for Ca2+ and Mg2+, and cause a poor soil structure (Bernstein 1975). This gradient would clearly separate Limonium A group and saltcedar woodlands from the remaining vegetation formations within the non-succulent zone, and it would also distinguish H. strobilaceum communities from the remaining perennial succulent communities (Fig. 5). Álvarez-Rogel et al. (2000) exposed Tamarix communities were related to soils with lower SAR, although, in this mentioned study, SAR is probably representing salinity since in saltmarshes salinity and sodicity are highly correlated. In addition, similar results have also been obtained for the separation of H. strobilaceum and A. macrostachyum by Álvarez-Rogel et al. (2000) and Abdul-Halim and Ismail (1990).

Plant-soil model in Mediterranean saltmarshes
The second gradient would correspond to soil texture, mainly led by the percentage of sand. Álvarez-Rogel et al. (2001) suggested soil texture as a relevant edaphic factor, which might influence the plant zonation of saltmarshes, although references to soil texture differences were initially reported in certain western Mediterranean vegetation studies (Tadros 1953; Wolff 1968). However, this factor has not been considered in recent studies about plant assemblage in saline environments (González-Alcaraz et al. 2014). In this framework, our study presents for the first time a quantitative analysis about soil texture factors and their notable influence on saltmarsh structure. Our CCA data revealed the relevance of percentage of sand in plant distribution in saltmarshes, although ANOVA was not significant. Despite the low number of samples analysed, the highest values of percentage of sand, were found in H. strobilaceum communities, which was a plant community in the zones also characterised by high E.C. These results coincide with other ecological studies, where H. strobilaceum communities appeared in sandy soils with high E.C. (El-Ghani et al. 2014; El-Amier 2016). Both studies were located in Egypt, therefore our results would be similar to other zones of the Mediterranean Basin. The percentage of sand would also clearly separate two different halophytic communities: Limonium A zones and saltcedar woodlands, both plant communities over the lowest salinity. The soils of Tamarix communities were characterised by a low percentage of sand and a high percentage of clay, whereas Limonium A appeared in soils with opposite values. These results are congruent with those found in the sodicity gradient, since both factors influence soil structure. Halocnemum strobilaceum and Limonium A communities would appear in zones with coarse texture soils and with low SAR (considering the salinity), the last of which promotes a well-developed structure in soil. Both factors favour a high permeability to water and air (Bernstein 1975). In the case of saltcedar woodlands, the opposite patterns are observed.
Limonium morphotypes and edaphic relationships
The genus Limonium has been really bound to salinity, but different ecological ranges would be observed depending on the Limonium species. Some of these species are able to tolerate a wide gradient of salinity, although certain Limonium species could appear in environments other than saltmarshes (Erben 1993). In Mediterranean saltmarshes, this genus includes a great diversity of species and they would be located in low saline soils (Alonso 2000; Álvarez-Rogel et al. 2000; González-Alcaraz et al. 2014). The treatment of Limonium species as morphotypes in these ecological studies is a novel methodology and, it could be really useful to focus on complex taxonomical plant groups. There are many endemic Limonium species and some of these species are led by geographical differences, but have similar soil characteristics. Each Limonium morphotype would appear in soils with specific edaphic features, thus showing a close morphotype-soil relationship. In this way, this methodology would avoid problems related to their precise taxonomical identification in studies about their role on specific ecosystems, such as saltmarshes.
In our model, the Limonium morphotype A, whose typical species would be L. caesium or L. insigne, is characterised by the presence of numerous sterile branches and the absence of leaves at the anthesis, and would prefer sandy soils with a low E.C. and SAR. These species have been observed forming a plant community different and separated from the remaining Limonium species in the saltmarshes. Nonetheless, individuals of this morphotype would scarcely appear under the canopy of Tamarix communities, which would be explained by the similar range of salinity tolerance of species of this morphotype.
The Limonium species characterised by absence of sterile branches and the presence of green leaves, mostly coriaceous, at the anthesis such as L. cossonianum, L. angustebracteatum or L. delicatulum, defined as Limonium morphotype C, would typically appear in soils with higher E.C., Na+, K+, Cl−, and SAR, appearing in an intermediate ecological position between succulent plant communities and L. spartum steppes.
