
Abstract
Aims
A simulation model to demonstrate that soil water potential can regulate transpiration, by influencing leaf water potential and/or inducing root production of chemical signals that are transported to the leaves.
Methods
Signalling impacts on the relationship between soil water potential and transpiration were simulated by coupling a 3D model for water flow in soil, into and through roots (Javaux et al. 2008) with a model for xylem transport of chemicals (produced as a function of local root water potential). Stomatal conductance was regulated by simulated leaf water potential (H) and/or foliar chemical signal concentrations (C; H + C). Split-root experiments were simulated by varying transpiration demands and irrigation placement.
Results
While regulation of stomatal conductance by chemical transport was unstable and oscillatory, simulated transpiration over time and root water uptake from the two soil compartments were similar for both H and H + C regulation. Increased stomatal sensitivity more strongly decreased transpiration, and decreased threshold root water potential (below which a chemical signal is produced) delayed transpiration reduction.
Conclusions
Although simulations with H + C regulation qualitatively reproduced transpiration of plants exposed to partial rootzone drying (PRD), long-term effects seemed negligible. Moreover, most transpiration responses to PRD could be explained by hydraulic signalling alone.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Plants lose large amounts of water to the atmosphere through stomata that open for CO2 uptake and carbon assimilation. This water loss is compensated by water uptake from the soil. When soil dries, there may be damaging effects of root shrinkage and subsequent loss of contact with the surrounding soil, and the low plant water potentials required to maintain soil water uptake may induce xylem cavitation (Barigah et al. 2013). However, effective stomatal regulation prevents excessive loss of water when the soil dries out (Brodribb and McAdam 2011). This mechanism represents the main short-term regulation of water flow between the soil and the atmosphere (Taiz and Zeiger 2006).
In many experiments, leaf water potential is correlated with stomatal closure (Buckley 2005). One obvious strategy to avoid cavitation is to close stomata to maintain the leaf water potential (ΨL) at a certain level. This is called pressure homeostasis (isohydric behaviour). On the other hand, in some species, stomata are insensitive to changes in ΨL within a certain range, and their stomata remain fully open during soil drying and higher transpiration demand. Large diurnal variations of ΨL may occur, leading to so-called anisohydric behaviour (Franks et al. 2007), in contrast to isohydric species where low soil water potentials cause stomatal closure which maintains ΨL. The hydraulics of the root and shoot system provide a direct link between root-zone water potential, transpiration flow and leaf water potential and may influence long-distance hydraulic signalling thereby linking plant water potentials and stomatal closure (Christmann et al. 2013).
However, plants have also developed non-hydraulic strategies to regulate transpiration (Tardieu and Simonneau 1998). Experiments that split the roots between two soil compartments have suggested that hydraulics alone cannot explain certain stomatal responses. Although drying part of the root system decreases stomatal conductance in the absence of changes in leaf water potential, continued drying of parts of the rootzone actually increases stomatal conductance (Khalil and Grace 1993; Stoll et al. 2000). These effects were proposed to result from the roots in drying soil synthesizing a chemical signal which is transported from the roots to limit stomatal conductance, but then stomata re-open when signal transport is restricted as the soil becomes too dry. These split-root experiments have been upscaled to implement partial rootzone drying as a water-saving irrigation technique to improve crop water use efficiency, with periodic alternation of wet and dry sides of the rootzone to ensure continued signal transport to the shoots (Bravdo 2005; Dodd et al. 2006; Kang 2004; Stoll et al. 2000).
The production, transport and release of chemicals such as the phytohormone abscisic acid (ABA) or hormone precursors such as 1-aminocyclopropane-1-carboxylic acid (ACC - the ethylene precursor) into leaves (Tardieu and Davies 1993; Wan and Zwiazek 2001) can be critical in stomatal regulation (Davies and Zhang 1991). However, the source of chemical signals that are important for stomatal regulation is contentious. Early evidence for root ABA production (Hartung and Aboumandour 1980) was supported by later observations that xylem ABA concentrations increase significantly under water stress conditions and correlate with stomatal closure (Schurr et al. 1992). Furthermore, soil drying increases xylem sap pH, which enriches ABA in the leaf apoplast by decreasing ABA uptake by mesophyll cells (Wilkinson and Davies 1997). These mechanisms suggest that root sourced ABA can act as a long-distance signal initiating stomatal closure.
In contrast, other studies cast doubt on the hypothesis of root sourced ABA acting as a long-distance signal for stomatal closure. Reciprocal- and self-grafts of wild-type and ABA-deficient tomatoes demonstrated that stomatal regulation depends on the shoot genotype only, and is independent of the rootstock (Dodd et al. 2009; Holbrook et al. 2002). Furthermore, osmotic stress applied to the roots reveals ABA-dependent reporter gene expressions only in the leaves (Christmann et al. 2007). Those experiments suggest that leaves are the main source of ABA, and a subsequent basipetal transport of ABA via the phloem towards the roots, where the hormones are recycled to the xylem and transported upwards again. However, the proportional contributions of ABA recycling and root synthesis to xylem ABA concentration vary according to the root environment (Wolf et al. 1990). Mechanistically, both sources of an additional chemical signal can be (mathematically) described by the same approach, namely a production or a release of the chemical within the roots into the xylem and a subsequent transport to the leaves.
In hydrological models, signalling between rootzone conditions and stomatal closure is not modelled explicitly but stomatal regulation in response to low soil water potentials is implicitly accounted for through “stress functions”. These functions relate the reduction of the potential transpiration rate directly to bulk soil water potential (instantaneous effect) thereby bypassing other variables that control stomatal conductance and root-to-shoot signalling mechanisms . An oft-used stress function is the Feddes function which describes the transpiration reduction factor as a piece-wise linear function of soil water potential (Feddes et al. 1978). The decreasing part of the function for low soil water potential implicitly represents the effect of soil water potential on stomatal closure. However, this relationship is indirect and may therefore depend on other factors. For instance, for the same bulk soil water potential, lower leaf water potential may be expected with higher transpiration rates. On the other hand, when production rate of the hormonal signal is not a function of sap flow rate, a higher transpiration rate will dilute the signal, causing lower signal concentrations in the leaves. Depending on which mechanism relates soil water potential to stomatal conductance, the impact of higher transpiration rate on the onset of stomatal closure will be different.
As discussed above, several models link stomatal conductance to environmental factors. However, most of these models “do not include satisfactorily the effects of drought, impairing our capacity to simulate plant functioning in conditions of limited water supply” (Damour et al. 2010). Here, a new model is presented that combines both hydraulic and chemical signalling processes into a common modelling framework. A 3D model that explicitly solves the water flow equations in the root system (Javaux et al. 2008) was coupled to a model that simulates signal production in the rootzone and transport through the root system to the leaves. Hereby, the stem and leaves were not explicitly considered but only modelled as an additional root segment with the same volume as the total root system. First, how the different signalling mechanisms are implemented in the model is discussed. Secondly, the effect of different signalling processes on the relation between soil water potential and transpiration is demonstrated and a sensitivity analysis presented. Finally, water and chemical flows from different parts of the rootzone of split-root plants, and whole plant transpiration were modelled during alternate partial rootzone drying. This study intends to (i) simulate observed plant behaviour during partial rootzone drying and (ii) show the impact of a chemical signal on the relation between soil water potential and transpiration reduction.
