
Abstract
Aims
In serpentinitic areas non-endemic plants suffer from the serpentine syndrome, due to high Ni and Mg concentrations, low nutrients and Ca/Mg ratio. We evaluated the environment-soil-vegetation relationships in a xeric inner-alpine area (NW Italy), where the inhibited pedogenesis should enhance parent material influences on vegetation.
Methods
Site conditions, topsoil properties, plant associations and species on and off serpentinite were statistically associated (51 sites).
Results
Serpentine soils had higher Mg and Ni concentrations, but did not differ from non-serpentine ones in nutrient contents. The 15 vegetation clusters often showed substrate specificity. Two components of the Canonical Analysis of Principal Coordinates, respectively related to Mg and to Ni and heat load, identified serpentine vegetation. Random Forests showed that several species were positively correlated with Ni and/or Ca/Mg or Mg, some were negatively associated with high Ni, Mg excess affected only few species. Considering only serpentine sites, nutrients and microclimate were most important.
Conclusions
Ni excess most often precludes the presence of plant species on serpentinite, while an exclusion due to Mg is rarer. Endemic species are mostly adapted to both factors. Nutrient scarcity was not specific of serpentine soils in the considered environment. Considering only serpentine sites, nutrient and microclimatic gradients drove vegetation variability.
Similar content being viewed by others

Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
Serpentinite is a metamorphic rock mainly composed of Mg silicates (serpentine minerals, with accessory chlorites, talc, and sometimes olivine and pyroxenes), and often includes magnetites and chromites. Serpentinite weathering originates soils (normally called “serpentine soils”) typically characterized by chemical and physical properties that reduce plant productivity and create stress and toxicity to non-adapted species, the so called “serpentine syndrome” (Jenny 1980). Several factors are thought to be responsible of the serpentine syndrome (Alexander et al. 2007), such as a low Ca:Mg ratio caused by the high amounts of Mg released from the parent material and abundant heavy metals (Ni, Cr, Co). In addition, soils often have low macronutrient (N, P, K) concentrations both because of their paucity in the rock and of the presence of sparse vegetation. Susceptibility to drought and erosion often characterizes serpentine soils too, because of dark colour, coarse soil texture, rockiness and shallow soil depth (Brooks 1987).
Although vegetation growing on these soils is typically sparse and stunted, plant diversity is often high with abundant endemic taxa (e.g. Kruckeberg 1984). Some of these species hyperaccumulate metals (commonly Nickel) and Ni concentrations in their tissues may be more than 1,000 mg kg-1 on the dry weight (Van der Ent et al. 2013). Many serpentine endemic species in Europe are known Ni hyperaccumulators (e.g. Brooks and Radford 1978) and often they have a narrow geographical distribution, such as Alyssum argenteum (Cecchi et al. 2010), which is a perennial Brassicaceae endemic to serpentine rocky outcrops in the Western Alps. It grows at low altitude on south-facing slopes, from Val Chisone in the south to Aosta Valley in the north (Pignatti 1992).
The role of the factors involved in the distribution and competitive capacity of serpentinite endemic plants, and of metal hyperaccumulators in particular, has always attracted the curiosity of ecologists, biologists and soil scientists (Roberts and Proctor 1992). The relative importance of each factor of the serpentine syndrome greatly varies depending on local climate, plant community (Proctor and Nagy 1991) and single species (Lazarus et al. 2011). In particular, the role of heavy metals in influencing serpentine vegetation is unclear. Brady et al. (2005) in their review reported that the serpentine syndrome is mostly associated with low Ca/Mg ratio and with high Mg contents; adaptative traits to high Ni (e.g., hyperaccumulation or, more often, exclusion) are rarer phenomena, evidencing low Ni ecological impacts. Some works pointed out that tolerance to high Ni is readily achieved, reducing its ecological effect on plant communities (e.g., Proctor 1997). Conversely, other authors have reported that Ni has strong negative effects on plant cover (Lee 1992; Chardot et al. 2007) or on biodiversity (Batianoff and Singh 2001), while it is positively correlated with the number of endemic taxa (Batianoff and Singh 2001). Typically, the works on serpentine ecology explore the edaphic gradients between dry and nutrient-poor barrens and more favourable forested sites (e.g. Chiarucci 2004; Carter et al. 1987), or the environmental gradients causing vegetation differences within serpentine habitats (Tsiripidis et al. 2010). Common results are that soils in open rocky outcrops or taluses (i.e. serpentine barrens) are nutrient-poor (Chiarucci 2004) and are characterized by a high Mg/Ca molar ratio (e.g. Carter et al. 1987). As a demonstration of low Ni impact on plant life, Ni availability is often higher under the better developed and structured communities (Chiarucci 2004; Tsiripidis et al. 2010) because of enhanced acidification under forest vegetation.
Few works, instead, consider edaphic gradients and their effects on vegetation patterns on serpentinite and on nearby, analogous non-serpentinite habitats (i.e. comparing barren or forested sites on and off serpentinite). In some of these cases, such as in humid, high-altitude subalpine (boreal) forest and alpine habitats, serpentine endemic species or plant communities were well correlated with high levels of available Ni, while the Ca/Mg ratio or the nutrient contents did not explain vegetation variability caused by substrate: different nutrient contents and element cycling characterized differently developed soils, both on and off serpentine (D’Amico and Previtali 2012).
Element mobility through the profile is high in humid environments, because of both leaching and biocycling; in particular Ca biocycling is enhanced in serpentine-rich soils (Bonifacio et al. 2013), hence effectively decreasing the effect of the Ca/Mg ratio on vegetation. The effect of soil development is thus superimposed on that of the parent material in humid areas, and the final soil characteristics may diverge from those of poorly weathered and poorly leached soils. On the contrary, xeric climates inhibit element leaching from soils, which are typically less developed, thus the ecological effect of the edaphic components of the serpentine syndrome on vegetation should be enhanced, allowing a better evaluation of the soil-vegetation relationships.
