
Abstract
Background and aims
Physical and chemical soil properties determine local plant conditions and resources, affecting plants’ ability to respond to disturbances. In alpine grasslands, wild boar disturbances occur at different intensities, what may affect differently their soil properties. Alpine soils from five contrasted plant communities were explored within and outside disturbances, accounting for an overall and community scale effect. Additionally, we analysed the effect of disturbance intensity on soil NO3 --N and NH4 +-N.
Methods
Soils were analyzed for physical (bulk density, moisture content and electrical conductivity), and chemical properties (pH, total N and C, oxidizable C, C:N ratio, available K, P, Ca2+, Na+ and Mg2+). Resin bags were used to compare the effect of the disturbance occurrence and intensity on soil NO3 --N and NH4 +-N.
Results
Bulk density, total N and NO3 --N concentration were significantly higher in disturbed areas, while soil moisture, C:N, NH4 +-N, Na+, Mg2+ and Ca2+ concentrations were significantly lower. However, low disturbance intensity reduced NO3 --N and increased NH4 +-N concentrations.
Conclusions
Wild boar occurrence and intensity strongly alter physical and chemical conditions of alpine soils, increasing soil compaction, and altering the availability of N forms. These changes may affect most plant species, thus affecting the structure and dynamics of alpine plant communities.
Similar content being viewed by others


Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
Soils are essential parts of terrestrial ecosystems, as they physically support primary producers and decomposers, maintain biotic communities and determine key processes such as nutrient cycling or material-energy flows throughout the system (Brady and Well 2002; Lavelle et al. 1997). At the ecosystem and community levels, soils determine the structure and dynamics of communities through their response to environmental fluctuations and disturbances (Pickett and White 1985). The effects of soil disturbance to ecosystem functioning, will depend on ecosystem structure and the processes affected by the disturbance (Pickett and White 1985). For instance, sensitivity to soil disturbance may be exacerbated in alpine ecosystems, because most plant biomass is within the first few cm of the soil and its response is limited by a relatively short growing season (García González 2008; Körner 1999).
Wild boars are one of the most widely distributed mammals worldwide (Olivier and Reeder 2008), being one of the main agents causing soil disturbance in the alpine habitats they inhabit (Bueno et al. 2009; Massei and Genov 2004; Olivier and Reeder 2008). The primary means by which wild boars disturb soils is by turning over the soil looking for underground feeding resources (hereafter ‘rooting’), which include nutritive plant and animal foods, such as rhizomes, bulbs, fungi, earthworms, insect larvae, and even small underground vertebrates (Schley and Roper 2003). Although rooting can cause extensive disturbance to soils, affecting up to hundreds of hectares (Bueno et al. 2009; Massei and Genov 2004; Welander 2000), it is unclear if rooting activities may enhance or degrade soil conditions for plants. In fact, few studies have investigated the effects of wild boar disturbances to soils, relative to the great diversity of ecosystems wild boars inhabit (Barrios-Garcia and Ballari 2012).
Soil disruption by wild boars may affect physical, and chemical soil properties (Barrios-Garcia and Ballari 2012; Brady and Well 2002), where some of them, such as bulk density, can be largely affected by rooting, whereas others like soil moisture may remain unaltered (Barrios-Garcia and Ballari 2012). Singer et al. (1984) found a decrease in bulk density of rooted soils in beech forests of Tennessee, related with the mixing of the organic matter and mineral horizons. In turn, soil moisture did not change after rooting, neither in costal grasslands and oak forests of California (Cushman et al. 2004; Moody and Jones 2000), nor in Mabi forests of Queensland, Australia (Mitchell et al. 2007), but see Wirther et al. (2011). Broadly, soil chemical properties, can be divided in two groups regarding their response to rooting: one group has a positive or neutral response, with an increase or no change in its concentration following disturbance, and the other with decreasing or neutral concentration values. Specifically, N and its mineral compounds, NO3 --N and NH4 +-N, belong to the first group. Changes in N have led to arguments on a neutral (Bruinderink and Hazebroek 1996; Cushman et al. 2004; Moody and Jones 2000; Tierney and Cushman 2006) or a positive effect of rooting (Siemann et al. 2009; Sims 2005; Singer et al. 1984) through the increase in soil nutrients. On the other hand, C, P and exchangeable cations, Ca2+, Na+, K+, and Mg2+, belong to the second group, where neutral or negative effects on plants, have been argued through a potential nutrient loss by leaching (Lacki and Lancia 1983; Mohr et al. 2005; Moody and Jones 2000; Singer et al. 1984).
No study has covered the effect of wild boar rooting on the properties of alpine soils, where the high recurrence of rooting activities (Bueno et al. 2009; Welander 2000) may enhance their positive or negative effects, or both. Soil disturbances caused by other mammals, such as pocket gophers (Thomomys talpoides) (Laycock and Richardson 1975; Litaor et al. 1996; Sherrod and Seastedt 2001), moles (Talpa europaea) (Canals and Sebastiá 2000) or grizzly bears (Ursus arctos horribilis) (Tardiff and Standford 1998) affect alpine soils. However, these disturbances differ in their frequency, extent and effects on soil properties, particularly in the way each mammal disturbs soil, i.e. burial, excavation or rooting (Hole 1981; Kotanen 1997). Even the effect of a given mammal species can differ among habitats and soils (Canals and Sebastiá 2000), so the effects of wild boar rooting on alpine soils cannot be directly inferred from disturbances caused by other mammals.