Model approach
Definition of water potential, water pressure, and pressure head
Plants take up water from the soil through their roots and transport it to the leaves where it is transpired through the stomata. The driver for this transport is the difference in water potential (a measure of the energy state of water) between the atmosphere and the soil. The energy of the plant (or soil) water or water potential can be either stated as total water pressure (Ψ [Pa], energy per volume) or as hydraulic head (H [m], energy per weight). Total water pressure and hydraulic head can be converted using Eq. (1). Gradients in total water pressure or hydraulic head are relevant for driving water flow. The total water pressure and hydraulic head represent the sum of different partial potentials, including the gravitation potential or the energy of water due to its elevation above a certain reference height.
where ρ is the density of water [kg m−3] assumed to be constant, g the acceleration due to gravity [m s−2], and z [m] is the height above a reference level. By approximating g as 10 m s−1 and ρ as1000 kg m−3, we can state that h (m) ≈ 10,000 ψ [Pa] or that a pressure head of 1 cm approximates a pressure of 1*10−4 MPa.
Concept of stomatal conductance
Stomatal closure is the main short-term control mechanism that decreases actual transpiration (Tact) as the soil dries. The potential transpiration (Tpot) represents the transpiration of the plant under the same climatic conditions but in a well-watered soil. Following Tardieu and Simonneau (1998), who classified stomatal sensitivities for different plant species, anisohydric behaviour refers to stomata that remain fully open during drying until the plant reaches its permanent wilting point. The leaf pressure head, hL, of anisohydric plants thus shows strong fluctuations with changes in soil water availability and transpiration rates (e.g. diurnal or seasonal cycles). In contrast, isohydric plants try to regulate hL and keep it higher than or equal to a certain threshold which is higher than the permanent wilting point value, thereby reducing Tact when soil is drying. We chose an adapted version of the Tardieu and Davies (1993) model that describes stomatal conductance as a function of the internal plant variables hL and concentration of chemical signal in the leaves.
Relative stomatal conductance α, relative to the maximum conductance under the same conditions but when water content is optimal, was calculated using:
where αR [-] is the residual relative stomatal aperture, cL [nmol cm−3] is the concentration of chemical signal in the leaf, hL is the leaf pressure head and hcrit is a threshold pressure head in the leaf, sh [cm−1] and sc [cm3 nmol−1] are fitting parameters for pressure head and signal concentration respectively. The variable cd is a Boolean variable that either enables only pressure head regulation (cd = 1) or an interaction between chemical and pressure regulation (cd = 0). Equation 2a is valid for h L < h crit , otherwise
where at concentration cL = 0, α will be equal to 1. Note that α corresponds with the ratio of actual transpiration, Tact, to the potential transpiration, Tpot, with values between αR (residual stomatal conductance) and 1. The effect of both hydraulic and chemical signalling (‘H + C’ = hydraulic and chemical control) on the transpiration reduction as described by Eq. (2) is illustrated (Fig. 1). While keeping the concentration of the chemical constant, α is plotted versus hL (parameters are detailed in Table 1). Different lines represent α for different signal concentrations. Lower leaf water potentials as well as higher chemical concentrations decrease transpiration. An anisohydric plant (‘NR’ = No Regulation) keeps the ratio α at 1 and thus actual transpiration equals potential transpiration until the permanent wilting point is reached (acc. Eq. 2, this would correspond to sc = 0 and hCrit = hWP). For the case sh = 0 and sc > 0, the leaf conductance does not directly depend on hL and reduces only with increasing concentrations (‘C’ = chemical control). With chemical control only, it should be noted that an indirect relation between stomatal conductance and leaf water potential follows from the fact that the chemical signal is produced as a function of root water potential which is hydraulically linked to leaf water potential.

General behaviour of Eq. 2 over varying leaf water potentials, where multiple lines refer to different signal concentrations (arrows indicate towards higher concentrations; (0.5, 1, 2)*10-3 nmol cm-3). NR: No regulation, H + C: hydraulic and chemical control (sc = 5*1010 cm3 mol-1, sh = 1*10−3 cm−1, aR = 0.1; hcrit = -5,500 cm); H: hydraulic control (sc = 0; sh = 1*1010; aR = 0.1, hcrit = -5,500 cm), C: chemical control (sc = 5*1,010; sh = 0, aR = 0.1)
Another extreme case would be a hydraulically controlled plant (‘H’ = hydraulic control) in which the leaf conductance remains constant as long as the leaf pressure head, hL, stays above a critical pressure head, hcrit. When hL equals hcrit, stomatal conductance is reduced until the transpiration from the leaves to the atmosphere matches the transpiration stream in the plant that is driven by the hydraulic head difference between the rootzone and the leaves where the pressure head equals hcrit. This corresponds with cd = 1 and sh equal to infinity so that Eq. 2 is a step function in terms of hL. Such step functions are often used to describe stomatal closure in hydrological models (Feddes et al. 1978).
In Fig. 1, the curves shown represent lines on the α (hL, cL) surface that are obtained when the signal concentration cL is kept constant. These curves demonstrate the behaviour of the α (hL, cL) function but do not represent a projection of the trajectory of α (hL, cL) on the hL axis during a drying event, in which both hL and cL change over time. Thus these curves only represent the stomatal response for a single concentration value.
Hydraulic signalling
R-SWMS (Javaux et al. 2008) calculates the pressure head in the leaves, hL, by solving the hydraulic equations within the xylem network, between the xylem and the soil-root interface, and between the soil-root interface and the bulk soil. The water flow distribution in the soil and root systems can be predicted in 3-D by solving respectively the Richards (1931) and the Doussan et al. (2006) equations. Radial uptake Jr [m3 s−1] and axial flow within the root xylem, Jx [m3 s−1] are defined for each root segment, i, by the Doussan et al. (2006) equations:
where Kr * [s−1] is the radial conductivity, Ar [m2] the root outer surface, hs,int the soil pressure head at the root-soil interface and hx the pressure head in the root xylem, Kx * [m s−1] the xylem hydraulic conductivity, Ax [m2] the root cross-sectional area, Δhx the difference in pressure head within the root segment, Δz [m] the difference in elevation, and li, the length of the root segment. The R-SWMS model (Javaux et al. 2008; Schroder et al. 2009, 2012) was used to solve these equations numerically by coupling, on a voxel basis, the sink term of the Richards equation to the root water uptake (∑Jr). Thus distributions of pressure heads within the soil and the root system and the root-water-uptake distribution within the soil could be predicted.