Our hypothesis was that soil development and the consequent divergence of chemical properties from those of the parent material may mask the relationships between soil and vegetation, particularly when highly mobile nutrients or metals with high affinity to organic matter are concerned. The purpose of this study was thus to investigate soil-vegetation relationships on serpentinite in xeric, low altitude habitats in the Western Alps, where element leaching and pedogenesis should be inhibited. To reach this aim, we selected soils from a xeric inner alpine area, both on serpentine and non-serpentine parent materials, evaluated that all soils have a comparable poor development degree and then assessed the relationships between vegetation and those soil properties which are considered the most important in the serpentine syndrome, such as macronutrient availability, Ca:Mg molar ratio, heavy metal availability and rockiness, making use of both well-known statistical tools and recently proposed methods for data treatment. The statistical approach can help to disentangle the respective importance of different factors in soil-vegetation relationships thanks to permutation methods, although direct cause-effect relationships can only be evaluated by lab or field experiments.
Materials and methods
Study area
The central part of the Aosta Valley (Regione Autonoma Valle d’Aosta), characterized by yearly average precipitation below 750 mm, was considered in our study. This region is located in the north-western Italian Alps, close to the French (in the west) and the Swiss border (in the north) (Fig. 1). The climate is primarily related to topography: the central part of the region is surrounded by high mountain ranges and it is characterized by rain-shadow effects in every season giving rise to continental, xeric, inner-alpine climate (Mercalli 2003). In the considered xeric area, winter temperatures are between 2° and −5 °C, with the lowest values in the valley floors (because of thermal inversions) and at high altitude, while summer temperatures are between 21 °C and 10 °C (decreasing with altitude). In the lowest parts of south-facing slopes the average temperature is always above 1 °C. The average rainfall is below 600–700 mm y−1, and can be as low as 485 mm y−1 in the Aosta area. Spring and autumn months have the highest rainfall amounts, while winter and summer minima are typical; the average rainfall of July is below 40 mm (Mercalli 2003). Water scarcity is a strong ecological constraint, particularly on south-facing slopes, where steppe vegetation (dominated by Bromus erectus, Festuca valesiaca, Stipa pennata and Teucrium chamaedrys) with only scattered trees (Quercus pubescens, Pinus sylvestris and Castanea sativa) is the natural vegetation type at the montane (sub-boreal) phytoclimatic level. Dense Castanea sativa, Pinus sylvestris and Quercus pubescens forests cover the cooler north-facing slopes at the same altitude. Thus, a xericity gradient can be observed within the xeric area. In the driest part of the region, steppe vegetation is found up to 2,400 m a.s.l., where steppe species mingle with alpine ones, adapted to low temperatures and short growing seasons.

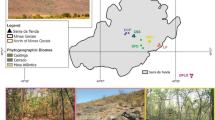
The study area in the central part of the Valle d’Aosta region. The area characterized by inner-alpine xeric climate is roughly marked by a dotted line; soil and vegetation sampling sites are represented by triangles (several overlaps due to scale); serpentine outcrops are evidenced by shaded areas
Several rock complexes are found in the Aosta Valley: serpentinite, mafic rocks and calcschists, included in the Piedmontese Ophiolite Complex, are common in the eastern sector, while the western part is dominated by calcschists and gneiss. Pleistocene glaciers covered large portions of the area until 12,000–15,000 years BP, and mixed glacial till, with a calcareous matrix, is widespread.
Field data collection, soil sampling and analysis
Fifty one sites (soil profiles associated with vegetation surveys) were selected according to the average annual rainfall (below 750 mm), among 384 previously observed sites scattered around the whole region, representative of the typical inner-alpine environments. In each site, plant species (identified according to Pignatti 1992) were recorded in homogeneous square areas of 16 m2, visually estimating the cover (%) of each species.
At each site, the following data were also collected: altitude, slope steepness, aspect, surface rockiness, bare soil, and tree cover (calculated as % area on a 100 m2 surface). Surface rockiness, bare soil and tree cover were determined by visual area estimation. Aspect and slope steepness were combined into the heat load factor, a proxy of potential solar radiation and potential evapotranspiration (McCune and Leon 2002). Rainfall data were collected from 12 regional weather stations located close to the sampling sites and the average annual rainfall was included among environmental parameters.
Soil pits were dug at each site, down to the C or R horizon (parent material) and the soil profile was examined to assess soil development and pedogenic processes. The soils found in the inner-alpine, xeric area were usually weakly developed Regosols, Leptosols or, rarely, slightly more evolved Cambisols, Phaeozems and Calcisols (IUSS Working Group 2006). Samples were collected from all genetic horizons, but only A horizons were considered in this study.
The soil samples were air dried, sieved to 2 mm and analyzed according to the USDA methods (Soil Survey Staff 2004). The pH was determined potentiometrically in water extracts (1:2.5 w/w). Exchangeable Ca, Mg, K and Ni (Caex, Mgex, Kex, Niex) were determined after exchange with NH4-acetate at pH 7.0. The acid-extractable element concentrations (CaT, MgT, NiT) were determined after HCl-HNO3 hot acid digestion). In all extracts, the elements were analysed by Atomic Absorption Spectrophotometry (AAS, Perkin Elmer, Analyst 400, Waltham, MA, USA). The total C and N concentrations were evaluated by dry combustion with an elemental analyser (CE Instruments NA2100, Rodano, Italy). The carbonate content was measured by volumetric analysis of the carbon dioxide liberated by a 6 M HCl solution. The Organic Carbon (OC) was then calculated as the difference between total C measured by dry combustion and carbonate-C. Available P (POlsen) was determined by extraction with NaHCO3 with P detection by molybdate colorimetry.
Rock fragments >5 mm were cleaned with sodium hexametaphosphate, sorted according to the lithology and weighted to semi-quantitatively characterize the soil parent material. The frequency distribution of serpentinite clasts in the soils was clearly bimodal: no serpentinite clasts, 21 samples; 10 %, 20 %, 30 % and 50 %, 1 sample each; 60 %, 3 samples; 70 % 1 sample; 80 %, 2 samples; 90 %, 3 samples and 100 %, 17 samples.
Data analysis
Statistical analysis were carried out using either SPSS for Windows version 17.0 or R 2.15.1 software (R Foundation for Statistical Software, Institute for Statistics and Mathematics, Vienna, Austria).