The aim of this study is to determine the effects of wild boar rooting on soils of alpine grasslands in the Spanish Central Pyrenees, comparing soil physical (i.e. electrical conductivity, bulk density and moisture) and chemical (i.e. pH, C, N, P, and exchangeable cation content: Ca2+, Na+, K+, and Mg2+) properties, within and outside areas disturbed by wild boars. In addition, the study evaluates the potential effects of soil disturbances on the availability of nutrients; by examining the relationship between soil mineral N (i.e. NO3 --N and NH4 +-N) and the occurrence and intensity of wild boar disturbances. We expect wild boar rooting to have an effect on the physical and chemical soil properties of alpine grasslands, as this ecosystem functionally relies on belowground biomass located within the depth range of this disturbance. Specifically we hypothesized that: 1) wild boar rooting will decrease bulk density by mixing organic matter with mineral soil. 2) Wild boar rooting will simultaneously have positive effects, through increasing the availability of N forms, and negative effects to plants, through decreasing the availability of P and cations. 3) Rooting intensity (relative to occurrence) will increase the concentration of N mineral compounds up to a point, where the soil system might get exhausted. Determining how this extensive and recurrent disturbance affects alpine soil properties is critical for understanding how this disturbance might compromise the preservation of alpine soils and affect grassland dynamics.
Methods
Study area
The study was carried out in Ordesa and Monte Perdido National Park (OMPNP) (42º 36′N, 0º 00′) in the Spanish Central Pyrenees. The study area comprises natural grasslands between 1,500 and 2,800 m a.s.l. with an average annual temperature of 5 °C and an average annual precipitation of 1,750 mm, most of which occurs in spring and autumn (García-González et al. 2007). The geology is mainly limestone and calcareous sandstone, while the topsoil has a clay-loam texture, with illite as dominant clay. The soils are acidic, desaturated of bases, and have high organic matter content and structural stability in the A-horizon (umbric epipedon). Thus, the soil profiles are Humic Dystrocryepts (Badía et al. 2008, 2002).
We selected the five grassland communities most extensively disturbed by wild boars, which are among the most widespread alpine grassland communities in the Pyrenees (Bueno et al. 2009). The topography among those dense grasslands is smooth with slopes < 20º (Bueno et al. 2009). Three of the plant communities were each dominated by a single grass: Festucion paniculatae (FP, terms following phytosociological nomenclature) by Festuca paniculata, Festucion eskiae (FE) by Festuca eskia and Nardion strictae (NS) by Nardus stricta. The other two are co-dominated by more than three species; Briomion erecti (BE) by Festuca gr. rubra, Agrostis capillaris, Lotus alpinus, etc., and Rumicion pseudoalpini (RP) by Poa supina, Rumex pseudoalpinus, Chenopodium bonus-henricus, etc. (Fillat et al. 2008).
Grazing activities in the area, mainly by sheep and cattle, date back to at least the Middle Ages (Montserrat and Fillat 1991) and differ in the abundance of livestock (Fillat et al. 2008). Average stocking rates for each community describe a grazing gradient, from the most grazed (RP: 0.50 SLU/ha) to the least (NS: 0.06 SLU/ha) specifically: RP > FP > BE > FE > NS (Bueno et al. 2011). These grazing activities should not be considered disturbances, as their historic use and prevalence are strongly linked with the origin and development of most Pyrenean alpine grasslands (Fillat et al. 2008; Montserrat and Fillat 1991).
The plant communities evaluated covered 54.3 % (2,773 ha) of the study area, but nearly 98 % of the wild boar disturbances occurred there (Bueno et al. 2009). The high recurrence of rooting, locally, made it very difficult to estimate the precise age of individual disturbances (Bueno et al. 2009; Welander 2000); therefore, we compared soils of areas where the vegetation was either disturbed or undisturbed, irrespective of age of disturbance. Disturbed patches had been rooted by wild boar, recently or within the previous 2 years and were identified by the presence of overturned pieces of turf, which differed from the signs of disturbance caused by other mammal species (Bueno et al. 2009). Undisturbed patches were completely covered by vegetation without any presence of overturned turf and forming a continuum with the surrounding grassland.
Sampling design
To quantify the effects of wild boar rooting on alpine grassland soils, we used a random stratified paired sampling design. In each one of the five plant communities, 20 sampling points were randomly distributed within, and 20 outside areas disturbed by wild boar in a pairwise fashion (200 points in total). We used a previous disturbance map (Bueno et al. 2009) to randomly locate the points in the disturbed area using ArcGis 8.3 (ESRI). Then, in the field, we located the undisturbed areas, closest to the previous points. In 2006, 100 soil samples (20 per community, 10 outside and 10 within disturbed areas) were collected with a cylindrical metal core sampler that extracts a soil sample of 5 cm diameter and 10 cm deep. Disturbance depth relative to the surrounding soil level was assessed by taking 300 depth measures within wild boar disturbances and calculating the average disturbance depth (10.7 ± 0.3 cm, mean ± standard error) in the study area. In August, 2007, at the peak of the growing season, 200 ion exchange resin bags were placed 10 cm below ground and left there for 45 days, for measuring the available mineral N content in the soils (NO3 --N and NH4 +-N). The geographical coordinates of each sampling point were recorded using a GPS (Garmin Etrex Venture, USA).