The shoot and leaves were effectively modelled with a segment connected to the root system collar. The length of this segment was relatively short so that the pressure head drop across this element was small. The simulated leaf pressure heads were therefore similar to the pressure heads at the root collar. By increasing the effective length or reducing the xylem conductivity of this segment, a larger pressure head difference between root collar and leaves can be simulated. However, accounting for an extra pressure drop between the root collar and the leaves will not influence the effect of different signalling mechanisms.
Chemical signalling
Chemical signalling refers to the loading of chemical signals into the xylem of plant roots as a function of rootzone water potential, their transport in the xylem to the shoot, and the regulation of stomatal conductance by their concentrations in the leaves. Thus production of chemicals in the leaves as a function of the leaf pressure head is not considered as chemical signalling between roots and leaves. Rootzone conditions and stomatal conductance are in this case related by hydraulic signalling. The concentration cL in our model therefore only refers to the concentration of chemicals in the leaves that originate from the roots. This implies that the function α (hL, cL) cannot be parameterized based on chemical concentrations that are measured in the leaves.
-
a.
Production of chemicals in the roots
The chemical production rate in a segment i, MSignal,i [mol d−1], is modelled as a function of the pressure head in the root xylem, hRoot,i, and the dry mass of the root segment mi (gDM) following Dodd et al. (2010) and Liu et al. (2005).
$$ {M}_{signal, i}=\left\{\begin{array}{cc}\hfill 0\hfill & \hfill for\left|{h}_{Root, i}\right|<\left|{h}_0\right|\hfill \\ {}\hfill \mathrm{a}\left(\left|{h}_{Root, i}\right|-\left|{h}_0\right|\right){m}_i\hfill & \hfill for\left|{h}_{Root, i}\right|\ge \left|{h}_0\right|\hfill \end{array}\right. $$(4)where a [mol gDM −1 m−1 d−1] is the production rate per dry mass of the root segment and pressure head, and h0 [m] a threshold pressure head, below which production is started. This threshold was introduced to avoid hormone production for unstressed conditions and to lower computational costs for the particle tracking algorithm, as explained below. As ABA production was independent of root type or age but dependent on root water potential (Simonneau et al. 1998), a and h0 are considered to remain constant in time.
-
b.
Hormone transport to the leaves
Hormone transport in the root system was simulated using a particle tracking algorithm. At each time step, Δt, in any particular segment i, when |hRoot,i| ≥ |h0|, one particle with the mass Msignal,i*Δt is generated. This particle is transported with the root water flow towards the leaves. By knowing the water flow through the root system, the xylem flow velocity vR,i in a given root segment, is calculated by Eq. 5.
$$ {v}_{R, i}=\frac{J_{x, i}}{A_{x, i}} $$(5)where Jx,i [m3 d−1] is the axial water flow through the root segment i, and Ax,i [m2] is the cross section of the segment.
To account for particle mixing in the shoot, this segment was modelled as a perfectly mixed buffer with a volume, VBuffer, that was a predefined multiple or fraction of the total root volume, VRoot (Eq. 6), representing the shoot volume. For herbaceous plants, the root:shoot ratio decreases with age and is strongly influenced by environmental conditions (Wilson 1988). Crop growth models (e.g. gecros) assume variations over a plant life from 0.5 to 2.0 (Yin and van Laar 2005). A reference buffer size, equalling the total root volume (root:shoot = 1.0), was assumed. A transpiration rate of 10 cm3d−1 and a buffer volume of around 1 cm3 (plant age around 20 days) leads to an average residence time of the chemical in the buffer of around 0.1 days, within the range of previously observed half-life of ABA in stressed plants (Liang et al. 1997). The concentration in this uppermost segment, cL at time tj + Δt was calculated from the concentration at time tj as:
where Σmass is the mass of all particles that arrive in the upper segment during time tj and tj + Δt.
To evaluate the impact of chemical transport from the roots to the leaves, a model in which root produced hormones are assumed to arrive instantaneously at the leaves (‘H + C, i’) was considered, so that cL [nmol cm−3] can be calculated as the sum of the root tip production rates, ΣM signal,i = M signal,tot , divided by the actual transpiration (Eq. 7). In this approach it is not necessary to simulate transport through the root system.
Materials and methods
Virtual experiments
First, a split root experiment in which one part of the rootzone was dried was simulated to compare different mechanisms that relate heterogeneous rootzone conditions to stomatal closure and transpiration reduction: hydraulic (H), chemical transport (C), the combination of both (H + C), and a hypothetical instantaneous chemical signal (H + C,i). Second, to evaluate the model’s general behaviour and its response to changes in the parameters of the stomatal model (s c , s h , h 0 ) (Eq. 2a), a sensitivity analysis was carried out by varying each parameter by one order of magnitude. Additionally shoot buffer size was varied from 0.02 to 2*V Root (see Appendix 1). Third, the impact of chemical versus hydraulic signalling on transpiration when rootzone drying was alternated between the two soil compartments was tested for a constant and for a transient, diurnal, transpiration demand.
For all simulations, daily irrigation rates as well as the daily transpiration demand (Tpot) remained the same. Different responses were thus due to a different parameterization (sensitivity analysis) and/or due to different signalling mechanisms (H, C, H + C, H + C,i) . Details of the soil compartments, root system and boundary conditions are given below. Parameterization of the setups and sensitivity analyses are given in Table 1 and Table 2 respectively.
Soil domain
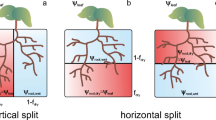
The soil domain (Fig. 2) of 7 × 3 × 15.5 cm3 was subdivided by a grid with voxel sizes of 0.25 × 0.5 × 0.5 cm3. To simulate spatially variable water application leading to contrasting water distribution over the rootzone, the domain was split in two equal parts, separated by a non-conductive layer in the y-plane with a width of 4 voxels (Table 3, #2). Soil parameters for the two compartments were set to a clay loam (Table 3, #1) taken from Carsel and Parrish (1988).

3D distribution of soil water content within the soil domain [cm3 cm−3] at Day 25, after stopping irrigation of the right compartment, together with distribution of water potential in the root xylem [cm] ≈ ψ [10−4 MPa]. The soil domain was 7×3×15.5 cm3. The detail shows the particles, representing a chemical signal, within the roots
Root system
The root system was considered to be static (non-growing) during the simulation (25–40 days), to focus on non-growth related plant responses to water deficit. The split-root system was generated by using the root growth module within R-SWMS (Fig. 2), as previously described (Clausnitzer and Hopmans 1994; Somma et al. 1998). Both soil compartments have an equal number of root tips but root structures are not mirror images of each other as the growth model includes a random component. With the same input configuration, two subsequent runs of the growth model result in different root structures. Since the root surface in the two compartments differed by 1.8 % and the root length by 2.1 %, simulated root water uptake rates from the two compartments deviated by 1.2 % when uniform irrigation was supplied to both soil compartments.