Based on the bimodal distribution of serpentine content in the parent material, we decided to split soil profiles into two groups depending on whether the parent material was dominated by serpentinites (>= 60 % of serpentinitic clasts by weight) or not. To verify whether clear thresholds existed or if a chemical/ecological gradient was instead present, we also performed additional analyses considering different abundance of serpentine coarse fragments. The differences in soil properties were evaluated by a one-way analysis of variance (ANOVA), using lithology as independent variable. The homogeneity of variance was checked by the Levene test and the variables showing significant differences ( i.e. CaCO3, MgT, CaT/MgT, NiT, Caex/Mgex, Mgex, Caex/Mgex, Niex, POlsen) were log-natural transformed for analysis. The correlation between variables was evaluated using the Pearson’s coefficient (two-tailed), after a visual inspection of the data to verify that the dependence relationship was linear. In case of non linearity, Spearmann’s correlation coefficient was instead used.
The numerical elaborations regarding vegetation and soil-vegetation relationships were performed excluding tree species, which are mostly correlated with climatic and pedoclimatic site properties.
Vegetation types were classified using Cluster Analysis (CA), selecting average linkage as agglomeration criteria owing to its high cophenetic correlation value. The best dissimilarity algorithm (Bray-Curtis) was selected according to the function rankindex in the Vegan package (Oksanen et al. 2011), which correlates the species data with a given gradient (in this case, soil-environmental properties) using many dissimilarity algorithms. To facilitate the ecological interpretation of the clusters, common indicator species (Legendre and Legendre 1998) for each cluster were obtained with the help of the indval function, included in the labdsv package. Cluster stability was assessed through the “bootstrap” noise-adding and subsetting methods (Hennig 2007): if the resulting Clusterwise Jaccard mean is below 0.5, the cluster is considered “dissolved” and not significant, while it is regarded as “stable” and significant if the value is above 0.75. The number of clusters to be considered during the following analysis was chosen based on the ratio between the total number of clusters and the number of stable ones and according to their ecological significance. The bootstrap method was applied to a variable number of clusters (2–18).
A correlation analysis was performed on the soil-environmental properties, to detect collinearities (R2 above 0.75) and select a subset of independent variables to be used in the following elaborations. The selected variables were altitude, surface rockiness, bare soil, N, C/N ratio, available P, molar Caex/Mgex ratio, Mgex, Kex, Niex, CaCO3, tree cover, heat load and average yearly rainfall. Caex, pH and C were omitted as strongly collinear with, respectively, CaCO3 and Ca/Mg, and N. Altitude, N, P, Kex, Caex, Mgex and Niex were log-transformed prior to analysis.
We used constrained analysis of principal coordinates (CAP, function capscale, included in the vegan R-package), based on the Bray–Curtis distance, to determine the most influential environmental variables involved in the plant community composition (Anderson and Willis 2003). This multivariate technique offers an appropriate way to apply canonical constraints using a flexible choice of dissimilarity measures. The significance tests for each variable were computed using the marginal testing method included. In order to detect the ranking of importance of pedo-environmental factors involved in soil-vegetation relationships, we applied a stepwise analysis on the CAP (function ordistep), which shows how much the model is improved when each variable is added, with random permutations.
We used the Random Forests (RF) (Breiman 2001), included in the RandomForest R library (Liaw and Wiener 2002), to detect the important factors involved in the presence/absence of the species growing in more than 10 % of the study sites. RF is an improvement of the Classification Tree method (CART), which shows the optimal distribution ranges of plant species (Vayssiéres et al. 2000). RF is a more robust method than most normally used in ecological niche modelling (Evans and Cushman 2009), and does not need further accuracy estimates (Cutler et al. 2007). The importance of predictive (soil-environmental) variables is estimated by looking at how much the misclassification error (the out of bag error) increases when each variable is permuted while the others are left unchanged. The increase of the error is proportional to the importance of each predictive variable. After checking the optimal number of trees (ntree) reducing the out of bag error to a minimum, we modified the ntree in the RF from 500 to 3,000. The optimal number of randomly selected variables (mtry) to be used in each step of the bootstrap process was also checked for each species.
Positive or negative interactions between predictive variables and plant species were checked using generalized linear models (Guisan et al. 1998, glm function, family binomial), using only the important variables for each considered species.
In order to check if soil-vegetation patterns were substrate-specific or generic for the whole xeric area in Valle d’Aosta, we performed the analyses described above also on the subset of sites with a serpentinite content in the parent material ≥60 %. This subset was called “serpentine-dominated soils” through the text.
Results
Soils
The A horizons of both serpentine-dominated and non-serpentine soils showed a wide pH range (from 4.4–4.8 to above 8, Table 1) and often had carbonates. Calcium carbonate enrichment in surface soils was common, up to 160 g kg−1, and precipitation of secondary carbonates was often observed in subsurface horizons, mainly in soils derived from non-serpentine parent materials but sometimes also on serpentinic ones. The elemental composition was well related with the parent material: serpentine-dominated soils had significantly higher MgT and NiT, and a significantly lower CaT/MgT ratio (Table 1) than soils formed on other rock types. CaT and the CaT/MgT ratio were, however, relatively high and values up to 1.5 were found. The specificity of the parent material was retained also in the available pools of elements: Niex and Mgex were significantly higher on serpentinite, while Caex did not differ (Table 2). The molar Caex/Mgex ratio was often above 1 also on serpentinite, but it was still significantly lower than on other substrates. While the exchangeable contents of both Mg and Ni were significantly correlated to their total concentration (rP = 0.415 and 0.778, respectively, p < 0.01), Caex and CaT were not (p = 0.158). Tree cover was uncorrelated with all substrate-related edaphic factors and with parent material lithologies.
As expected, all significant differences were also present when a higher threshold was chosen for serpentine-dominated soils and significant differences (p < 0.01) in carbonate contents and pH appeared (lower on serpentinitic soils) when almost pure serpentinite was selected as a threshold (e.g. ≥80 % serpentinitic clasts).