Finally, the Limonium morphotype B, which includes species with withered leaves present at the anthesis such as L. supinum or L. parvibracteatum, has been generally observed close to or even in L. spartum steppes, sharing the same ecological space and edaphic features. Halophilous L. spartum steppes typically contains some Limonium species (Alonso 2000) so it is not surprising both plant species share similar requirements to grow. It might also be explained by the availability of soil between the clusters of L. spartum individuals and the high colonisation of this Limonium morphotype, or even by the ability to do photosynthesis efficiently using their sterile branches in the case of dense L. spartum formations.
Temporal changes in halophytic plant communities
Both E.C. and ionic concentrations have revealed, in general, higher values in the cold period than in the warm period, which might be related to the annual rainfalls. Seasonal differences have been observed previously in saline environments (Wolff 1968; Neiring and Warren 1980; Álvarez-Rogel et al. 2000; Koull and Chehma 2016), but perennial species were not generally affected by them (García et al. 1993). Wolff (1968) reported higher salinity in summer than in winter, and Álvarez-Rogel et al. (2000) showed similar patterns in a temporal wet-dry model, in which the wet period shown higher rainfall accumulation. Our warm period (April–October) recorded the highest annual precipitations, with an unusually rainy month of August. In this way, the total amount of rainfall would determine the salinity factors in the saltmarshes. Hence, E.C., ionic concentrations and ratios, have normally showed the lowest values during the rainy season. This decreased pattern would be markedly related to the lixiviates of salts (Álvarez-Rogel et al. 2000). Despite the fact that temporal variations would not be similar in every saltmarsh zones, it might also be due to the influence of soil moisture on the dynamic of cation adsorption by the complex exchange, the different levels of salts solubility, and the different types of clay mineral (Álvarez-Rogel et al. 2000). In addition, other factors that might be considered would be the specific behaviour of some salts. For instance, certain salts, such as NaCl, have a high solubility and are more mobile in the soil profile, and its seasonal changes would affect the solubility of other salts such as CaCl2, MgCl2 and Na2SO4 (Álvarez-Rogel et al. 2000). Our data indicated the general presence of the lowest values of Na+, Mg2+, SO42− and Cl− in the warm period, related to the fact that more precipitations were recorded, being the differences even significant for certain halophytic vegetation, such as A. macrostachyum, S. fruticosa and L. spartum communities. Therefore, it would be feasible to consider that the soil texture together with the dynamic of cation adsorption by the complex exchange would be both involved in temporal changes, and the variations in soil features of saline environments, in general, would be a function of precipitation/dissolution of salts (Koull and Chehma 2016).
Conversely to previous studies (Álvarez-Rogel et al. 2000; González-Alcaraz et al. 2014), high soil moisture has still been observed in the period with low precipitations. It might be explained by the temperature regime, since the lower temperature during the cold period would slow down evaporation, and soil moisture would maintain higher values in the cold period. In addition, water table level would also be related to the changes in soil moisture, though no quantitative information is available from any of the studied Mediterranean ecosystems. All the studied halophytic communities have exhibited changes in soil moisture, showing their highest values in the cold period, but the difference between both periods was rather minimal. However, S. fruticosa and L. spartum zones were the only plant communities with significant differences. Both vegetal communities were characterised by unique and similar proportions of sand and clay, showing approximately twice as much sand as clay (about 33% and 16%, respectively). Therefore, the different soil texture features would be also influencing edaphic moisture, as Álvarez-Rogel et al. (2000) and González-Alcaraz et al. (2014) suggested. Nevertheless, it should be pointed out that although our study includes variables related to soil texture, more detailed analyses would be necessary to support and complete these findings.