Boundary and initial conditions
The upper soil boundary condition was set to a flow boundary (Cauchy type). Both soil compartments were independently, uniformly irrigated during the first 5 days (each side received 5.85 cm3d−1, irrigation rate = 0.65 cm d−1). On subsequent days, one side of the root system received 11.7 cm3d−1 whereas the other compartment was not irrigated. Irrigation of the two soil compartments was sometimes alternated, as in PRD experiments under field conditions. Lateral walls of the soil domain were set to a no-flow condition and the bottom to free drainage, i.e. dh/dz = 0.
To solve the flow equations in the root system, the transpiration rate, Tact = αTpot, was set as a flow boundary condition at the upper segment connected to the root network. The potential transpiration rate was either kept constant over time with Tpot = 10 cm3d−1or transient with a diurnal cycle that was modelled as a truncated sine with zero transpiration during night (12 h). For the C, H + C and H + C,i scenarios, the relative stomatal conductance α was derived from the pressure head and chemical signal concentration in the leaves. For the H scenario, a flow boundary condition was used with α = 1 for hL > hcrit. When hL = hcrit, the boundary condition was switched to a so-called Dirichlet condition for which hL was kept at a constant pressure head hcrit.
The steady state water pressure heads obtained under uniform irrigation were used as initial condition. Simulation durations, with varying irrigation regimes and stomatal regulation, ranged from 25 to 40 days.
Definition of equivalent rootzone pressure head
Typically in hydrological models a piece-wise linear relation between rootzone pressure head and transpiration rate (Feddes et al. 1978) is used to relate the reduction of transpiration and soil water potential. To investigate the effect of chemical signalling on this relationship, we calculated averaged rootzone pressure heads in the entire rootzone and in each of the two compartments of the split root experiment.
Couvreur et al. (2012) derived a procedure to define an equivalent rootzone hydraulic head hS,equiv that takes the root architecture into account by only considering soil voxels that are affected by root water uptake:
where the subscript k stands for the kth soil voxel and sk [-] is the standardized sink fraction (SSF [-]) that corresponds to fraction of root water uptake from this voxel for the case that the hydraulic heads (h + z) are uniformly distributed in the rootzone (Couvreur et al. 2012).
Results and discussion
Influence of stomatal regulation mechanisms
Initially, the effect of different mechanisms (NR, H, C, H + C) that relate the rootzone water pressure head with stomatal conductance on the pressure head in the leaves and on transpiration rate were simulated (Fig. 3). During the first five days, when irrigation was applied uniformly to both soil compartments, the plant was unstressed (Tact = Tpot, Fig. 3b). After switching to non-uniform irrigation (Day 5), while keeping the amount of applied water constant, the pressure head at the leaves started to drop (Fig. 3a). Until approximately Day 9, the pressure heads for the different mechanisms are similar and actual transpiration rates equal potential transpiration rates. Water uptake from the drying compartment progressively decreased until it ceased around Day 18, when the irrigated compartment alone supported transpiration (Fig. 3d).

Responses of the system: a leaf water potential and b transpiration rate, and c equivalent soil water potential and d) water uptake from the two soil compartments for different mechanisms that link rootzone water potential with stomatal conductance and transpiration: NR = no reduction of transpiration, H = hydraulic control, C = Chemical control, H + C = hydraulic and chemical control, and H + C,i = hydraulic and instantaneous chemical signallingrootzone
For the NR case, after Day 9 transpiration remained constant over time but the pressure head at the leaves declined until about Day 20 when it reached a steady state (ca. −6,400 cm ≈ −0.64 MPa, Eq. 1). When only one compartment is irrigated, the same volume of water must be taken up and transported to the leaves through fewer roots, thus the pressure head drop between the soil and leaves becomes larger and the pressure head at the leaves smaller (Fig. 3c).
For the H case, stomatal conductance regulates the pressure head in the leaves and keeps it constant when hcrit = -5,500 cm is reached. Thus the pressure head at the leaves cannot decrease to the value (−6,400 cm) required to maintain a steady transpiration rate when only one compartment of the rootzone is irrigated. Consequently, transpiration starts to decrease when the pressure head at the leaves becomes −5,500 cm, until it reaches a new steady value after around 25 days.
For two of the chemical signalling cases (C, H + C), transpiration was reduced earlier than in purely hydraulic regulation (H). Since chemicals were produced at pressure heads in the rootzone below −4,500 cm (Table 1, h0), chemical signalling started to decrease stomatal conductance and transpiration for leaf pressure heads slightly smaller than −4,500 cm in the case of instantaneous transport (H + C,i) and more negative values when convective transport in the plant root system was considered. Although there are some differences between chemical (C and H + C) and hydraulic (H) controls, the transpiration rate as well as the pressure head at the leaves were similar when steady state was reached (data not shown). Responses to an instantaneous chemical signal (H + C,i, Eq. 6) were comparable to hydraulic only signalling (H) but with a higher steady state pressure head of around −4,800 cm.
Generally, simulated transpiration rate evolved similarly with time for the different cases. However, transpiration rate monotonically declines for the H and H + C,i cases whereas for C and H + C cases it starts to gradually increase again at around Day 15 (Fig. 3b). This recovery of transpiration during uninterrupted drying of one part of the root system was previously reported from split-root experiments (Khalil and Grace 1993; Stoll et al. 2000). Our simulations indicate that it could be explained by the decline in water uptake from the drying compartment (Fig. 3d) and, for the C and H + C case, the corresponding decrease in transport of chemicals produced in the drying compartment. After reaching a peak at Day 14, the mass flux of chemical signals from the dry root system to the leaves declines again (Fig. 4), as demonstrated experimentally (Dodd et al. 2008b). When comparing transpiration rates (Fig. 3b), transport limitation of chemical signals produced in the drying roots (H + C) leads to 50 % less transpiration reduction than in the case of instantaneous chemical signal transfer to the shoot (H + C,i).

Concentration effect: Chemical signal arriving at the leaves from the dry (dashed line) and the irrigated (dash-dotted line) compartment and total (solid line). a Comparison of mass fluxes and b concentrations of chemical signal for the H + C case
Effect of chemical signal concentration and linkage to soil pressure head
These simulations allow chemical signal concentrations and mass fluxes coming from the two parts of the split root system and arriving at the shoot to be examined (Fig. 4). A signal concentration range between 1*10−4 and 1 nmol cm−3 (1 nmol cm−3 = 1 μM or 1,000 nM) matches the observed concentration ranges of ABA in xylem sap (Li et al. 2011; Martin-Vertedor and Dodd 2011; Stoll et al. 2000). Signal concentration in the transpiration stream arriving from the dry side is nearly three orders of magnitude higher than the concentration coming from the irrigated side (Fig. 4). Ranges of two orders of magnitude were observed experimentally (Dodd et al. 2008b). However, the mass flow of chemical arriving from the dry compartment is only about twice as high as the flow from the irrigated compartment, due to limited transport of chemical signals out of roots in dry soil (Fig. 4a). The model also simulated chemical signal production in the irrigated rootzone, which considerably contributed (ca. 30 %) to the total mass that arrived in the shoot (Fig. 4a).