Disturbance intensity
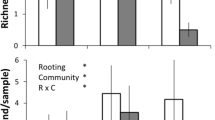
Disturbance intensity was estimated by calculating the approximate volume (m3) of soil that was overturned by wild boar rooting. At each sampling point within disturbed patches, two measures of disturbance depth and two orthogonal diameters of each disturbed patch were recorded. In addition, we approximated the shape of each disturbance (squared, rectangular, circular, ellipsoid or triangular). To calculate the volume of each disturbance, the average depth was multiplied by the area of the disturbance, which served as a proxy for disturbance intensity, based on the assumption that the more intensively wild boars disturbs the larger the volume of soil that is overturned. Disturbance intensity differed among plant communities (highest in FE and lowest in FP), which indicated that wild boar rooting impacted alpine grasslands, differentially (Fig. 1).

Wild boar rooting intensity (log-transformed) in the five most extensively disturbed alpine grasslands in the Central Pyrenees, Spain. Lower-case letters indicate significant differences between plant communities based on a non-parametric multiple test at alpha < 0.05. BE Bromion erecti; FE Festucion eskiae; FP Festucion paniculatae; NS Nardion strictae; RP Rumicion pseudoalpini
Soil analyses
For all measurements except moisture content, samples were air-dried, sieved through a 2-mm mesh and ground finely. Moisture content was quantified by calculating the amount of mass lost by a 2-g soil sample after it was dried at 105 °C for 24 h. Bulk density was calculated by dividing the mass of each soil core sample by its volume (Klute 1986). Soil pH was measured in a 1:5 soil water suspension after shaking for 2 h (Buurman et al. 1996). Total C, oxidizable C, as well as total N were determined by an Elementar Analysen Systeme GMBH Autoanalyzer (Page 1982). To quantify organic matter, oxidizable C content was multiplied by the Van Bemmelen factor (1.724) (Heanes 1984). A 0.5-M sodium bicarbonate extraction at a nearly constant pH of 8.5 (Page 1982) quantified available P. To extract soil cations (Ca 2+, Mg2+, Na+ and K+) 10-g sub-samples were shaken for 1 h in a 1 M ammonium-acetate solution at pH 7, and concentration was determined with ICP-OES (Thermo iCAP 6,000 series). The C:N ratio was calculated by dividing the proportion of oxidizable C by the proportion of total N of each sample.
To calculate the relative concentrations of soil mineral N (NH4 +-N and NO3 --N) ion exchange resin bags were used; specifically 8.5 g wet mass of cation and anion mixed bed ion exchange resin (J.T. Baker mixed-bed ion exchange resin, VWR Scientific Products) were placed in nylon stockings, thermo-sealed and cut (Binkley and Matson 1983; Page 1982). Resin bags were then buried at 10 cm for 45 days in the field. Once the resin bags were retrieved, they were air-dried for 2 days, before the cations were extracted using 100 ml of 1 M KCl (Tardiff and Standford 1998). NH4 +-N and NO3 --N concentrations were determined colorimetrically (Maynard and Kalra 1993; Tardiff and Standford 1998).
Statistical analyses
Soil properties
To determine which soil properties were significantly altered by the occurrence of wild boar disturbances, two complementary analyses were performed: a linear mixed model (LMM) and a Wilcoxon signed-rank test. The LMM was used to analyze the relative effect of disturbances in all the plant communities studied, where each soil property was the response variable for each model, and the presence/absence of disturbances, the plant communities (that were transformed into dummy variables) (Quinn and Keough 2002), and their interaction, were introduced as predictors. To account for a possible lack of independence between paired samples, mixed-effects models were used (Zuur et al. 2009). Specifically, a “pair” factor was added to the model as the random factor, which was estimated by assigning to each pair of observations (n = 100; disturbed-undisturbed) a number from 1–50. Because of the lack of homogeneity in residuals spread across the different plant communities, we added an identity variance structure to the models that allows for different variances for each community. Three soil properties (electrical conductivity, P and K) were log-transformed to account for normality requirements in the residuals. To select the final model, likelihood ratio tests were used to compare the models with and without each predictor variable, non-significant terms were dropped from the final models (Zuur et al. 2009). A correlogram of the model residuals (Dormann 2007) did not reveal any significant spatial autocorrelation.
To evaluate the effects of wild boar rooting on each soil property within each plant community, non-parametric Wilcoxon Signed Rank tests were used because soil samples were paired and normality could not be assumed (Moore et al. 2008). These analyses complemented the LMM because they can identify the effects on soil properties that are specific to each plant community, which might go undetected in LMM.
Soil mineral N
To examine the relationship between available mineral N (NO3 --N and NH4 +-N) and the occurrence and intensity of disturbances in the soils from the five grassland communities, two complementary analyses were performed, both attending to the occurrence and intensity of disturbances: LMMs and partial redundancy analyses (pRDA) (Legendre and Legendre 1998). LMMs captured the response of NH4 +-N and NO3 --N in all communities, while pRDA described how plant communities differed based on their soil concentration of NH4 +-N and NO3 --N. The LMMs with the occurrence of disturbance as predictor, were carried out in the same way as LMMs in the Soil properties section, but with NH4 +-N and NO3 --N as response variables. For the LMs taking into account the intensity of rooting, two aspects differentiated them from to the previous LMMs: the data only contained samples within disturbances (n = 100), so without the random factor, and the predictor disturbance occurrence was replaced by the intensity of rooting.