Plant-soil model in Mediterranean saltmarshes
According to the model proposed here (Fig. 5), Mediterranean saltmarshes can be divided in two areas based on the salinity gradient, led by E.C., although additional soil gradients would also act in community distribution. Firstly, the salinity gradient would separate two well-distinguished areas: (1) a succulent zone, formed by A. macrostachyum, H. strobilaceum and S. fruticosa communities with the highest E.C. values, and (2) a non-succulent zone, including Limonium zones, L. spartum steppes and saltcedar woodlands, characterised by the intermediate and lowest E.C. values (Fig. 5). Some preceding studies from small areas of the southeastern Iberian Peninsula showed, in general, that two main edaphic gradients led by soil salinity and soil moisture would be involved in halophytic zonation (Ortiz et al. 1995; Álvarez-Rogel et al. 2000; González-Alcaraz et al. 2014). However, Álvarez-Rogel et al. (2000) provided a classification based only on soil moisture characteristics, and two saline zones were distinguished: (1) wet saltmarsh, where perennial succulent plants, hydrophytes and Tamarix communities were included, and (2) dry saltmarsh, where L. spartum steppes, Limonium and halonitrophilous shrub communities were located. Recently, González-Alcaraz et al. (2014) suggested a classification based not only on soil moisture but also on soil salinity: (1) the most saline and the wettest soils, where succulent halophytes and P. australis (Cav.) Trin. ex Stende communities developed, and (2) the driest and least saline soils, where L. spartum steppes, Limonium and halonitrophilous shrub communities appeared.
Soil moisture has not been identified as a relevant gradient in our analysis, being highly correlated with salinity and having a low variance explained by itself. Other previous studies included Phragmites australis and Juncus sp. pl. communities, which have been typically observed reaching the maximum cover in the most humid soils in saltmarshes because they can resist permanent flood (Alonso 2000; Álvarez-Rogel et al. 2000; González-Alcaraz et al. 2014; Koull and Chehma 2016). Nevertheless, we have considered that it was not appropriated to include hydrophyte communities in this study since its inclusion would enhance the effect of soil moisture and reduce that of E.C., allowing us to avoid the overestimation of the effect of soil moisture in plant-soil relationships of the non-permanently flooded communities. As a result, our data emphasise the importance of E.C. in the zonation of halophytes, being the soil salinity gradient strongly supported whereas the soil moisture gradient would not be so highlighted in saltmarshes. Nevertheless, the soil moisture gradient would still have a remarkable importance when considering hydrophytic communities. Regarding Tamarix communities, Álvarez-Rogel et al. (2000) included them within the wet saltmarshes, together with perennial succulent communities, further characterised by a higher E.C. Conversely to their reports, our results had showed that saltcedar woodlands appeared in non-succulent zones characterised, in addition, by the lowest E.C. values. These differences could be explained by the high ecological variability in saltcedar woodlands, which can appear from non-saline wetlands, to dry and hypersaline environments (Moreno et al. 2017).
Besides the salinity gradient, the distribution of halophytic plant communities in saltmarshes would be based on the sodicity and the texture gradients (Fig. 5). In the succulent zone, H. strobilaceum communities would occupy soils with lower levels of SAR (considering the effect of E.C.) and higher percentage of sand with regard the remaining succulent communities. Arthrocaulon macrostachyum and S. fruticosa communities would be the dominant succulent vegetation in intermediate values of SAR and soil texture. Conversely, Limonium A zones would appear in sandy soils with low sodicity in non-succulent zones, whereas saltcedar woodlands would be located in soils characterised by the lowest percentage of sand and high sodicity (Fig. 5). The dominant vegetation in non-succulent zones with intermediate values of sand would be Limonium C and B zones, and L. spartum steppes; the latter two plant communities, Limonium B and L. spartum steppes, would share the same ecological requirements (Fig. 5).
Many authors have considered that vegetation patterns in saltmarshes would not be always well defined and markedly structured because of microtopography (Bertness and Ellison 1987; Pennings et al. 2005; Baumberger et al. 2012). In this sense, microtopography would break the edaphic gradient, not allowing a well-defined vegetation structure to appear in bands. However, microtopography does not affect the relationship between vegetation and soil, but only the spatial distribution of soil features, and hence, vegetation. Anyway, even though plots were sampled in zones with homogeneous vegetation and constant topography, microtopography is a factor that must have included variability in our model since it creates heterogeneity in soil conditions.