The distribution of pressure heads in the root xylem and in the soil voxels where water is taken up for a uniform hydraulic head distribution (SSF > 0) is shown before and after the partial irrigation started for the H + C scenario (Fig. 5). Since root water uptake in the irrigated part increased, the pressure difference between soil and root xylem increased and the pressure head in the roots decreased in the irrigated part of the rootzone even though the equivalent soil water pressure head increased. Furthermore, the pressure heads in the roots varied considerably so that the root pressure head decreased below h0 in parts of the irrigated root system (Fig. 5a) and thus chemical signal was produced. This illustrates that, for this study, signal production in the irrigated part of the rootzone, which is related to the local water pressure head in the roots, cannot be directly linked to the equivalent soil water pressure in that part. This missing link was previously observed in an experimental study (Puértolas et al. 2013). Since the flow from the non-irrigated part ceased, the pressure heads in the non-irrigated part converged to the leaf pressure heads (Fig. 5b, see also Fig. 3a–c).

Comparison of root xylem pressure head h [cm] ≈ ψ [10 − 4 MPa] and soil potential (only soil voxels where SSF > 0) for H + C at uniform irrigation (Day 1, left side) and for non-uniform irrigation (Day 25)
Effects of signal transport
Several authors attempted to link soil water content to xylem ABA concentrations (Dodd et al. 2008b; Liu et al. 2008) by assuming that soil water content was the driver for ABA production. Thus signal concentration in the shoot was plotted against the degree of (soil water) saturation (S = (θ - θR)/(θS - θR)) of the drying compartment (Fig. 6). For comparison, signal concentrations were calculated by injecting all rootzone produced chemicals instantaneously into the shoot. In this case, signal concentration continuously increased and was consistently one order of magnitude higher. When transport of the chemical signal with xylem water flow in the root system was simulated, a peak concentration was reached at a relative saturation of around 0.27, as in split-root experiments demonstrating a peak shoot hormone concentration at an intermediate soil water content of the drying compartment (Dodd et al. 2008b; Liu et al. 2008). Thus linearly relating hormone production to root pressure head may lead to this type of behaviour when transport of chemical signals from a drying part of the rootzone becomes limiting.

Concentrations of the chemical signal in the leaves versus the degree of saturation of the drying compartment for the case that the rootzone produced chemicals are transported with the water flow from root system to the leaves (‘with transport’) and for the case that the rootzone produced chemicals are instantaneously injected in the shoot (‘instantaneous’) (Eq. 7)
Sensitivity analysis
Stomatal conductance and resulting actual transpiration rate was modelled versus time and versus the equivalent soil water pressure head for different values of parameters used in Eq. 2a (Fig. 7). Both sensitivities of this equation, sh and sc influence the magnitude of the transpiration reduction but not when transpiration is reduced (Fig. 7 a). A higher sensitivity of stomatal conductance to chemical signal concentration (larger sc) decreases transpiration to a greater degree. The sensitivity to the pressure head is smaller and the influence of the pressure term in Eq. 2a is negligible when sh becomes smaller than 1*10−4 cm−1. The onset of reduction is related to the threshold for the release or production of chemical signals in the roots, h0. This parameter also influences the magnitude of transpiration reduction since it determines the chemical production rate as function of the root water pressure head.

Sensitivity analysis of the parameters of Eq. 2: a Transpiration rate over time and b transpiration reduction (α = Tact/Tpot) over equivalent soil water potential
When a threshold pressure in the leaves is maintained (H case), linear piece-wise relations (step functions) between transpiration reduction and the equivalent soil water pressure head as defined in Eq. (8), which represents a soil water pressure head sensed by the roots, are obtained for a given potential transpiration rate (Couvreur et al. 2012; Javaux et al. 2013). This agrees with the functions typically used in soil water flow models to describe the reduction of transpiration as a function of soil water pressure head (Molz 1981). If an additional chemical signal is considered, similar piece-wise linear relations between transpiration reduction and equivalent soil pressure head are obtained (Fig. 7b). The slope of the α (hS,equiv) increases with increasing sc and sh and the onset of the transpiration reduction shifts towards lower equivalent soil water pressures for smaller sh values. These results suggest that the effect of chemical signalling may be approximated by functions that relate transpiration reduction directly to soil water pressure head and that similar functions are obtained when only hydraulic signalling is considered. However, despite their similarity to piece-wise linear functions, the relationships obtained for chemical signalling (when soil pressure head is related to transpiration reduction) show some peculiarities when transport of chemicals produced in dry rootzones becomes limited. As observed experimentally, the relationship loses its linearity and transpiration recovers when chemicals produced in the dry rootzone are no longer transported to the shoot (Dodd et al. 2008a; Liu et al. 2008), thus increasing total root water uptake with decreasing equivalent soil water pressure head.
The influence of shoot volume on transpiration is explored in the Appendix.
Alternated partial rootzone drying
Constant transpiration demand
When irrigation was alternated between wet and dry soil compartments after 15 days, Simulation 1 regulated stomata only as a function of leaf pressure head (H) while Simulation 2 included an additional chemical signal (H + C). Additional simulations of C and H + C,i were not included as responses were similar to the H + C and the H case, respectively (see also Fig. 3). During the first five days, when irrigation was applied uniformly to both soil compartments, actual transpiration equalled the potential transpiration (Fig. 8a) in both simulations. Transpiration began to decline about four days after changing to partial irrigation. The general course of transpiration is similar for the H and H + C cases, even though transpiration was less in the H + C case, indicating that both hydraulic and chemical signalling can produce similar plant responses during alternate PRD.

Partial rootzone drying: a Actual transpiration rate for regulation as a function of hydraulic signalling (H) and of hydraulic + chemical signalling (H + C), Tpot remained at 10 cm3 d−1. Irrigation was either applied uniformly or partially at the right or the left compartment. b Equivalent soil water pressure head for the total soil domain for H and H + C. c Concentration of chemical signal in the shoot and mass flux of chemical arriving at the shoot from both compartments. Values are averaged over 0.1 days
However, immediately after alternating irrigation at Day 20, the transpiration rate of H + C sharply decreased, associated with an increased signal concentration in the shoot (Fig. 8c, black solid line). After an initial drop, signal mass flux from the previously non-irrigated root system showed a distinct peak for about a day. With continued re-wetting of this compartment (Fig. 8b) and increased water uptake from roots contained therein, chemicals accumulated in the roots can again be transported to the shoot. Interestingly, signal mass flux from the now-drying compartment is reduced. As this compartment still contains water from the previous irrigation cycle, and the other compartment is now irrigated, a larger part of the root system can contribute to water uptake (Fig. 8b). This leads to a more homogeneous distribution of soil water pressures and thus a relaxation of the total root system with higher root water potentials than under stress conditions.