Both Niex and NiT were well correlated with the other typical properties of serpentinitic soil, i.e. MgT (r = 0.862 and 0.656 in the case of total and exchangeable Ni contents, respectively, p < 0.01), Mgex (r = 0.678 and 0.416, p < 0.01), the total Ca to Mg ratio (r = −0.477 and r = −0.398, p < 0.01), as well as the Caex/Mgex ratio (r = −0.488 and −0.398, p < 0.01). No correlation was instead present between Ni and Ca. When the statistical analysis was performed only on the serpentine-dominated soils subset (≥60 % serpentine clasts), all significant correlation were retained in the case of NiT, while they became less significant (p < 0.05) and the one with Mgex disappeared in the case of Niex. The concentrations of organic C, N, P and K were not related to the parent material and better linked to land use. Some grassland soils were in fact particularly rich in nutrients, while forest habitats influenced the C/N ratio. Available P was dependent on the amounts of organic matter (rP = 0.472, p < 0.01), and negatively associated with bare soil (rP = −0.296, p < 0.05) and heat load (rP = −0.529, p < 0.01). These two variables were also correlated to each other (rP = 0.307, p < 0.05).
Considering only serpentine dominated soils, rainfall negatively influenced pH values (rP = −0.751, p < 0.01) and CaCO3, while it was positively associated with the C/N ratio (rP = 0.591, p < 0.01). The C/N ratio was no more related with tree cover and organic carbon was significantly correlated with all nutrients and exchangeable elements (Caex, Mgex, N, K and P). Niex was not correlated with any soil or environmental property. All correlations were retained when higher thresholds for serpentine-dominated soils were selected. A large variance of chemical properties characterized therefore also pure serpentinite soils.
Vegetation
Plant communities were grouped into 15 ecologically meaningful clusters (Fig. 2a). The full list of species associated with the vegetation clusters, and their indicator values are reported in the electronic annex. The Jaccard index was often slightly below 0.75 (electronic annex 1), probably because of the presence of some widespread species such as Teucrium chamaedrys and Bromus erectus. The first hierarchical cluster subdivisions identified clusters mainly characterised by the dominance of serpentinites (single-site clusters 1, 11, 9 and a group including small serpentine clusters 2, 3, 5 and non-serpentine clusters 12 and 13) and a group of mixed large clusters.

Cluster dendrograms of the total vegetation relevèes (a) and of serpentine-dominated sites (b), (hierarchical clustering, based on Bray-Curtis distance algorithm and on average linkage method). The first number in the parenthesis indicates the number of sites in each cluster, the second one the number of serpentine-dominated clusters
Xerophilous prairies or steppe formations, with scarce or absent tree or shrub cover (Fig. 3a), mostly on southward slopes (Fig. 3b) at different altitudes (Fig. 3c), were represented by cluster 14 and cluster 8 (differentiated by a higher shrub cover, mainly Juniperus communis, and by past agricultural activities, such as terraces or surface rock removal in cluster 8). These formations occurred both on serpentinite and on non-serpentinite parent material (Fig. 3d). Quite similar steppic grasslands, on stable serpentinite talus slopes at different altitudes (Fig. 3c and d), were included in cluster 2. Cluster 10 consisted of xerophilous understory communities of montane (sub-boreal) Pinus sylvestris forests (Fig. 3a). The most important indicator species of this type of community was Carex humilis (electronic annex 1), but many steppe or prairie species grew despite the weak solar radiation. Understory communities of similar forest habitats were also represented by cluster 3, on serpentinite (Fig. 3d), often on northward aspects; ferns were characteristic and some were serpentine endemics (i.e., Asplenium cuneifolium). Cluster 4 included the understory vegetation of low altitude Quercus pubescens, Pinus sylvestris and Castanea sativa open forests, with high heat load (Fig. 3b) and low tree cover, growing both on calcschists and serpentinite (included in the same cluster even if clearly differentiated by the presence of Alyssum argenteum on serpentinite). Rocky outcrops were included in cluster 5 (on serpentinite, characterized by A. argenteum), cluster 7 (never on serpentinite), and 13, on different substrates, at higher altitudes (Fig. 3). Caryophyllaceae often characterized serpentine vegetation, particularly on rocky outcrops or talus slopes (e.g., Minuartia laricifolia and Cerastium arvense). Among the endemic ultramafic species, the hyperaccumulator Alyssum argenteum was common in low altitude open forests and serpentinite rocky outcrops, but it was also found in montane Pinus sylvestris forests and open steppe communities (electronic annex 1).

Tree cover (%), heat load, altitude (m) and serpentinite content (%) in the soil parent material characterizing the 15 vegetation clusters
Considering only serpentine sites (Fig. 2b), 9 clusters were obtained: 4 single-site clusters, a large one (11 sites, including common xerophilous serpentine communities (the main indicator species was Alyssum argenteum), another dominated by thick Pinus sylvestris forests (4 sites) devoid of endemic species, and other smaller ones separating respectively scree slopes, thick Scots pine forests with ferns, rocky outcrops. No cluster separated pure serpentine sites from those having between 60 % and 90 % serpentinite in the soil coarse fragment fraction.
Soil-vegetation relationships: serpentine and non-serpentine sites
The CAP analysis evidenced that plant communities were well associated with both environmental and soil properties. CAP1 was negatively associated with Mgex, CAP2 positively with tree cover and rainfall, CAP3 increased with high available Ni and low heat load (Table 3). If CAP2 identified forest habitats, CAP4 was negatively correlated to rainfall and C/N ratio, separating therefore the most steppic grasslands from the most humid forests. The least fertile sites, with low Ca/Mg ratio and available P, were associated to the highest values of CAP5. The space identified by CAP1 and CAP3 well described the clusters previously found and allowed to separate serpentine adapted associations from those that were not present on this parent material (Fig. 4). In fact, many sites of cluster 4 were well associated with negative CAP1 values (high Mg levels). Similar associations with high Mg levels were observed for most sites belonging to clusters 5, 9 and 15. On the contrary, clusters 1, 8, 10 and 13 appeared mostly negatively associated with soil Mg. The correlation of plant communities with soil Ni was masked by the opposite correlation with heat load: sites apparently well correlated with high or low soil Ni actually were often characterized by, respectively, low or high heat load values. Clusters 1, 5, 10, 13 were mostly associated with high values of CAP3 (high Ni and/or low HL), while clusters 14, 6 and 9 were mostly associated with low values of this axis. The space identified by positive values of CAP1 and negative CAP3 values comprised only non-serpentine sites. The stepwise analysis indicated that the CAPSCALE performance was strongly dependent on Ni, Ca/Mg ratio, elevation, heat load and tree cover, which were thus the most important variables associated with vegetation gradients in the study area (Table 3).