Further studies have been carried out in saltmarshes from other Mediterranean climatic regions such as California (Peinado et al. 1995). Although soil analyses were not included, these authors compared halophytic plant inventories among different Mediterranean areas, i.e. the west coasts of Europe and Africa, and California territories using cluster dendrograms. Their results provided useful information about halophyte communities and the structure of Mediterranean saltmarshes, suggesting that similar vegetation types appeared in different geographical areas around the world. Besides, similar vegetation patterns have been observed in other non-Mediterranean saltmarshes around the world (e.g. Bertness et al. 1992; Pennings and Bertness 1999; Baumberger et al. 2012), but the importance of particular edaphic factors varied geographically (Cantero et al. 1998; Pennings et al. 2005). Studies of plant zonation in higher latitudes such as New England (U.S.A.) or England (U.K.) showed flooding as the main factor limiting plant distribution (Bertness et al. 1992; pers. obs.). In contrast, studies conducted at lower latitudes showed soil salinity as an important factor mediating plant zonation patterns (Pennings and Callaway 1992; Cantero et al. 1998; Pennings et al. 2005). Therefore, soil salinity would play a significant role in the halophytic vegetation distribution at lower latitudes around the Mediterranean Basin, as has been verified here.
Finally, halonitrophilous shrubs such as Suaeda vera can be found near Limonium and L. spartum areas in saltmarshes (Alonso 2000; Álvarez-Rogel et al. 2000). Suaeda vera was a very common plant species in the studied plant inventories, and it was able to form dense communities in very deteriorated saltmarshes. However, even though this halonitrophilous plant species appeared in many different studied places, this species did not make up a plant group by itself according to the obtained DCA. Suaeda vera generally preferred lower saline soils under conditions of anthropogenic uses such as nitrification and fragmentation (Alonso 2000; Álvarez-Rogel et al. 2000; González-Alcaraz et al. 2014); nevertheless, this species could tolerate high levels of salinity in notably deteriorated saltmarshes and, it could form dense and well-formed plant communities. Further studies focused on these plant communities might be interesting to analyse the role of nitrogen in saline habitats.
Conclusions
Perennial succulent halophytes would appear in the highest saline zones of the saltmarshes, with a dense plant cover during the whole year. Halocnemum strobilaceum would tend to conform dense plant communities over soils to a high percentage of sand and low sodicity (considering the effect of E.C.), whereas Arthrocaulon macrostachyum and Salicornia fruticosa would appear in intermediate values. Different Limonium communities, Lygeum spartum steppes and Tamarix woodlands would be observed at the lowest saline zones in the saltmarshes. Limonium C zones would be related to higher E.C. and ionic concentrations than those for L. spartum steppes, and Limonium B zones would have similar soil conditions as L. spartum steppes. Limonium A communities would appear in sandy soils with low sodicity and the lowest E.C., ionic concentration and soil moisture values. Finally, saltcedar woodlands, formed by Tamarix boveana and T. gallica, would be observed at the lowest salinity zones of the Mediterranean saltmarshes, and their soils were characterised by a low percentage of sand and high sodicity (considering the effect of E.C.).
Our study was performed for a high number of saline habitats throughout the southeast of the Iberian Peninsula under different climates, which would imply a wide distribution both geographically and climatically. Our results might be used to increase the knowledge about plant-soil relationships in saltmarshes around the Mediterranean Basin, complementing the previous studies carried out by Peinado et al. (1995), Álvarez-Rogel et al. (2000, 2001), González-Alcaraz et al. (2014) and Koull and Chehma (2016). Finally, the proposed plant-soil model might be useful to detect the role of halophytes as bioindicator of disturbance, to carry out habitat regeneration according to the species preferences, as Álvarez-Rogel et al. (2001) and González-Alcaraz et al. (2014) suggested, and even to control environmental impacts in saline habitats.