To quantify the relationship between soil NH4 +-N and NO3 --N content, and the occurrence (presence and absence) and intensity of wild boar disturbances pRDAs were used. For rooting occurrence, the response matrix was composed by 200 observations (rows) and 10 binary vectors (columns), of which five represented the presence (‘1’) or absence (‘0’) of plant communities within disturbed soils and five represented undisturbed soils. The environmental matrix comprised two columns: the concentrations of NO3 --N and NH4 +-N in each of the sampling points (n = 200). To quantify the relationship between the concentrations of mineral N compounds in the soil and the intensity of disturbances, the response matrix contained 100 observations (the samples from disturbed soils, only), and the five vectors that represented the disturbance intensity in each of the five plant communities comprised the environmental matrix. To detect multicollinearity among predictors VIF and Pearson correlation tests were used, and all of the predictors had values <0.3 and <0.5 respectively. By incorporating the axis obtained after applying spatial eigenvector maps (SEVM) from the distance matrix of the geographical coordinates on the observations (Dormann 2007), spatial autocorrelation in the data was taken into account in the model.
The SEVM procedure extracts the potential spatial component of the model in orthogonal vectors, which can be included in regression or ordination models to account for spatial dependencies at different scales (Dray et al. 2006; Griffith and Peres-Neto 2006). A forward selection of spatial predictors was performed (Blanchet et al. 2008; Griffith and Peres-Neto 2006) before SEVM axes were incorporated into pRDAs. To test for spatial autocorrelation, the Mantel correlogram of the residuals (Legendre and Legendre 1998) and a global test of spatial autocorrelation using Moran’s Index (Dormann 2007) was calculated. Spatial dependencies were not detected with the incorporation of selected SEVM predictors into the pRDAs.
All of the statistical analyses were performed with R 2.13.0 (R Development Core Team 2009); specifically npmc for non-parametric multiple comparisons, lme for LMM, ncf for correlograms, vegan for pRDA, packfor for forward selection of spatial predictors, and spdep and spacemakeR for SEVM.
Results
Effects of wild boar rooting on soil properties
Wild boar rooting had a significant effect on all properties measured, in all or some of the plant communities studied, with the exception of pH, K concentrations and the properties based on C such as total C, oxidizable C and organic matter (Tables 1, 2 and 3).
Regarding the physical properties, in all plant communities except FP, the bulk density and moisture content of soils was higher in disturbed patches than in undisturbed ones (Table 1; Fig. 2), being significant for each community in the case of the bulk density (Table 3; Fig. 2). Among chemical properties the concentrations of N were significantly higher in disturbed than in undisturbed patches (Table 1; Fig. 2), while the concentration of Mg2+, Na+ and C:N ratio tended to be lower within disturbed areas than in undisturbed areas (Table 1; Fig. 2). Finally the responses of P and Ca were community-dependent, where Ca concentrations increased in RP and BE communities, while both decreased in FP community within disturbed areas (Table 3).

Fitted values of the significant soil properties that differ in undisturbed and disturbed areas by wild boar rooting. Fitted values were obtained from the adjusted Linear Mixed Models (see Table 2 for details) with respect to the occurrence of disturbances in the five most disturbed plant communities in the alpine grasslands of the Spanish Pyrenees, Spain. a-b Physical properties. c-g Chemical properties
Among communities, in undisturbed areas soil pH was lowest in the FE community (Table 4), and C:N ratios were highest in the three communities with a single dominant grass species (FE, NS & FP; Table 4). In FP, the most productive community in terms of biomass, C:N ratio was lowest in disturbed areas (Table 3), which probably reflected a high decomposition rate of organic matter because the amount of soil organic matter was highest in disturbed areas (Table 3). Both in disturbed and undisturbed sites, Ca soil concentrations were lowest in the FE community (Table 3), probably because the soils were acidic and received limited input of Ca from livestock.
Soil mineral N
The amount of NO3 --N and NH4 +-N in alpine soils was strongly influenced by the occurrence and intensity of wild boar disturbances (Table 2; Fig. 3a and b). In all five plant communities, wild boar rooting significantly increased the mineralization rate of N as reflected by soil NO3 --N concentrations (Table 2; Fig. 3a and 4a). In the communities dominated only by tall-tussock grass (FP and FE), disturbances have a significant (p < 0.05) effect on the NH4 +-N concentration in soils (Fig. 3b and b). Specifically, soil NO3 +-N concentrations were negatively correlated with the presence of wild boar rooting in FE communities (Fig. 4a) and positively correlated with disturbance intensity in FP communities (Fig. 4b). The trend in soil NO3 --N and NH4 +-N concentrations changed when disturbance intensity reached a value around 5 m3, indicating a clear intensity threshold for soil mineral processes in these grasslands (Fig. 3c and b).