CAP scatterplots of xerophilous vegetation; the combination of axis 1 (CAP1) and 3 (CAP3) is shown, as most representative of serpentine habitats. Sites are represented by cluster numbers and serpentinite-dominated sites are indicated by a black dot. The following abbreviations have been used: SR surface rockiness, ALT altitude, HL heat load, BS bare soil
The factors associated with the presence of common species, as well as the separate ecological effect of heat load and Ni, could be evidenced by the RF analysis. Taking into account the soil properties that allowed to distinguish between fertility conditions of serpentine and non-serpentine soils, i.e. Niex, Mgex and Caex/Mgex (Table 2), the RF rankings (Table 4) showed that some species were well associated to serpentine conditions, such as high Niex, low Caex/Mgex or high Mgex values, while some others were negatively correlated to substrate-derived soil properties. In particular, Alyssum argenteum, Alyssoides utriculata and Cerastium arvense were very well correlated with both high Ni and Mg contents or Ca/Mg ratio. A good association with serpentine soils was shown also by Biscutella laevigata, well correlated with high Niex and low values of Caex, while a good association with high Niex levels without any effect of exchangeable basic cations was observed for Minuartia laricifolia and Sedum montanum (well correlated also to elevated Caex concentrations). Some species were correlated to high Mgex levels and/or low Caex/Mgex ratios without any relationships with Ni contents: Armeria arenaria, Anthyllis vulneraria s. l. and Stachys recta .
Negative association with high Niex contents was observed on many species: Achillea millefolium, Alyssum alyssoides, Berberis vulgaris, Carex halleriana, Festuca valesiaca, Fumana ericoides, Helianthemum nummularium and Koeleria vallesiana. High Mgex contents were negatively correlated only with Euphorbia cyparissias and Teucrium chamaedrys. The explanatory power of all the considered edaphic and environmental variables was extremely low for some common species, such as Dianthus carthusianorum, Dianthus sylvaticus, Potentilla neumanniana (negative correlation with high Niex), Carex humilis and Sempervivum arachnoideum (negative association with high Mgex). Available P never helped in explaining species distribution and the effect of Kex was also limited: Anthyllis vulneraria was indeed the only species markedly associated to the K-richest sites.
Soil-vegetation relationships: serpentine sites
The CAP analysis evidenced that serpentine plant communities were strongly dependent on climatic factors (rainfall and heat load), which influenced also the few “important” factors retained after the variable selection based on intercorrelation and variance inflating factors (Table 5). In particular, CAP1 identified a Mg gradient associated with an inverse rainfall one, CAP2 represented a CaCO3 - heat load gradient, CAP3 was well correlated with negative heat load and with positive CaCO3, P and C/N, making it not easily interpretable; CAP4 was influenced by high P and Mgex, CAP5 by C/N and Mg, CAP6 by rainfall. Niex was excluded as its influence on the overall model fitting was negligible.
RF rankings (Table 6) for presence-absence species data showed a generalized, strong decrease in Niex importance compared with the complete data set: only Alyssoides utriculata and Cerastium arvense kept their positive correlation with high Niex, while no species were negatively associated with it. Mgex was a rather important factor in explaining the presence/absence of many species: the positive correlation obtained when analyzing the whole data set were retained also in this reduced set (Alyssum argenteum, Cerastium arvense and Stachys recta), while a positive association with Helianthemum nummularium and Berberis vulgaris and a negative one with Minuartia laricifolia appeared. The negative associations between Caex and Alyssum argenteum and Biscutella laevigata were retained. On serpentine soils, Festuca valesiaca, Galium lucidum, Helianthemum nummularium, Teucrium chamaedrys had negative relationships with Caex; the first two species, in particular, had positive correlations with Caex on the complete data set. P became an important factor for many species growing on serpentine soils; this nutrient was never important when considering the complete data set.
Discussion
Pedogenesis in xeric soils in mountain habitats
In this work we wanted to evaluate soil-plant relationships in serpentinic areas, taking into consideration the effect that soil development has on element concentration and, more generally, on soil chemical properties. We therefore maximized the possibilities to detect differences by selecting a xeric environment, where no marked element movements should have occurred. Both the lack of carbonate dissolution in A horizons and CaCO3 precipitation in deeper ones confirmed the pedogenic trends of xeric environments. The high acidity observed in the surface horizons of some serpentinitic soils is common also in other dry environments, such as Mediterranean Italy (Bonifacio et al. 1997) and California (e.g. Lee et al. 2004) and in the study area it was related to the presence of organic matter (rS = −0.591, p < 0.01, n = 20, i.e. excluding the soils where pH was buffered by carbonates), thus more linked to the acidifying capacity of organic matter than to leaching. The differences between parent materials were well visible in the total element concentrations, and also partially retained in the available forms. The MgT and NiT concentrations were high on serpentinite, although with a very high variance, but CaT and the CaT/MgT ratio were much higher than normal in ultramafic soils: in fact, the CaT/MgT ratio usually varies between 0.01 and 0.1 (Brooks 1987). The high ratio we found in the study area may be related to the presence of carbonate inclusions or to aeolian additions. Aeolian inputs in the Alps are common and arise both from Saharan dusts (Goudie and Middleton 2001) and from shorter range intra-alpine sources (Küfmann 2002). However, the lack of correlation between CaT and Caex suggests a decoupling of Ca availability from direct weathering of the parent material, and indicates that processes affecting only available forms, such as biocycling, likely played an important role. Calcium indeed is one of the elements that undergo important nutrient uplift and biocycling (Blum et al. 2008), which are particularly enhanced on serpentinitic soils due to element deficiency (Bonifacio et al. 2013). Biocycling may thus contribute to increase the Ca available pool in surface horizons, eliminating the differences between parent materials and decreasing therefore the effects that Ca deficiencies have on vegetation. Serpentinitic soils were characterized by much higher available Ni concentrations than non-serpentinitic ones, even at high pH values and in the presence of carbonates. High Niex contents were observed elsewhere in weakly developed soils, and were related to the incipient weathering of Ni-rich primary minerals (Carter et al. 1987). The Niex contents and the lack of correlation between Niex and pH values were similar to those found in extremely acidic subalpine or weakly developed alpine soils on serpentinite in nearby valleys (D’Amico and Previtali 2012), where it was leached more efficiently from the most acidic soils; in humid subalpine and alpine serpentine soils, Niex was well correlated with organic carbon, while in these xeric soils Niex and C were not associated, suggesting that the metal affinity to organic matter is not sufficient to fully explain Ni availability.