Abbreviations
- ANOVA:
-
Analysis of variance
- CCA:
-
Canonical Correspondence Analysis
- DCA:
-
Detrended Correspondence Analysis
- E.C.:
-
Electrical conductivity
- PAWC:
-
Plant Available Water Capacity
- SAR:
-
Sodium Adsorption Ratio
References
Abdul-Halim MS, Ismail AAM (1990) Vegetation composition of a maritime salt marsh in Qatar in relation to edaphic features. J Veg Sci 1:85–88. https://doi.org/10.2307/3236057
Adams DA (1963) Factors influencing vascular plant zonation in North Carolina salt marshes. Ecology 44(3):445–456. https://doi.org/10.2307/1932523
Alcaraz F, Ortiz R, Hernández S (1987) Contribución al conocimiento de las relaciones suelo-agua-vegetación en un sector de las salinas de Santa Pola (Alicante). Anales de Edafología y Agrobiología 46:273–283
Alonso MA (2000) Estudio geobotánico de los saladares del sureste peninsular (Albacete-Alicante-Almería y Murcia). Ph.D. thesis. University of Alicante, Spain
Álvarez-Rogel J (1997) Relaciones suelo-planta en saladares del sureste de España. Ph.D. thesis. University of Murcia, Spain
Álvarez-Rogel J, Alcaraz F, Ortiz R (2000) Soil salinity and moisture gradients and plant zonation in Mediterranean salt marshes of Southeast Spain. Wetlands 20(2):357–372. https://doi.org/10.1672/0277-5212(2000)020[0357:SSAMGA]2.0.CO;2
Álvarez-Rogel J, Ortiz R, Alcaraz F (2001) Edaphic characterization and soil ionic composition influencing plant zonation in a semiarid Mediterranean salt marsh. Geoderma 99:81–98. https://doi.org/10.1016/S0016-7061(00)00067-7
Álvarez-Rogel J, Jiménez-Cárceles FJ, Roca MJ, Ortiz R (2007) Changes in soils and vegetation in a Mediterranean coastal salt marsh impacted by human activities. Estuar Coast Shelf Sci 73:510–526. https://doi.org/10.1016/j.ecss.2007.02.018
Baumberger T, Affre L, Croze T, Mesléard F (2012) Habitat requirements and population structure of the rare endangered Limonium girardianum in Mediterranean salt marshes. Flora 207:283–293. https://doi.org/10.1016/j.flora.2011.11.008
Bernstein L (1975) Effects of salinity and sodicity on plant growth. Annu Rev Phytopathol 13:295–312
Bertness MD, Ellison AM (1987) Determinants of pattern in a New England salt marsh plant community. Ecol Monogr 52(2):129–147. https://doi.org/10.2307/1942621
Bertness MD, Wikler K, Chatkupt T (1992) Flood tolerance and the distribution of Iva frutescens across New England salt marshes. Oecologia 91:171–178. https://doi.org/10.1007/BF00317780
Blanca G, Cabezudo B, Cueto M, Salazar C, Morales-Torres C (eds) (2011) Flora Vascular de Andalucía Oriental. 2ª edición corregida y aumentada. Universidades de Almería, Granada, Jaén y Málaga, Granada.
Braun-Blanquet J (1946) Über den Deckungswert der Arten in den Pfl anzengesellschaften der Ordnung Vaccinio-Piceetalia. Jahresber Naturforsch Ges Graubündens 130:115–119
Braun-Blanquet J (1979) Fitosociología. Bases para el estudio de las comunidades vegetales. Blume, Madrid
Burt R (2004) Soil survey laboratory methods manual. United States Department of Agriculture (USDA) - Natural Resources Conservation Service (NRCS), Lincoln
Cantero JJ, Cisneros JM, Zobel M, Cantero A (1998) Environmental relationships of vegetation patterns in salt marshes of Central Argentina. Folia Geobot 33:133–145. https://doi.org/10.1007/BF02913341
Castroviejo S (coord gen) (1986–2015) Flora iberica 1–16(I), 17–18, 20–21. Real Jardín Botánico, CSIC, Madrid
Chapman VJ (1939) Studies in salt-marsh ecology. Sections IV and V. J Ecol 27:160–201. https://doi.org/10.2307/2256306
Chapman VJ (1974) Salt marshes and salt desert of the world. 2nd edition. Lehre, Stuttgart
Chigani KH, Khajeddin SJ, Karimzadeh HR (2010) Soil-vegetation relationships of three arid land plant species and their use in rehabilitating degraded sites. Land Degrad Dev 23(1):92–101. https://doi.org/10.1002/ldr.1057
Costa M, Boira H (1981) La vegetación valenciana: los saladares. Anales Jard Bot Madrid 38(l):233–244