Concentrations of soil NO3 --N and NH4 +-N with respect to the occurrence of wild boar disturbances (a-b) and to the intensity of disturbances (c-d), in the alpine grasslands of the Spanish Pyrenees, Spain. Concentration values were previously fitted using Linear Mixed Models (see Table 2 for details). Disturbance intensity values were log-transformed for improving model fit. For detailed visualization of the relationship between disturbance intensity and soil nitrate and ammonium concentrations, (c) and (d) plots respectively, an adjusted local smoothed regression line (blue) with its 95 % confident intervals (gray) was used. Black vertical lines indicate the level of disturbance intensity were the concentration trend of NO3 --N and NH4 +-N started to change (at disturbance intensity of 4.95 m3 in both cases)

Concentrations of the exchangeable soil NO3 --N and NH4 +-N in the occurrence of disturbances by wild boar (a) and with respect to disturbance intensity (b), among the five most disturbed plant communities in the alpine grasslands of the Central Pyrenees, Spain, modelled with pRDA. Axes were significant (p < 0.05) based on a permutation test (N = 1,000 permutations), except for NO3 --N axis in (b). BE Bromion erecti; NS Nardion strictae; RP Rumicion pseudoalpini; FE Festucion eskiae; FP Festucion paniculatae
Discussion
In the alpine grasslands of the Spanish Pyrenees, wild boar rooting causes large disturbances to both physical and chemical soil properties, affecting on average the upper 10 cm of the soil, where wild boars may found most of their food (Herrero et al. 2005; Singer 1981). This is the first reference to the effect of wild boars rooting on soils in alpine habitats, although such effects have been observed in other ecosystems worldwide (Barrios-Garcia and Ballari 2012; Groot Bruinderink and Hazebroek 1996; Kotanen 1994; Tierney and Cushman 2006).
Contrary to our expectations, wild boar rooting increased bulk density in alpine grasslands in all plant communities but FP, where values were similar within and outside disturbances. Wild boars turn over the soil, so an immediate effect might be a reduction in bulk density in disturbed areas; however, rooting removes the plant cover along with the root fraction, which provides a physical support to alpine soils (Pohl et al. 2009). Without the structural support provided by the roots, the effect of livestock trampling in summer and snow weight in winter can easily compact the disturbed soils. In the absence of factors that compact soil, e.g. livestock trampling, disturbed soils can have reduced bulk density; for instance in mountain forests of Tennessee, USA (Lacki and Lancia 1983; Singer et al. 1984). On the contrary, in mountain rangelands of Oregon, USA, the presence of sheep increased the bulk density of soils disturbed by pocket gopher digging (Laycock and Richardson 1975). The effect of plant cover removal on bulk densities is inferred by the invariance in the bulk densities of soils when disturbances do not remove plant cover; e.g., disturbances caused by a vehicle in Alaska (Chapin and Shaver 1981). Within disturbed areas in FP grasslands, a substantial amount of litter from F. paniculata is mixed with the soil, which might ameliorate soil compaction rates in this community. This was the case in Tennessee, where the relatively greater amounts of organic matter within areas disturbed by feral pigs reduced the bulk density of the soils in these locations (Lacki and Lancia 1983).
As expected, wild boars had a simultaneous positive and negative effect on nutrient concentrations. With their rooting, wild boars may alter most of the biotic communities that are responsible for most of the nutrient cycling processes in soils (Lacki and Lancia 1983; Singer et al. 1984). In the Pyrenees, wild boar rooting, increased the availability of soil N and NO3 --N, decreased it for Mg2+, and Na+ but did not alter significantly the concentrations of soil NH4 +N, nor K+. Disturbance can break up stable aggregates of soil particles, increase the mineralization of soil organic matter, with the subsequent release of available nutrients (Canals et al. 2003) The increase in the release of soil nutrients can be partly absorbed by plants if disturbances are small and they do not remove entire root systems. In addition, nutrients can be also stored in the soil in available forms, which increases their availability in disturbed areas e.g., disturbance to the soil by pocket gopher disturbances in Utah (Laycock and Richardson 1975) and tuco-tuco (Ctenomys talarum) in Argentina (Malizia et al. 2000). However, if disturbances are large enough, such as those created by wild boars, plant roots can be dramatically affected and these nutrients are easily leached by rain or snowmelt (Lacki and Lancia 1983; Singer et al. 1984), which reduces their concentrations in disturbed soils.
In Pyrenean alpine soils after wild boar rooting, N mineral forms (NO3 --N & NH4 +-N) were altered by disturbance intensity until a value 5 m3 was reached. This value corresponds for example, to a square of 7 m side and 10 cm depth. Relatively larger or deeper disturbances, those with higher disturbance intensity, may not affect the mineralization processes for these soil N forms. In addition, and regarding the disturbance common size in the area, higher that the aforementioned size (C.G. Bueno pers. obs), we can expect that this mineralization saturation may be reached easily.
Soil NO3 --N concentration is the result of a complex tradeoff; NO3 --N ‘input’ that is the result N mineralization rates, while the ‘output’ depends on plant nitrogen uptake, nitrogen immobilization by decomposers, leaching or run-off and denitrification rates (Vitousek 1985). In the soils of mountain forests of Tennessee (Singer et al. 1984) and Hawaii (Vitousek 1986) the effects of rooting by wild boar or feral pig on NO3 --N concentrations were similar to those observed in the Pyrenees, but simulated feral pig disturbances in coastal grasslands of California either, increased NH4 +-N availability concentrations only (Kotanen 1997), or had no significant effect on either form of N (Cushman et al. 2004). The variation among habitats in their responses to similar types of disturbances suggests, perhaps, that the habitat differs in the composition of the NO3 --N trade-off, in its sensitivity to disturbance or both.