The concentration of organic carbon and of N, P and K, whose availability in forest soils is mainly linked to the turnover of organic matter (Lal et al. 2007), was not related to the parent material. Therefore, nutrient scarcity is not a specific feature of serpentine soils in Alpine environments. The negative correlations between available P and bare soil or heat load suggested that nutrient scarcity was more linked to the limitation to vegetation productivity of the hottest and driest areas, where drought stress associated with topographic position is frequent. The correlations between P, bare soil and heat load were retained also amidst serpentine habitats, confirming the poor primary productivity and the slow nutrient biocycling in the most xeric habitats. Vegetation development on serpentinitic outcrops in Tuscany (Chiarucci 2004) was also inhibited by topographic positions, causing heat and drought stress. In xeric environments of the western Alps, however, this drought stress was not dependent on the substrate lithology and thus could not be considered part of the serpentine syndrome.
The effect of serpentinite soils on vegetation: different approaches, different results
The effect that parent material has on vegetation development was well depicted by cluster analysis, although mainly the smallest clusters showed substrate specificity, while most of the largest ones embraced sites on different substrates. In most cases, single site and small clusters identified serpentine plant communities, pointing to a marked heterogeneity among serpentine vegetation. Such high local diversification of serpentine plant communities is common to other serpentine habitats, such as dry areas in the American Pacific Northwest (del Moral 1972) or dry subtropical South African mountains (Reddy et al. 2009). Some of these small clusters included serpentine-endemic species, such as Aethionema thomasianum, Notholaena marantae and Asplenium cuneifolium, or serpentine-adapted species (Silene vulgaris with peculiar morphological characteristics including small stature, thickened and purple leaves, which may represent a unique serpentine ecotype, as described in D’Amico and Previtali 2012).
The Canonical Analysis of Principal Coordinates allowed to better evidence the relationships between site and soil characteristics and vegetation clusters, although the proportion of inertia explained by the model was rather low (28.02 %). This was probably due to the typical disorder of ecological systems hypothesized by other authors (e.g. Chiarucci et al. 2001 and Tsiripidis et al. 2010), associated with a weak differentiation in species composition and to the spatial distribution of some species in a complex mountain area; many xerophilous species were actually confined to small parts of the study area, probably because of Pleistocene glaciations history rather than of present environmental conditions. We cannot exclude however that the omission of other environmental properties, such as winter snow depth, human disturbances, localized grazing, fire history, etc., which may deeply influence vegetation composition (Guisan et al. 1998; Tsiripidis et al. 2010), might have contributed to the low inertia explained. Although the analysis was useful in showing how both substrate (Mgex and Niex in particular), and land use-related (C/N and P) chemical properties were important for plant community distribution, the effects that Ni has on species and community distribution was partially masked by the opposite effect of heat load, and therefore a definitive result was obtained only using RF.
A good association with serpentine soils was shown by several species, some of which are well known on serpentinite in various areas. Biscutella laevigata was well associated with high Ni and low values of exchangeable Ca also in nearby humid alpine valleys (D’Amico and Previtali 2012). Many species belonging to the Cerastium genus seem well adapted to serpentine soils in Mediterranean (Marsili et al. 2009) and in boreal habitats, where they showed adaptation to high concentrations of both Ni and Mg (Nyberg Berglund et al. 2004). Adaptation to serpentine conditions is reported also for Minuartia and Stachys genera: endemic subspecies of Minuartia laricifolia and Stachys recta grow in Mediterranean Italian ophiolitic outcrops, but they have not been recorded in serpentine soils on the Alps (Pignatti 1992). Both species (M. laricifolia and S. recta) in the study area are well associated with serpentine edaphic properties, respectively to high Niex and high Mgex values. The presence of these species was correlated with, respectively, Niex and Mgex also considering only serpentine soils. The adaptation of hyperaccumulator species was also well depicted. Considering all substrate lithologies, the presence of high concentrations of exchangeable Ni and Mg seems a favourable factor involved in the distribution of Alyssum argenteum in the xeric inner-alpine environment in the north-western Italian Alps. In fact, this species confirmed its selectivity for serpentinitic areas, but even if it typically grows in steppe or rocky outcrops (Pignatti 1992), it was able to thrive also under forest vegetation, although with smaller cover values. The localized inclusions of carbonates or allochtonous materials causing enrichment in exchangeable Ca did not inhibit the growth of this serpentine endemic Ni-hyperaccumulator species. Adaptation has been shown in other European serpentine areas for different Alyssum species: in particular, Gabbrielli et al. (1989) demonstrated a positive effect of high soil Ni on the metabolic efficiency of Alyssum bertolonii in Mediterranean Italy. Alyssoides utriculata, most common on serpentinite but growing also on non-serpentinic soil in the study area, has been sometimes observed hyperaccumulating Ni on particularly Ni-rich serpentinic substrates in North-western Italy (Roccotiello et al. 2010). High Mg was negatively associated only with Euphorbia cyparissias and Teucrium chamaedrys. Thus, high available Mg does not appear to be an important limiting factor for vegetation in the study area, when comparable poorly developed soils both on serpentine and on other parent materials are considered. The effect of Ni on species distribution was more marked than that of Mg based on the number of species that were negatively correlated with high Ni. The RF was extremely powerful in disentangling the respective importance of different factors, while taking into account the full set of soil and environmental properties, but the results were confirmed when only the typical factors of the serpentine syndrome were taken into account. The results are also visible observing the differences in the most important soil properties in the sites where the same species grow (Fig. 5 and electronic annex 2). The ratios between the average values of Niex, Mgex and Caex/Mgex in the sites where the species were present or absent are reported in Fig. 5 (distribution data in electronic annex 2) and clearly show a great discriminating ability of Niex with respect to both Mgex and the Ca to Mg ratio. Niex therefore seems the most important of the serpentine syndrome characteristics in shaping vegetation distribution on and off serpentinite in xeric inner-Alpine environment.