Davis MM, Sprecher SW, Wakeley JS, Best GR (1996) Environmental gradients and identification of wetlands in north–Central Florida. Wetlands 16:512–523. https://doi.org/10.1007/BF03161341
Deckers JA, Nachtergaele FO, Spaargaren OC (eds) (1998) World reference base for soil resources. Introduction. ISSS/ISRIC/FAO, Acco, Leuven/Amersfoort
El-Amier YA (2016) Vegetation structure and soil characteristics of five common geophytes in desert of Egypt. Egyptian J Basic Appl Sci 3(2):172–186. https://doi.org/10.1016/j.ejbas.2016.03.001
El-Ghani MA, Soliman A, El-Fattah RA (2014) Spatial distribution and soil characteristics of the vegetation associated with common succulent plants in Egypt. Turk J Bot 38(3):550–565
Engels JG, Jensen K (2010) Role of biotic interactions and physical factors in determining the distribution of marsh species along an estuarine salinity gradient. Oikos 119(4):679–685. https://doi.org/10.1111/j.1600-0706.2009.17940.x
Engels JG, Rink F, Jensen K (2011) Stress tolerance and biotic interactions determine plant zonation patterns in estuarine marshes during seedling emergence and early establishment. J Ecol 99(1):277–287. https://doi.org/10.1111/j.1365-2745.2010.01745.x
Erben M (1993) Limonium Mill. In: Castroviejo S, Aedo C, Cirujano S, Laínz M, Montserrat P, Morales R, Muñoz Garmendia F, Navarro C, Paiva J, Soriano C (eds) Flora iberica 3. Real Jardín Botánico, CSIC, Madrid, pp 2–142
FAO-UNESCO-ISRIC (1988) Soil map of the world, revised legend. World soil resources n°. FAO, Rome, p 60
Flowers TJ, Colmer TD (2008) Salinity tolerance in halophytes. New Phytol 179(4):945–963. https://doi.org/10.1111/j.1469-8137.2008.02531.x
García LV, Marañón T, Moreno A, Clemente L (1993) Above-ground biomass and species richness in a Mediterranean salt marsh. J Veg Sci 4:417–424. https://doi.org/10.2307/3235601
González-Alcaraz MN, Jiménez-Cárceles FJ, Álvarez Y, Álvarez-Rogel J (2014) Gradients of soil salinity and moisture, and plant distribution, in a Mediterranean semiarid saline watershed: a model of soil–plant relationships for contributing to the management. Catena 115:150–158. https://doi.org/10.1016/j.catena.2013.11.011
Gray AJ (1994) Saltmarsh plant ecology: zonation and succession revisited. In: Allen JRL, Pye K (eds) Saltmarshes: Morphodynamics, conservation and engineering significance. Cambridge University Press, New York, pp 63–79
Hackney CT, Brady S, Stemmy L, Boris M, Dennis C, Hancock T, O’Bryon M, Tylton C, Barbeew E (1996) Does intertidal vegetation indicate specific soil and hydrological conditions? Wetlands 16:89–94. https://doi.org/10.1007/BF03160649
Harris DC (2003) Quantitative chemical analysis, 6th edn. W.H. Freeman, New York
Juárez M, Sánchez A, Jordá J, Sánchez J (2004) Diagnóstico del potencial nutritivo del suelo. Universidad de Alicante, Alicante
Kassas M, Zahran MA (1967) On the ecology of the Red Sea littoral salt marsh, Egypt. Ecol Monogr 37:297–315
Koull N, Chehma A (2016) Soil characteristics and plant distribution in saline wetlands of Oued Righ, northeastern Algeria. J Arid Land 8(6):948–959. https://doi.org/10.1007/s40333-016-0060-5
Lambers H, Chapin FS III, Pons TL (1998) Plant physiological ecology. Springer, New York
Lendínez ML (2010) Estudio fitosociológico y fitocenótico de la vegetación halófila andaluza: Bases para su gestión y conservación. Ph.D. thesis. University of Jaén, Spain
Lepš J, Šmilauer P (2014) Multivariate analysis of ecological data using CANOCO 5, 2nd edn. Cambridge University Press, New York
Mateo G, Crespo MB (2009) Manual para la determinación de la flora valenciana, 4th edn. Librería Compás, Alicante
Montasir AH (1943) Soil structure in relation to plants at Mariut. Bull Inst d’Egypte 15:205–236
Moreno J, Terrones A, Alonso MA, Juan A, Crespo MB (2016) Limonium tobarrense (Plumbaginaceae), a new species from the southeastern Iberian Peninsula. Phytotaxa 257(1):61–70. https://doi.org/10.11646/phytotaxa.257.1.4