Increased transport of chemical signals upon rewetting part of the rootzone (presumably originating from previously stressed parts of the rootzone) was observed in field experiments with grapevine in the morning shortly after the onset of transpiration (Romero et al. 2012) and in pot-grown tomato plants for 8 h after alternating irrigation (Dodd et al. 2006). Production and accumulation of chemical signals during periods without root water uptake (e.g. during night or in non-irrigated parts of the rootzone) and subsequent transport when root water uptake starts again (e.g. in the morning or re-wetting of roots in dry soil) may be critical in regulating transpiration of plants exposed to PRD, and may contribute to the agronomic advantages of alternated partial rootzone drying over deficit irrigation (Dodd 2009).
Transient transpiration demand
To obtain more realistic conditions, the previously described scenario was modified by applying a diurnal, sinusoidal transpiration demand and an alternating irrigation cycle with a frequency of five days. Irrigation was only applied during day time at a constant rate of 2.6 cm d−1 (=11.7 cm3d−1), when plants transpired. Again, during the first five days, irrigation was uniform and the actual transpiration equalled the potential daily averaged transpiration of 10 cm3 (Fig. 9b). About three days after changing to partial irrigation, transpiration declined. Unlike simulations with constant transpiration demand, transpiration rate did not recover immediately after alternating irrigation but remained low for another day until it recovered again. Here, the transpiration reduction was slightly higher for the purely hydraulic case (H) due to parameterization of either hcrit or the parameters of Eq. 2a (see the section ‘Sensitivity’). Generally, transpiration was similar with and without additional chemical signalling, suggesting it could be described by a hydraulic signal alone.

Partial rootzone drying: a Cumulated actual daily transpiration for regulation as a function of hydraulic signalling (H) and of hydraulic + chemical signalling (H + C), Tpot remained at 10 cm3 d−1. Irrigation was either applied uniformly or partially at the right (R) or the left side of the compartment (L). Amount of irrigation per day remained constant. b Daily course of mass flux of chemical signal arriving at the leaves from each compartment and of the actual transpiration for H and H + C in comparison to potential transpiration (light grey). Irrigation was only applied during day (indicated by black bars)
Effects of an alternation event on the daily courses of transpiration and chemical signalling were examined in more detail (Fig. 9b). Initially, actual transpiration follows potential transpiration (light grey) and as maximum transpirational demand at midday approaches, both cases start to reduce their transpiration rate. Tact equals Tpot again in the ‘evening hours’. Thus, both cases reproduce the well-known midday depression in transpiration and its subsequent recovery. Before switching the irrigation, the drying side of the root system provides most of the chemical signal that decreases transpiration. This is maintained for 2 days after the irrigation was alternated, reflecting a lag phase of the system due to the time it takes to re-wet the previously dry soil, and dry out the previously irrigated rootzone.
Conclusions
We developed a model that simulates the effect of a root-produced chemical signal on stomatal regulation, transpiration and root water uptake. This model evaluated the impact of different regulatory mechanisms on plant transpiration when the root system is in a soil with spatial variations in soil water potentials. The model reproduced some experimental observations such as absolute xylem ABA concentrations, transient effects of root-produced chemical signals on transpiration, a transpiration restriction when previously dried roots re-commence water uptake, and transpiration oscillations (see Appendix). Those effects are attributed to transport limitations of the chemical signal from the roots that emerge when parts of the root zone dry out strongly so that there is no uptake and water flow from these regions. Transport limitation leads to a reduction of the stomatal regulation that may result in an increase in transpiration during a drying process. This phenomenon cannot be reproduced by classical root water uptake models that describe relative root water uptake (relative to potential transpiration) only as a function of rootzone water potential. Another transport related example is that of the flushing of chemical signals from a dry rootzone upon rewetting. This flush may lead to a temporal decrease in transpiration rate but may as well have other plant physiological effects. However, effects of chemical signals on modulating tissue hydraulic conductivity (e.g. via altering aquaporin expression in the leaves) that may enhance the impact of hydraulic signals on stomatal closure (Pantin et al. 2012) were ignored in this study.
Alternatively, chemical signalling seems less important over longer time scales, since models that predicted root water uptake based only on hydraulic signalling adequately simulated plant responses to drying soil. However, a detailed soil and root water uptake model shows that soil moisture heterogeneity influences plant hydraulics. Contrary to current assumptions, simulations indicate that effects of heterogeneous distribution of rootzone soil water potential and alternate partial rootzone drying on stomatal conductance can largely be explained by hydraulic signalling. This suggests that the spatial dynamics of root water uptake from soils with temporally varying spatial distributions of water content may be described by models that consider only the hydraulics of the soil-root system, even when the mechanism that relates transpiration to soil water potential is chemical signalling.
Furthermore, the model allows the effects of a simultaneously increasing rate of signal production and a decreasing water uptake rate on signal concentration in PRD plants to be quantified. While the mass fluxes from the dry side were about twice as high as those from the irrigated side, the concentration difference was three orders of magnitude. Therefore, measuring signal concentrations from roots in drying soil (Dodd et al. 2008a; b) may overestimate effects of chemical signalling on transpiration, as water (and signal) transport out of these roots decreases. Chemical signal fluxes cannot be linked directly to bulk nor equivalent soil water potentials (see also Eq. 8), as the heterogeneous distribution of soil and root water potential might initiate chemical signalling in only parts of the root system. To verify this model, experiments should simultaneously measure chemical signalling (concentrations and mass fluxes) as well as soil and root water potentials in both soil compartments of plants exposed to PRD.
The sensitivity analysis showed that the fitting parameters for hydraulic and chemical signalling sh and sc both influence the magnitude of transpiration reduction while the threshold pressure head for chemical signal production hL affects the onset of transpiration reduction. Although the relationship between soil water pressure and regulation of transpiration became non-linear for chemical signalling alone, further investigations are needed in relation to the distribution of soil moisture heterogeneity and to soil and root hydraulic properties.
In conjunction with experiments, the model might prove useful to further investigate the influence of soil properties (e.g. soil water holding capacity) on root water uptake patterns, chemical signalling and stomatal regulation. Being able to simulate chemical signalling between root and shoot may allow simulation of other physiological effects related to shoot growth and assimilate redistribution, that may be important for optimizing irrigation strategies such as PRD.
