
Abstract
Background and aims
This study aims to investigate the effect of nitrogen (N) on grain phosphorus (P) accumulation in japonica rice.
Methods
Six cultivars with contrasting agronomic traits were grown for 3 years (from 2008 to 2010) of field experiments under seven N treatments and 1 year (in 2010) of pot experiments with five N treatments to study the effect of N on grain phosphorus accumulation and to explore its physiological foundation.
Results
Grain total P and phytic acid concentration showed a clearly decreasing trend as N rate increased for both field and pot experiments. Pot experiment revealed that application of N increase plant biomass, but tended to lower plant P uptake, especially for the split topdressing treatments. Both harvest index (HI) and P harvest index (PHI) increased with N rate, but PHI was consistently higher than HI, indicating the larger proportion of P translocation to grain than that of dry matter by N. Further, ratio of PHI/HI differed significantly among genotypes, but was stable across contrasting N treatments.
Conclusions
The combination of decreased plant P uptake and dilution effect of increased grain yield by N is proposed as underlying mechanism of the decreased grain P concentration by high N.
Similar content being viewed by others

Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
Phosphorus (P) is critical in the metabolism of plants, playing a role in the transfer and storage of energy from photosynthesis and the metabolism of carbohydrates. It is also a structural component of the nucleic acids of genes and chromosomes and of many coenzymes, phosphoproteins and phospholipids. Application of P is recommend as a major agronomical practice for increasing crop yield, given the fact that about 5.7 billion hectares worldwide lack sufficient available P (Batjes 1997). P is a non-renewable resource and some commercially viable sources of rock phosphate are likely to be exhausted in the next century (Lott et al. 2011). Thus, strategies including breeding P efficient cultivars, optimizing P fertilizer rates, and minimizing soil P mining had been proposed for better P management of cropping systems (Ismail et al. 2007; Rose et al. 2010).
A large fraction of the P taken up by plants during vegetative growth is translocated to the developing seed. The phosphorus stored in mature seeds of grain and legume crops is estimated to represent a sum equivalent to more than 50 % of phosphate fertilizer applied annually worldwide (Lott et al. 2000). Consequently, seed phosphorus has value as a target in P management. Typically, inorganic P (Pi) consists of ~10 % of total phosphorus in cereal grains, whereas the majority of grain phosphorus, accounting for 50–80 %, occurs in the form of phytic acid (PA), a myo-inositol hexakisphosphate (Raboy 2007). The potential negative charges on the phosphate groups of PA bind various cations, especially K + and Mg2+, to form a salt called phytate (Raboy 2007). Pi is of higher bioavailability for humans and animals. By contrast, PA-P is indigestible for humans and non-ruminant animals such as poultry, swine, and fish, because they lack the enzyme to decompose phytic acid. In addition, phytic acid also forms complexes with proteins, digestive enzymes and minerals, and as such is considered to be an anti-nutritional factor (Lia et al. 2008; Raboy 2001). Substantial efforts have been made to identify mutants with impaired phytate biosynthesis, with low PA mutants being available now for key staple food crops such as barley, rice, and wheat. This development might offer potential benefits for the phosphorus nutrition of humans and animals (Raboy 2009).
Rice is one of the most consumed cereal crops, contributing a main source of energy, protein, and other nutrients to half of the world’s population (Frederick 2004). Rice production has to be improved sustainably to meet the requirement of this increasing population. For agronomical practices, nitrogen (N) fertilization is more efficient for obtaining high rice yields as compared to P and potassium (K) fertilization. In China, there is growing concern over the current agriculture paradigm of high usage of fertilizers mainly N. Overuse of N has not only caused low N use efficiency and soil acidification (Guo et al. 2010), but also resulted in the deleterious effect on rice quality especially the appearance quality (Qiao et al. 2011). Interest in rice quality has greatly increased in recent years because of the rapid economic development, which in return has increased the purchasing power and need for higher food quality. As a result, ways must to be found to reduce the detrimental influence on rice quality through elucidating the mechanisms of N effect. This will be critical in developing better N management for both quantity and quality in China’s rice production.
Our previous investigation of N influence on grain biochemical composition revealed a negative effect of N rate on phytic acid concentration in grains of 31 japonica cultivars (Ning et al. 2009). In the present study, six representative cultivars were subjected to 3 years (from 2008 to 2010) of field experiments with seven N treatments and 1 year (in 2010) of pot experiments with five N treatments. The main objectives are to study the effect of N on grain phosphorus and to explore its physiological foundation. We report the main results and discuss their significance for human nutrition, rice quality, and P use efficiency (PUE).