Moreno J, Terrones A, Juan A, Alonso MA (2017) A quantitative plant-soil model of saltcedar woodlands: the influence of abiotic factors on the floristic composition of Tamarix communities. Catena in press
Moreno J, Terrones A, Alonso MA, Juan A, Crespo MB (2018) Taxonomic revision of the Limonium latebracteatum group (Plumbaginaceae), with the description of a new species. Phytotaxa 333(1):41–57. https://doi.org/10.11646/phytotaxa.333.1.3
Muñoz-Rodríguez AF, Sanjosé I, Márquez-García B, Infante-Izquierdo MD, Polo-Ávila A, Nieva FJJ, Castillo JM (2017) Germination syndromes in response to salinity of Chenopodiaceae halophytes along the intertidal gradient. Aquat Bot 139:48–56. https://doi.org/10.1016/j.aquabot.2017.02.003
Munsell® (1994) Soil colour charts, Revised edition. Macbeth Division of Kollmorgen Instruments Corporation, New Windsor
Neiring WA, Warren RS (1980) Vegetation patterns and processes in New England salt marshes. Bioscience 30(5):301–307. https://doi.org/10.2307/1307853
Ortiz R, Álvarez-Rogel J, Alcaraz F (1995) Soil-vegetation relationships in two coastal salt marshes in southeastern Spain. Arid Soil Res Rehabil 9(4):481–493. https://doi.org/10.1080/15324989509385914
Peinado M, Alcaraz F, Aguirre JL, Delgadillo J, Álvarez J (1995) Similarity of zonation within Californian-Baja Californian and Mediterranean salt marshes. Southwest Nat 40(4):388–405
Pennings SC, Bertness MD (1999) Using latitudinal variation to examine effects of climate on coastal salt marsh pattern and process. Curr Top Wetland Biogeochem 3:100–111
Pennings SC, Callaway RM (1992) Salt marsh plant zonation: the relative importance of competition and physical factors. Ecology 73(2):681–690. https://doi.org/10.2307/1940774
Pennings SC, Grant M, Bertness MD (2005) Plant zonation in low-latitude salt marshes: disentangling the roles of flooding, salinity and competition. J Ecol 93(1):159–167. https://doi.org/10.1111/j.1365-2745.2004.00959.x
Pielou EC, Routledge RD (1976) Salt marsh vegetation: latitudinal gradients in the zonation patterns. Oecologia 24(4):311–321. https://doi.org/10.1007/BF00381137
Piernik A (2003) Inland halophilous vegetation as indicator of soil salinity. Basic Appl Ecol 4:525–536. https://doi.org/10.1078/1439-1791-00154
Piernik A (2012) Ecological patterns of inland salt marsh vegetation in Central Europe. Nicolas Copernicus University Press, Toruń
Pignatti S (1952) Note fitosociologique su alcune associazioni alofile del litorale tunisino. Bol Soc Veneziana Stor Nat 6(1):77–94
Piirainen M, Liebisch O, Kadereit G (2017) Phylogeny, biogeography, systematics and taxonomy of Salicornioideae (Amaranthaceae / Chenopodiaceae) – a cosmopolitan, highly specialized hygrohalophyte lineage dating back to the Oligocene. Taxon 66(1):109–132. https://doi.org/10.12705/661.6
Pujol JA, Calvo JF, Ramírez-Díaz L (2000) Recovery of germination from different osmotic conditions by four halophytes from southeastern Spain. Ann Bot 85:279–286. https://doi.org/10.1006/anbo.1999.1028
R Core Team (2016) R: a language and environment for statistical computing. R Foundation for Statistical Computing, Vienna, Austria. URL https://www.R-project.org/
Redondo S, Rubio-Casal AE, Castillo JM, Luque CJ, Álvarez AA, Luque T, Figueroa ME (2004) Influences of salinity and light on germination of three Sarcocornia taxa with contrasted habitats. Aquat Bot 78:255–264. https://doi.org/10.1016/j.aquabot.2003.11.002
Rivas-Martínez S (2007) Mapa de series, geoseries y geopermaseries de vegetación de España. Memoria del mapa de vegetación potencial de España, Parte I. Itinera Geobot 17:5–436
Sari-Ali A, Benabadji N, Bouazza M (2012) Floristic composition of the halophilic and salt-resistant plant population in Hammam-Boughrara (Oran-Algeria). Open J Ecol 2:96–108. https://doi.org/10.4236/oje.2012.22012