In general, the reduction of plant cover by rooting might reduce plant NO3 --N uptake, accelerate mineralization because of the soil aeration, and reduced nitrogen immobilization by decomposers (bacteria), which has been described for pocket gopher soil disturbances (Canals et al. 2003). In the FE grasslands of the Pyrenees, which was the community most intensively disturbed by wild boar rooting, the concentrations of NO3 --N and NH4 +-N were inversely correlated. It is possible that the aeration effect of disturbances influenced the nitrification of NH4 +-N because if the intensity of disturbances increased the availability of oxygen in the soil, it would facilitate the nitrification of NH4 +-N and thereby, increase NO3 --N concentrations (Brady and Well 2002), as follows:
Nevertheless, the impact of wild boar disturbances on the relative abundance of each N-form may depend on the intensity of disturbance and the environmental characteristics and dynamics of each community. For instance, in FP grasslands, only, disturbance intensity and NH4 +-N concentrations were positively correlated. FP is the most xeric of the five alpine communities studied (Benito 2006; Fillat et al. 2008), and the soils there had the highest organic matter content in disturbed areas and the highest concentrations of total C and N in undisturbed areas. The dominant species in FP community, F. paniculata, has one of the highest rates of biomass productivity among the Pyrenean grasslands (Fillat et al. 2008). Intensive and frequent disturbances can cause a steady increase in the amount of oxygen in the soil, but in turn, reduce water soil content because of the loss of plant cover, which prevents the direct evaporation of soil water. An increase in the amount of oxygen in the soil can increase the ammonification of organic N to NH4 +-N (Brady and Well 2002). This would reduce the rate at which NO3 --N is formed for FP community, probably because of the high intensity of soil disturbance and the abovementioned xeric conditions of this community that would produce a reduction in the activity of nitrifying bacteria (Stark and Firestone 1995).
Our results pointed out a shift on soil conditions for plants, in particular an increase in soil compaction and availability of N forms. These effects, with an intrinsic reduction on plant competition, filter the species, which may reach these gaps, or survive rooting. In fact different plant compositions have been described for disturbed, when compared to closest undisturbed, vegetation patches (Barrios-Garcia and Ballari 2012). Species diversity within disturbed areas usually decreases (Barrios-Garcia and Ballari 2012; Kotanen 1994), but at the local scale; if disturbance recurrence holds populations of fugitive or disturbance-tolerant species, the overall species richness may increase (Welander 1995). However, it is unclear if diversity at regional scales and among affected communities (beta diversity) may benefit from wild boar rooting. In any case, in the Pyrenean alpine grasslands, two plant functional groups have been pointed out to increase their abundance due to wild boar rooting: annuals and geophytes (Bueno 2011; Palacio et al. in rev). On one hand, annuals are enhanced by rooting through stimulation of their seed banks, although they are very scarce and highly dependent on livestock grazing activities (Bueno et al. 2011). On the contrary, geophyte communities seems to take advantage of the increase in the availability of N form after rooting, increasing their fit and, likely, their dynamics (Palacio et al. in rev).
Conclusion
In summary, in the Pyrenean grasslands, wild boar rooting deeply affects soil properties because it alters soil structure, compacts the soil and modifies its composition. The effect is different and more substantial as the intensity of disturbance increases; i.e., mineralization rates are higher until the saturation point is reached, which leads to deeper changes in soil properties. Nutrients such as NO3 --N or available P increase because rooting removes plants and might alter decomposer communities, which are no longer available to take up these nutrients and immobilize them. In addition, those nutrients may remain available to plants unless rain and snowmelt leach them, which suggest that differences in the availability of nutrients might contribute to an increase in the diversity of plant functional groups whenever rooting is not excessively intense.
References
Badía D, García González R and Martí C. Clasificación de suelos en pastos alpinos de Aisa y Ordesa (Pirineo Central). Edafología. 2002;9.
Badía D, Fillat F, Martí C, Aguirre J, Gómez D, Sánchez JR (2008) Soil chemistry in long-term livestock camping areas in Pyrenean summer pastures. Agrochimica 52:188–99
Barrios-Garcia MN, Ballari S (2012) Impact of wild boar (Sus scrofa) in its introduced and native range: a review. Biol Invasions 14:2283–300
Benito JL (2006) Vegetacion del Parque Nacional de Ordesa y Monte Perdido (Sobrarbe, Pirineo Central Aragonés). Publicaciones del Consejo de Protección de la Naturaleza de Aragón, Zaragoza
Binkley D, Matson P (1983) Ion exchange resin bag method for assessing forest soil nitrogen availability. Soil Sci Soc Am J 47:1050–2
Blanchet FG, Legendre P, Borcard D (2008) Forward selection of explanatory variables. Ecology 89:2623–32
Brady NC and Well RR. The nature and properties of soils. Prentice Hall; 2002. pp. 960.
Bruinderink GWTAG, Hazebroek E (1996) Wild boar (Sus scrofa scrofa L.) rooting and forest regeneration on podzolic soils in the Netherlands. For Ecol Manage 88:71–80
Bueno CG. Las perturbaciones de jabalí en los pastos alpinos del Pirineo Central: una aproximación multiescalar. In: Departamento de Agricultura y Economía Agraria. Zaragoza: Universidad de Zaragoza; 2011. p. 191.