Ratios between mean values of serpentine syndrome factors (Niex, Mgex and Mgex/Caex) in the areas where the same species was absent or present; given the negative correlation values between Ca/Mg and Ni, the Mgex/Caex ratio is shown, in order to make the rankings of importance easier to be compared. The species shown are those having a rank of positive or negative correlations with the same serpentine factors in Table 3
However, when only serpentine-dominated sites were considered, the main ecological gradients were mostly related with microclimatic features, nutrients (whose cycling and bioaccumulation are related with xericity gradients) and Mgex. Niex, able to characterize serpentine soils as a whole despite its large concentration variability, disappeared from the important edaphic factors explaining both vegetation community distribution and species presence/absence. These results are similar to the ones obtained by many studies performed in many serpentine environments in the world, and appeared with a threshold of serpentinite abundance in the parent material of 60 %. Above 60 % serpentinite, significant differences were found between Ni, Mg and Ca/Mg compared to soils with less than 60 % serpentinite, serpentine endemics appeared and their frequency remained more or less stable above the threshold, indicating that relatively low amounts of serpentine may deeply influence soil and vegetation properties, without any measurable gradient at higher contents.
The differences between the two approaches to the study of the relationships between soil properties and vegetation in serpentinitic soils are therefore striking. Nutrient deficiencies were not specific of serpentinitic areas in the Alpine environment we have studied, and neither were the harsh site conditions depicted by e.g. heat load or rockiness. Ni therefore was probably the most important single edaphic factor in differentiating serpentine vegetation from non-serpentine. Its primary importance is verified by the strong decrease in model performances when this element was excluded from the analysis. Within the serpentinitic sites, many species which were negatively correlated with high Ni disappeared or became extremely rare, and the presence of the remaining ones was mainly associated to particular site conditions, such as bare soil or low tree cover (Helianthemum nummularium, Festuca valesiaca, Koeleria vallesiana, Potentilla neumanniana) or thick tree cover with large amounts of exchangeable Mg in the soil (Berberis vulgaris). Other fertility factors became more important for species presence, such as macronutrient and Ca abundance, which were not correlated to Ni availability when the serpentine-dominated subset was considered.
Conclusions
Soils developed in xeric, inner-alpine climates in the Alps underwent no important base and metal leaching, thus serpentine soils were still particularly rich in total and exchangeable Ni and Mg. Even though the Ca/Mg ratio was lower than on non-serpentine soils, the values were never excessively low, thanks to Ca bioaccumulation and carbonate inclusions. N, P, K scarcity characterized barren soils on every substrates, and did not characterize serpentine soils in particular, being associated with south-facing aspects, steep slopes and high heat load. Plant communities on serpentinite had a higher heterogeneity than non-serpentinite ones, thanks to the presence of several endemics, and differences in vegetation between serpentine and non-serpentine sites were correlated with Mg and Ni. Ni excess most often precluded the presence of plant species, while an exclusion due to Mg is rarer. Endemic species were instead mostly adapted to both factors.
When only serpentine-dominated areas were considered, vegetation variability was mostly linked with nutrient concentration gradients and microclimatic features.
Different approaches to the study of the “serpentine syndrome” thus lead to differently comparable results, in particular on the role of Ni as a driver for vegetation pattern. Nickel availability discriminated between serpentinitic and non serpentinitic areas, while climate and small variations in nutrient cycling and availability predominated at the high Ni background of serpentine soils.
References
Alexander EB, Coleman RG, Keeler-Wolf T, Harrison SP (2007) Serpentine geoecology of western North America. Oxford University Press, New York
Anderson MJ, Willis TJ (2003) Canonical analysis of principal coordinates: a useful method of constrained ordination for ecology. Ecology 84:511–525
Batianoff NG, Singh S (2001) Central Queensland serpentine landforms, plant ecology and endemism. S Afr J Sci 97:495–500
Blum JD, Dasch AA, Hamburg SP, Yanai RD, Arthur MA (2008) Use of foliar Ca/Sr discrimination and 87Sr/86Sr ratios to determine soil Ca sources to sugar maple foliage in a northern hardwood forest. Biogeochemistry 87:287–296
Bonifacio E, Zanini E, Boero V, Franchini-Angela M (1997) Pedogenesis in a soil catena on serpentinite in Northwestern Italy. Geoderma 75:33–51
Bonifacio E, Falsone G, Catoni M (2013) Influence of serpentine abundance on the vertical distribution of available elements in soils. Plant Soil 368:493–506
Brady KU, Kruckeberg AR, JrHD B (2005) Evolutionary ecology of plant adaptation to serpentine soils. Ann Rev Ecol Evol Syst 36:243–266
Breiman L (2001) Random forests. Mach Learn 45:15–32
Brooks RR (1987) Serpentine and its vegetation: a multidisciplinary approach. Dioscorides, Oregon
Brooks RR, Radford CC (1978) Nickel accumulation by European species of genus Alyssum. Proc R Soc Lond B Biol 200:217–224
Carter SP, Proctor J, Slingsby DR (1987) Soil and vegetation of the Keen of Hamar serpentine, Shetland. J Ecol 75:21–42
Cecchi L, Gabbrielli R, Arnetoli M, Gonnelli C, Hasko A, Selvi F (2010) Evolutionary lineages of nickel hyperaccumulation and systematics in European Alyssae (Brassicaceae): evidence from nrDNA sequence data. Ann Bot 106:751–767