Snow AA, Vince SW (1984) Plant zonation in an Alaskan salt marsh: II. An experimental study of the role of edaphic conditions. J Ecol 72:669–684. https://doi.org/10.2307/2260075
Tabachnick BG, Fidell LS (2007) Using multivariate statistics, 5th edn. Pearson, Boston
Tadros TM (1953) A phytosociological study of halophilous communities from Mareotis (Egypt). Plant Ecol 4(2):102–124. https://doi.org/10.1007/BF00822833
Teege P, Kadereit JW, Kadereit G (2011) Tetraploid European Salicornia species are best interpreted as ecotypes of multiple origin. Flora 206:910–920. https://doi.org/10.1016/j.flora.2011.05.009
ter Braak, C.J., Šmilauer, F.P. (1999) CANOCO for Windows v. 4.02. Centre for Biometry Wageningen CPRO-DLO, Wageningen, The Netherlands
Vicente MJ, Conesa E, Álvarez-Rogel J, Franco JA, Martínez-Sánchez JJ (2007) Effects of various salts on the germination of three perennial salt marsh species. Aquat Bot 87(2):167–170. https://doi.org/10.1016/j.aquabot.2007.04.004
Waisel Y (1972) Biology of halophytes. Academic Press, New York
Wolff WJ (1968) The halophilous vegetation of the lagoons of Mesolonghi, Greece. Vegetatio 16(1–4):95–134. https://doi.org/10.1007/BF00261359
Zedler JB, Callaway JC, Desmond JS, Vivian-Smith G, Williams GD, Sullivan G, Brewster AE, Bradshaw BK (1999) Californian salt marsh vegetation: an improved model of spatial pattern. Ecosystems 2:19–35. https://doi.org/10.1007/s100219900055
Acknowledgements
The authors wish to thank Prof. Francesco de Bello and Prof. Petr Šmilauer for the assistance and suggestions in the statistical analyses; Antonio Sánchez and Margarita Juárez for the assistance in the edaphic analyses; Alicia Vicente, José Luis Villar, Jonás Agulló, Manuel Ortiz and Laura Mora for the assistance in fieldwork; and the University of South Bohemia (Czech Republic) for providing CANOCO v.5 (Microcomputer Power, Ithaca, NY, US) to perform the statistical analyses. We greatly appreciate the comments of two anonymous reviewers. This research was supported by project OAPN 354/2011 (M° de Agricultura, Alimentación y Medio Ambiente, Spanish Government) and FPU grant AP-2012-1954 (M° de Educación, Spanish Government). This research has been supported by the Languages Service (University of Alicante) for the elaboration of Ph.D. Theses in Valencian and foreign languages. This research is part of the Ph.D. Thesis of Joaquín Moreno.
Author information
Authors and Affiliations
Corresponding author
Additional information
Responsible Editor: Wieland Fricke.
Electronic supplementary material
ESM 1
(XLSX 13.2 kb)
ESM 2
(XLSX 13.5 kb)
ESM 3
(XLSX 28.3 kb)
ESM 4
(XLSX 62 kb)
ESM 5
(XLSX 16.3 kb)
ESM 6
(XLSX 13.7 kb)
ESM 7
(XLSX 12.7 kb)
ESM 8
(XLSX 16.7 kb)
ESM 9
(XLSX 15.1 kb)
Figure A1
Canonical Correspondence Analysis (CCA) with E.C. as covariable of forty-three samples of halophytic communities from studied Mediterranean saltmarshes showing correlations between samples and edaphic variables. Arrows indicate the edaphic variables and their directions and length show their relationships to the ordination axes. Edaphic variables abbreviations: E.C., electrical conductivity; Moisture, soil moisture; PAWC, plant available water capacity; SAR, sodium adsorption ratio. Plant community abbreviations: A. macrostachyum, Arthrocaulon macrostachyum; H. strobilaceum, Halocnemum strobilaceum; L. spartum, Lygeum spartum; S. fruticosa, Salicornia fruticosa. (PDF 56 kb)
Rights and permissions
About this article

Cite this article
Moreno, J., Terrones, A., Juan, A. et al. Halophytic plant community patterns in Mediterranean saltmarshes: shedding light on the connection between abiotic factors and the distribution of halophytes. Plant Soil 430, 185–204 (2018). https://doi.org/10.1007/s11104-018-3671-0
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11104-018-3671-0