Bueno CG, Alados CL, Gómez-García D, Barrio IC, García González R (2009) Understanding the main factors in the extent and distribution of wild boar rooting on alpine grasslands. J Zool 279:195–202
Bueno CG, Reiné R, Alados CL, Gómez-García D (2011) Effects of large wild boar disturbances on alpine soil seed banks. Basic Appl Ecol 12:125–33
Buurman P, van Lagen B, Velthorst EJ (1996) Manual for soil and water analysis. Backhuys, Wageningen, p 314
Canals RM, Sebastiá MT (2000) Soil nutrient fluxes and vegetation changes on molehills. J Veg Sci 11:23–30
Canals RM, Herman DJ, Firestone MK (2003) How disturbance by fossorial mammals alters N cycling in a California annual grassland. Ecology 84:875–81
Chapin FS III, Shaver GR (1981) Changes in soil properties and vegetation following disturbance of Alaskan artic tundra. J Appl Ecol 18:605–17
Cushman JH, Tierney TA, Hinds JM (2004) Variable effects of feral pig disturbances on native and exotic plants in a California grassland. Ecol Appl 14:1746–56
Dormann CF (2007) Effects of incorporating spatial autocorrelation into the analysis of species distribution data. Glob Ecol Biogeogr 16:129–38
Dray S, Legendre P, Peres-Neto PR (2006) Spatial modelling: a comprehensive framework for principal coordinate analysis of neighbour matrices. Ecol Model 196:483–93
Fillat F, García González R, Gómez D, Reiné R (2008) Pastos del Pirineo. CSIC - Diputación de Huesca, Madrid, p 319
García González R. Management of Natura 2000 habitats. Alpine and subalpine calcareous grasslands. In: Technical Report 2008 11/24. Ed. E Commission; 2008. p. 23.
García-González R, Alados CL, Bueno G, Fillat F, Gartzia M, Gómez D et al (2007) Valoración ecológica y productiva de los pastos supraforestales en el Parque Nacional de Ordesa y Monte Perdido. In: Ramirez L, Asensio B (eds) Proyectos de investigación en Parques Nacionales: 2003–2006. OAPN-MMA, Madrid, pp 105–28
Griffith DA, Peres-Neto PR (2006) Spatial modeling in ecology: the flexibility of eigenfunction spatial analyses. Ecology 87:2603–13
Heanes DL (1984) Determination of total organic-C in soils by an improved chromic acid digestion and spectrophotometric procedure. Commun Soil Sci Plant Anal 15:1191–213
Herrero J, Irizar I, Laskurain NA, García-Serrano A and García González R. Fruits and roots: wild boar foods during the cold season in the southwestern Pyrenees. Ital J Zool. 2005.
Hole FD (1981) Effects of animals on soil. Geoderma 25:75–112
Klute A (1986) Methods of soil analysis. Part 1 physical and mineralogical methods. American Society of Agronomy, Madison, p 1188
Körner C (1999) Alpine plant life. Functional plant ecology of high mountain ecosystems. Springer, Berlin, p 338
Kotanen P (1994) Effects of feral pigs on grasslands. Fremontia 22:14–7
Kotanen P (1997) Effects of experimental soil disturbance on revegetation by natives and exotics in coastal California meadows. J Appl Ecol 34:631–44
Lacki MJ, Lancia RA (1983) Changes in soil properties of forests rooted by wild boar. Proc Annu Conf Southeasth Assoc Fish Wildl Agencies 37:228–36
Lavelle P, Bignell D, Lepage M, Wolters V, Roger P, Ineson P et al (1997) Soil function in a changing world: the role of invertebrates ecosystem ingineers. Eur J Soil Biol 33:159–93
Laycock WA, Richardson BZ (1975) Long-term effects of pocket gopher control on vegetation and soils of a subalpine grassland. J Range Manage 28:458–62
Legendre P, Legendre L (1998) Numerical ecology. Elsevier, Amsterdam, p 853
Litaor MI, Mancinelli R, Halfpenny JC (1996) The influence of pocket gopher on the status of nutrients in alpine soils. Geoderma 70:37–48
Malizia AI, Kittlein MJ, Busch C (2000) Influence of the subterranean herbivorous rodent Ctenomys talarum on vegetation and soil. Int J Mammlian Biol 65:172–82
Massei G, Genov P (2004) The environmental impact of wild boar. Galemys 16:135–45
Maynard DG, Kalra YP (1993) Nitrate and exchangeable ammonium nitrogen. In: Carter MR (ed) Soil sampling and methods of analysis. Lewis Publ, Boca Raton, pp 25–38, Can.Soc. Soil Sci
Mitchell J, Dorney W, Mayer R, McIlroy J (2007) Ecological impacts of feral pig diggings in north Queensland rainforests. Wildl Res 34:603–8
Mohr D, Cohnstaedt LW, Topp W (2005) Wild boar and red deer affect soil nutrients and soil biota in steep oak stands of the Eifel. Soil Biol Biochem 37:693–700
Montserrat P and Fillat F. The systems of grassland management in Spain. Manage Grasslands. 1991;37–70.