Chardot V, Echevarria G, Gury M, Massoura S, Morel JL (2007) Nickel bioavailability in an ultramafic toposequence in the Vosges Mountains (France). Plant Soil 293:7–21
Chiarucci A (2004) Vegetation ecology and conservation on Tuscan ultramafic soils. Bot Rev 69:252–268
Chiarucci A, Rocchini D, Leonzio C, De Dominicis V (2001) A test of vegetation-environment relationships in serpentine soils of Tuscany, Italy. Ecol Res 16:627–639
Cutler DR, Edwards TC, Beard KH, Cutler A, Hess KT, Gibson J, Lawler JJ (2007) Random forests for classification in ecology. Ecology 88:2783–2792
D’Amico ME, Previtali F (2012) Edaphic influences on ophiolitic substrates on vegetation in the Western Italian Alps. Plant Soil 351:73–95
Del Moral R (1972) Diversity patterns in forest vegetation in the Wenatchee Mountains, Washington. Bull Torrey Bot Club 99:57–64
Evans JS, Cushman SA (2009) Gradient modeling of conifer species using random forests. Landsc Ecol 24:673–683
Gabbrielli R, Grossi L, Vergnano O (1989) The effects of nickel, calcium and magnesium on the acid phosphatase activity of two Alyssum species. New Phytol 111:631–636
Goudie AS, Middleton NJ (2001) Saharan dust storms: nature and consequences. Earth-Sci Rev 56:179–204
Guisan A, Theurillat JP, Kienast F (1998) Predicting the potential distribution of plant species in an alpine environment. J Veg Sci 9:65–74
Hennig C (2007) Cluster-wise assessment of cluster stability. Comp Stat Data Anal 53:258–271
IUSS Working Group WRB (2006) World reference base for soil resources 2006. World Soil Resources Reports No. 103. FAO, Rome
Jenny H (1980) The soil resource: origin and behavior. Ecol Stud 37:256–259, Springer-Verlag, New York
Kruckeberg AR (1984) California serpentines: flora, vegetation, geology, soils and management problems. University of California Press, Berkeley
Küfmann C (2002) Soil types and eolian dust in high-mountainous karst of the Northern Calcareous Alps (Zugspitzplatt, Wetterstein Mountains, Germany). Catena 53:211–217
Lal R, Follett F, Stewart BA, Kimble JM (2007) Soil carbon sequestration to mitigate climate change and advance food security. Soil Sci 172:943–956
Lazarus BE, Richards JH, Claassen VP, O’Dell RE, Ferrel MA (2011) Species specific plant-soil interactions influence plant distribution on serpentine soils. Plant Soil 342:327–344
Lee WG (1992) The serpentinized areas of New Zealand, their structure and ecology. In: Roberts BA, Proctor J (eds) The ecology of areas with serpentinized rocks, a world view. Kluwer, Dordrecht, pp 375–417
Lee BD, Graham RC, Laurent TE, Amrhein C (2004) Pedogenesis in a wetland meadow and surrounding serpentinic landslide terrain, northern California, USA. Geoderma 118:303–320
Legendre P, Legendre L (1998) Numerical ecology. Elsevier, Amsterdam
Liaw A, Wiener M (2002) Classification and regression by random forest. Rnews 2/3:18–22
Marsili S, Roccotiello E, Rellini I, Giordani P, Barberis G, Mariotti MG (2009) Ecological studies on the serpentine endemic plant Cerastium utriense Barberis. Northeast Nat 16:405–421
McCune B, Leon D (2002) Equations for potential annual direct incident radiation and heat load. J Veg Sci 13:603–606
Mercalli L (2003) Atlante climatico della Val d’Aosta. SMI eds, Bussoleno (To)
Nyberg Berglund AB, Dahlgren S, Westerbergh A (2004) Evidence of parallel evolution and site-specific selection of serpentine tolerance in Cerastium alpinum during the colonization of Scandinavia. New Phytol 161:199–209
Oksanen J, Blanchet FG, Kindt R, Legendre P, O’Hara RB, Simpson GL, Solymos P, Stevens MHH, Wagner H (2011) Vegan: community ecology package. R Package Version 2.0-0. http://CRAN.Rproject.org/package=vegan. Accessed 21 April 2013
Pignatti S (1992) Flora d’Italia. Vol. 1–3. Edagricole, Bologna
Proctor J (1997) Recent work on the ultramafic vegetation of Scotland. Bot J Scot 49:277–285
Proctor J, Nagy L (1991) Ultramafic rocks and their vegetation: an overview. In: The vegetation of ultramafic (Serpentine) soils. Proceedings of the First International Conference on Serpentine Ecology. Intercept Lid., Andover (England)
Reddy RA, Balkwill K, Mclellan T (2009) Plant species richness and diversity of the serpentine areas on the Witwatersrand. Plant Ecol 201:365–381
Roberts BA, Proctor J (1992) The ecology of areas with serpentinized rocks, a world view. Kluwer, Dordrecht
Roccotiello E, Zotti M, Mesiti S, Marescotti P, Carbone C, Cornara L, Mariotti MG (2010) Biodiversity in metal-polluted soils. Fresenius Environ Bull 19(10b):2420–2425
Soil Survey Staff (2004) Soil survey laboratory methods manual, Soil Survey Investigations Report No. 42
Tsiripidis I, Papaioannou A, Sapounidis V, Bergmeier E (2010) Approaching the serpentine factor at a local scale—a study in an ultramafic area in northern Greece. Plant Soil 329:35–50
Van der Ent A, Baker AJM, Reeves RD, Pollard AJ, Schat H (2013) Hyperaccumulators of metals and metalloid trace elements: facts and fiction. Plant Soil 362:319–334
Vayssiéres MP, Plant RE, Alen-Diaz BH (2000) Classification trees: an alternative non-parametric approach for predicting species distribution. J Veg Sci 11:679–694
Acknowledgments
This study was performed thanks to the research agreement between the University of Turin, NATRISK centre, and Regione Autonoma Valle d’Aosta, Department of Soil protection and Water Resources. We also thank two anonymous reviewers and the Editor of Plant and Soil journal for the useful comments on previous versions of this paper.
Author information
Authors and Affiliations
Corresponding author
Additional information
Responsible Editor: Henk Schat.
Rights and permissions
About this article
Cite this article
D’Amico, M.E., Bonifacio, E. & Zanini, E. Relationships between serpentine soils and vegetation in a xeric inner-Alpine environment. Plant Soil 376, 111–128 (2014). https://doi.org/10.1007/s11104-013-1971-y
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11104-013-1971-y