References
Barigah TS, Charrier O, Douris M, Bonhomme M, Herbette S, Améglio T, Fichot R, Brignolas F, Cochard H (2013) Water stress-induced xylem hydraulic failure is a causal factor of tree mortality in beech and poplar. Ann Bot 112:1431–1437. doi:10.1093/aob/mct204
Bravdo BA (2005) Physiological mechanisms involved in the production of non-hydraulic root signals by partial rootzone drying - A review. In: LE Williams (ed) Proceedings of the Seventh International Symposium on Grapevine Physiology and Biotechnology. International Society Horticultural Science, Leuven 1
Brodribb TJ, McAdam SAM (2011) Passive origins of stomatal control in vascular plants. Science 331:582–585. doi:10.1126/science.1197985
Buckley TN (2005) The control of stomata by water balance. New Phytol 168:275–291. doi:10.1111/j.1469-8137.2005.01543.x
Carsel RF, Parrish RS (1988) Developing joint probability-distributions of soil-water retention characteristics. Water Resour Res 24:755–769. doi:10.1029/WR024i005p00755
Christmann A, Weiler EW, Steudle E, Grill E (2007) A hydraulic signal in root-to-shoot signalling of water shortage. Plant J 52:167–174. doi:10.1111/j.1365-313X.2007.03234.x
Christmann A, Grill E, Huang J (2013) Hydraulic signals in long-distance signaling. Curr Opin Plant Biol 16:293–300. doi:10.1016/j.pbi.2013.02.011
Clausnitzer V, Hopmans JW (1994) Simultaneous modeling of transient three-dimensional root growth and soil water flow. Plant Soil 164:299–314
Couvreur V, Vanderborght J, Javaux M (2012) A simple three-dimensional macroscopic root water uptake model based on the hydraulic architecture approach. Hydrol Earth Syst Sci 16:2957–2971. doi:10.5194/hess-16-2957-2012
Damour G, Simonneau T, Cochard H, Urban L (2010) An overview of models of stomatal conductance at the leaf level. Plant Cell Environ 33:1419–1438. doi:10.1111/j.1365-3040.2010.02181.x
Davies WJ, Zhang JH (1991) Root signals and the regulation of growth and development of plants in drying soil. Annu Rev Plant Physiol Plant Mol Biol 42:55–76. doi:10.1146/annurev.arplant.42.1.55
Dodd IC (2009) Rhizosphere manipulations to maximize ‘crop per drop’ during deficit irrigation. J Exp Bot 60:2454–2459. doi:10.1093/jxb/erp192
Dodd IC, Theobald JC, Bacon MA, Davies WJ (2006) Alternation of wet and dry sides during partial rootzone drying irrigation alters root-to-shoot signalling of abscisic acid. Funct Plant Biol 33:1081–1089. doi:10.1071/fp06203
Dodd IC, Egea G, Davies WJ (2008a) Abscisic acid signalling when soil moisture is heterogeneous: decreased photoperiod sap flow from drying roots limits abscisic acid export to the shoots. Plant Cell Environ 31:1263–1274. doi:10.1111/j.1365-3040.2008.01831.x
Dodd IC, Egea G, Davies WJ (2008b) Accounting for sap flow from different parts of the root system improves the prediction of xylem ABA concentration in plants grown with heterogeneous soil moisture. J Exp Bot 59:4083–4093. doi:10.1093/jxb/ern246
Dodd IC, Theobald JC, Richer SK, Davies WJ (2009) Partial phenotypic reversion of ABA-deficient flacca tomato (Solanum lycopersicum) scions by a wild-type rootstock: normalising shoot ethylene relations promotes leaf area but does not diminish whole plant transpiration rate. J Exp Bot 60:4029–4039
Dodd IC, Egea G, Watts CW, Whalley WR (2010) Root water potential integrates discrete soil physical properties to influence ABA signalling during partial rootzone drying. J Exp Bot 61:3543–3551. doi:10.1093/jxb/erq195
Doussan C, Vercambre G, Pagès L (1998) Modelling of the hydraulic architecture of root systems: an integrated approach to water absorption—distribution of axial and radial conductances in maize. Ann Bot 81(2): 225–232
Doussan C, Pierret A, Garrigues E, Pages L (2006) Water uptake by plant roots: II - Modelling of water transfer in the soil root-system with explicit account of flow within the root system - Comparison with experiments. Plant Soil 283:99–117. doi:10.1007/s11104-004-7904-z
Dzikiti S, Verreynne JS, Stuckens J, Strever A, Verstraeten WW, Swennen R, Coppin P (2010) Determining the water status of satsuma mandarin trees citrus unshiu marcovitch using spectral indices and by combining hyperspectral and physiological data. Agric For Meteorol 150:369–379. doi:10.1016/j.agrformet.2009.12.005
Farquhar GD, Cowan IR (1974) Oscillations in stomatal conductance - influence of environmental gain. Plant Physiol 54:769–772. doi:10.1104/pp. 54.5.769
Feddes RA, Kowalik PJ, Zaradny H (1978) Simulation of field water use and crop yield. Pudoc Wageningen, The Netherlands
Franks PJ, Drake PL, Froend RH (2007) Anisohydric but isohydrodynamic: seasonally constant plant water potential gradient explained by a stomatal control mechanism incorporating variable plant hydraulic conductance. Plant Cell Environ 30:19–30. doi:10.1111/j.1365-3040.01600.x
Hartung W, Aboumandour AA (1980) Abscisic-acid in root cultures of phaseolus-coccineus l. Zeitschrift Fur Pflanzenphysiologie 97:265–269
Holbrook NM, Shashidhar VR, James RA, Munns R (2002) Stomatal control in tomato with ABA-deficient roots: response of grafted plants to soil drying. J Exp Bot 53:1503–1514. doi:10.1093/jexbot/53.373.1503
Javaux M, Schröder T, Vanderborght J, Vereecken H (2008) Use of a three-dimensional detailed modeling approach for predicting root water uptake. Vadose Zone J 7:1079–1079
Javaux M, Couvreur V, Vanderborght J, Vereecken H (2013) Root Water Uptake: From Three-Dimensional Biophysical Processes to Macroscopic Modeling Approaches. Vadose Zone Journal 12. doi:10.2136/vzj2013.02.0042
Kang S (2004) Controlled alternate partial root-zone irrigation: its physiological consequences and impact on water use efficiency. J Exp Bot 55:2437–2446
Khalil AAM, Grace J (1993) Does xylem Sap Aba control the stomatal behavior of water-stressed sycamore (acer-pseudoplatanus L) seedlings. J Exp Bot 44:1127–1134. doi:10.1093/jxb/44.7.1127
Li B, Feng Z, Xie M, Sun M, Zhao Y, Liang L, Liu G, Zhang J, Jia W (2011) Modulation of the root-sourced ABA signal along its way to the shoot in vitis riparia × vitis labrusca under water deficit. J Exp Bot 62:1731–1741. doi:10.1093/jxb/erq390
Liang JS, Zhang JH, Wong MH (1997) How do roots control xylem sap ABA concentration in response to soil drying? Plant Cell Physiol 38:10–16
Liu FL, Jensen CR, Andersen MN (2005) A review of drought adaptation in crop plants: changes in vegetative and reproductive physiology induced by ABA-based chemical signals. Aust J Agric Res 56:1245–1252. doi:10.1071/ar05062
Liu FL, Song R, Zhang XY, Shahnazari A, Andersen MN, Plauborg F, Jacobsen SE, Jensen CR (2008) Measurement and modelling of ABA signalling in potato (Solanum tuberosum L.) during partial root-zone drying. Environ Exp Bot 63:385–391. doi:10.1016/j.envexpbot.2007.11.015