Moody A, Jones JA (2000) Soil response to canopy position and feral pig disturbance beneath Quercus agrifolia on Santa Cruz Island, California. Appl Soil Ecol 14:269–81
Moore DS, McCabe GP and Craig B. Introduction to the practice of statistics. W.H Freeman; (2008). pp. 100.
Olivier W and Reeder D. Sus Scrofa. In: IUCN Red list of threatened species Ed. IUCN. IUCN; 2008.
Page AL (1982) Methods of soil analysis. Part 2 chemical and microbiological properties. American Society of Agronomy, Madison, p 1159
Palacio S, Bueno CG, Azorín J, Maestro M and Gómez D. Wild boar disturbance increases nutrient and C stores of geophytes in subalpine grasslands. Am J Bot. in rev.
Pickett STA, White PS (1985) The ecology of natural disturbance and patch dynamics. Academic, Orlando, p 472
Pohl M, Alig D, Körner C, Rixen C (2009) Higher plant diversity enhances soil stability in disturbed alpine ecosystems. Plant Soil 324:91–102
Quinn GP, Keough MJ (2002) Experimental design and data analysis for biologist. Cambridge University Press, Cambridge, p 537
R Development Core Team. R: A Language and environment for statistical computing. In: R Foundation for Statistical Computing, Vienna, Austria; 2009
Schley L, Roper TJ (2003) Diet of wild boar Sus scrofa in Western Europe, with particular reference to consumption of agricultural crops. Mamm Rev 33:43–56
Sherrod SK, Seastedt TR (2001) Effects of the northern pocket gopher (Thomomys talpoides) on alpine soil characteristics, Niwot Ridge, CO. Biogeochemistry 55:195–218
Siemann E, Carrillo JA, Gabler CA, Zipp R, Rogers WE (2009) Experimental test of the impacts of feral hogs on forest dynamics and processes in the southeastern US. For Ecol Manage 258:546–53
Sims NKE. The ecological impacts of wild boar rooting in East Sussex. In: School of Biological Sciences. Sussex: University of Sussex; 2005. p. 268.
Singer FJ (1981) Wild pig population in the National Park. Environmetal Management 5:263–70
Singer FJ, Swank WT, Clebsch EEC (1984) Effects of wild pig rooting in a deciduous forest. J Wildl Manag 48:464–73
Stark JM, Firestone MK (1995) Mechanisms for soil moisture effects on activity of nitrifying bacteria. Appl Environ Microbiol 61:218–21
Tardiff SE, Standford JA (1998) Grizzly Bear digging: effect on subalpine meadow plants in relation to mineral nitrogen availability. Ecology 79:2219–28
Tierney TA, Cushman JH (2006) Temporal changes in native and exotic vegetation and soil characteristics following disturbances by feral pigs in a California grassland. Biol Invasions 8:1073–89
Vitousek PM (1985) Community turnover and ecosystem nutrient dynamics. In: Pickett STA, White PS (eds) The ecology of natural disturbance and patch dynamics. Academic, Orlando, pp 325–33
Vitousek PM (1986) Biological invasions and ecosystem properties: can species make a difference? In: Mooney HA, Drake JA (eds) Ecology of biological invasions of North America and Hawaii. Springer, New York, pp 163–76
Welander J (1995) Are wild boars a future threat to the Swedish flora? Ibex J Mt Ecol 3:165–7
Welander J. Spatial and temporal dynamic of a disturbance regime: wild boar (Sus scrofa L.) rooting and its effects on plant species diversity. In: Department of Conservation Biology. Uppsala: Swedish University of Agricultural Sciences; 2000.
Wirthner S, Frey B, Busse MD, Schütz M, Risch AC (2011) Effects of wild boar (Sus scrofa L.) rooting on the bacterial community structure in mixed-hardwood forest soils in Switzerland. European Journal of Soil Biology 47:296–302
Zuur AF, Ieno EN, Walker NJ, Saveliev AA and Smith GM. Mixed effects models and extensions in ecology with R. Springer. 2009:pp. 574.
Acknowledgments
We thank: Erin Cameron, Tan Bao, Samson Nyanumba and Isabel C. Barrio for their interesting suggestions on earlier versions of the manuscript. We gratefully acknowledge the support of the Spanish Science and Innovation Ministry (PN-MICINN) (CGL2008-00655/BOS) and the Spanish Environmental Ministry (MMAMRM, Organismo Autónomo de Parques Nacionales 002/2007). The Ordesa and Monte Perdido National Park (OMPNP) provided logistic support and free access to the National Park. We also want to thank Sergio (Burgos) and Silvia Chueca for field assistance, Isabel C. Barrio, María Bueno and Paula (Fañanás) for lab assistance and Santiago Pérez, Melchor Maestro and Ramón Galindo for assistance with the soil analyses. English was improved and revised by Bruce MacWhirter.
Author information
Authors and Affiliations
Corresponding author
Additional information
Responsible Editor: Zucong Cai.
Rights and permissions
About this article
Cite this article
Bueno, C.G., Azorín, J., Gómez-García, D. et al. Occurrence and intensity of wild boar disturbances, effects on the physical and chemical soil properties of alpine grasslands. Plant Soil 373, 243–256 (2013). https://doi.org/10.1007/s11104-013-1784-z
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11104-013-1784-z