Marenco RA, Siebke K, Farquhar GD, Ball MC (2006) Hydraulically based stomatal oscillations and stomatal patchiness in Gossypium hirsutum. Funct Plant Biol 33:1103–1113. doi:10.1071/fp06115
Martin-Vertedor AI, Dodd IC (2011) Root-to-shoot signalling when soil moisture is heterogeneous: increasing the proportion of root biomass in drying soil inhibits leaf growth and increases leaf abscisic acid concentration. Plant Cell Environ 34:1164–1175. doi:10.1111/j.1365-3040.2011.02315.x
Molz FJ (1981) Models of water transport in the soil-plant system - a review. Water Resour Res 17:1245–1260. doi:10.1029/WR017i005p01245
Pantin F, Simonneau T, Muller B (2012) Coming of leaf age: control of growth by hydraulics and metabolics during leaf ontogeny. New Phytol 196(2):349–366
Passioura JB, Tanner CB (1985) Oscillations in apparent hydraulic conductance of cotton plants. Aust J Plant Physiol 12:455–461
Puértolas J, Alcobendas R, Alarcón JJ, Dodd IC (2013) Long-distance abscisic acid signalling under different vertical soil moisture gradients depends on bulk root water potential and average soil water content in the root zone. Plant Cell Environ 36:1465–1475. doi:10.1111/pce.12076
Richards LA (1931) Capillary conduction of liquids through porous mediums. Phys-A J Gen Appl Phys 1:318–333. doi:10.1063/1.1745010
Romero P, Dodd IC, Martinez-Cutillas A (2012) Contrasting physiological effects of partial root zone drying in field-grown grapevine (Vitis vinifera L. cv. Monastrell) according to total soil water availability. J Exp Bot 63:4071–4083. doi:10.1093/jxb/ers088
Schroder T, Javaux M, Vanderborght J, Korfgen B, Vereecken H (2009) Implementation of a microscopic soil-root hydraulic conductivity drop function in a three-dimensional soil-root architecture water transfer model. Vadose Zone J 8:783–792. doi:10.2136/vzj2008.0116
Schroder N, Javaux M, Vanderborght J, Steffen B, Vereecken H (2012) Effect of Root Water and Solute Uptake on Apparent Soil Dispersivity: A Simulation Study. Vadose Zone Journal 11. doi:10.2136/vzj2012.0009
Schurr U, Gollan T, Schulze ED (1992) Stomatal response to drying soil in relation to changes in the xylem sap composition of helianthus-annuus.2. Stomatal sensitivity to abscisic-acid imported from the xylem sap. Plant Cell Environ 15:561–567. doi:10.1111/j.1365-3040.1992.tb01489.x
Simonneau T, Barrieu P, Tardieu F (1998) Accumulation rate of ABA in detached maize roots correlates with root water potential regardless of age and branching order. Plant Cell Environ 21:1113–1122. doi:10.1046/j.1365-3040.1998.00344.x
Somma F, Hopmans JW, Clausnitzer V (1998) Transient three-dimensional modeling of soil water and solute transport with simultaneous root growth, root water and nutrient uptake. Plant Soil 202:281–293. doi:10.1023/a:1004378602378
Stoll M, Loveys B, Dry P (2000) Hormonal changes induced by partial rootzone drying of irrigated grapevine. J Exp Bot 51:1627–1634. doi:10.1093/jexbot/51.350.1627
Taiz L, Zeiger E (2006) Plant physiology/Lincoln Taiz; Eduardo Zeiger. Sinauer, Sunderland, Mass
Tardieu F, Davies WJ (1993) Integration of hydraulic and chemical signaling in the control of stomatal conductance and water status of droughted plants. Plant Cell Environ 16:341–349. doi:10.1111/j.1365-3040.1993.tb00880.x
Tardieu F, Simonneau T (1998) Variability among species of stomatal control under fluctuating soil water status and evaporative demand: modelling isohydric and anisohydric behaviours. J Exp Bot 49:419–432. doi:10.1093/jexbot/49.suppl_1.419
Van Genuchten MT (1980) A closed form equation for predicting the hydraulic conductivity of unsaturated soils. Soil Sci Soc Am J 44:892–898
Wan XC, Zwiazek JJ (2001) Root water flow and leaf stomatal conductance in aspen (Populus tremuloides) seedlings treated with abscisic acid. Planta 213:741–747. doi:10.1007/s004250100547
Wilkinson S, Davies WJ (1997) Xylem sap pH increase: a drought signal received at the apoplastic face of the guard cell that involves the suppression of saturable abscisic acid uptake by the epidermal symplast. Plant Physiol 113:559–573
Wilson JB (1988) A review of evidence on the control of shoot: root ratio, in relation to models. Ann Bot 61:433–449
Wolf O, Jeschke WD, Hartung W (1990) Long-distance transport of abscisic-acid in nacl-treated intact plants of lupinus-albus. J Exp Bot 41:593–600. doi:10.1093/jxb/41.5.593
Yin X, van Laar HH (2005) Crop systems dynamics : an ecophysiological simulation model for genotype-by-environment interactions/Xinyou Yin; H. H. van Laar. Academic Publishers, Wageningen
Acknowledgments
This work is a contribution of the Transregio Collaborative Research Centre 32. Patterns in Soil-Vegetation-Atmosphere Systems: Monitoring, Modelling and Data Assimilation, which is funded by the German research association, DFG. ICD thanks the EU project SIRRIMED (FP7- KBBE-2009-3-245159) for continued support of work on PRD.
Author information
Authors and Affiliations
Corresponding author
Additional information
Responsible Editor: Rafael S. Oliveira.
Appendix
Appendix
Influence of shoot volume (buffer size) on transpiration reduction
When shoot volume was smaller than 0.5*VRoot, multi-frequency oscillations in simulated transpiration occurred (Fig. 10) for all scenarios which included chemical signalling. When averaged over time, the trend of transpiration and the signal concentrations (data not shown), were independent of buffer size. Oscillations in stomatal conductance have been observed in several plant species (Buckley 2005), including cotton (with frequencies ranging from 1.3 to 1.8 h−1) (Farquhar and Cowan 1974; Marenco et al. 2006; Passioura and Tanner 1985) and citrus trees when exposed to high vapour pressure deficits (mandarin tree, frequency ca. 1.8 h−1) (Dzikiti et al. 2010). Regardless of the physiological explanation (and site) of this buffer, simulations revealed that, stomatal conductance can oscillate due to a chemical signal arriving from the roots.

Influence of buffer size: Oscillations in transpiration rates are related to buffer size. A buffer size of 1*VRoot corresponds to an average residence time of 0.1 days and a root:shoot ratio of 1
Rights and permissions
About this article

Cite this article
Huber, K., Vanderborght, J., Javaux, M. et al. Modelling the impact of heterogeneous rootzone water distribution on the regulation of transpiration by hormone transport and/or hydraulic pressures. Plant Soil 384, 93–112 (2014). https://doi.org/10.1007/s11104-014-2188-4
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11104-014-2188-4