Materials and methods
Field experiments
Field experiments were performed at two different locations: Jiangning Experimental Station of Nanjing Agricultural University (31°56′39″N, 118°59′13″E) in 2008 and Danyang Experimental Station (31°54′31″N, 119°28′21″E) in 2009 and 2010. Randomized split-plot design with seven N treatments split for six japonica rice cultivars was used with four replications at Jiangning and three replications at Danyang. The soil type and fertility including total N, available P, and exchangeable K were described in our previous study (Qiao et al. 2011). Seven N treatments with three rates and two ratios of basal to topdressing were applied as illustrated by Ning et al. (2010): (1) CK, control of N fertilizer treatment, with no N being applied whereas P and K fertilizers being applied before transplanting; (2) L82 and L55, low N rate (90 kg/ha); (3) M82 and M55, moderate N rate (180 kg/ha); (4) H82 and H55, high N rate (270 kg/ha). Basal application was performed before transplanting, while N topdressing was conducted at panicle initiation stage. The fertilization modes of 82 and 55 indicate the ratios of basal/topdressing fertilizers are 8:2 and 5:5, respectively.
Seeding, transplanting, and heading dates of 2008 and 2009 were reported by Qiao et al. (2011). In 2010, cultivars were sown in seedbeds on May 25, transplanted to paddy field on June 28, and headed nearly on the same day as in 2009. In 2010, Zaofeng9 showed unnaturally lower filled grain percentage (~50 %), and thus Zaofeng9 and its counterpart Xudao4 were excluded and the remaining four cultivars were used. About 100 plants, with similar maturity, were harvested in each replication. The samples were naturally dried and dehulled. Brown rice was ground by a stainless steel grinder for three min and the resulting powders were used for chemical analysis.
For total P analysis, samples about 3.0 g were wet digested with 15 ml nitric/perchloric acid (3/1, V/V). Concentration of P in digests was measured spectrophotometrically with ammonium heptamolybdate-ammonium vanadate as chromogenic reagent. Phytic acid was measured by indirect method of iron precipitation, as reported by Ning et al. (2009). Phytic acid-P (PA-P) was calculated as multiplying phytic acid by 0.282, the P content in phytic acid.
Pot experiments
In 2010, pot experiments were conducted at Danyang to investigate changes in P accumulation and partitioning in response to N (Supplementary Fig. 1). The plants were allowed to grow under natural light in 25 L pots filled with 15 kg of soil from paddy field of Danyang. The soil is slightly alkaline (pH = 8.06), and contains 1.24 g/kg total N, 17.7 mg/kg available P, and 124.0 mg/kg exchangeable K. A basal application of N, P, and K as urea, calcium superphosphate, and potassium chloride was provided to each pot, with the rate equivalent to 90 kg/ha, 140 kg/ha, and 186 kg/ha in field conditions, respectively. Five N topdressing treatments were performed: (1) CK, control of N fertilizer treatment, with no N being applied whereas P and K fertilizers being applied before transplanting; (2) N9-0, low N rate, equivalent to 90 kg/ha in field conditions, with topdressing N being applied at panicle initiation (PI) stage; (3) N6-3, low N rate, topdressing N being split, 60 kg/ha at PI and 30 kg/ha at heading stage; (4) N18-0; and (5) N12-6. Each treatment had 30 pots.
Seedlings were transplanted on June 17, with three seedlings per pot. Similarly, Zaofeng9 showed unnaturally lower filled grain percentage, and thus it and its counterpart Xudao4 were excluded. About 30 plants were harvested at heading stage and maturity. The plants were then separated into stem (including sheath), leaf, and panicle (at heading, hull + rachis; at maturity, grain + hull + rachis). Samples were ground by a stainless steel grinder. Total P and phytic acid were measured as per field experiment. In addition, Pi in samples was extracted with 0.15 M tri-chloride acetic acid and measured colorimetrically.
Content of P in plant parts was calculated by multiplying the P concentration by dry weight, as shown in Supplementary table 1. The P harvest index (PHI) was calculated as the content of P in panicle divided by the total above ground P content (stem P +leaf P + panicle P).
Statistical analysis
Samples were analyzed in triplicate and mean values were used for comparisons. Variance analysis was performed with SPSS (Version 10.0) statistical software (Statistical Graphics Corp., Princeton, NJ). Means were compared by the least significant difference (LSD) test (P ≤ 0.05).
Results
Effect of N on grain total P, Pi and PA-P concentration
As reported previously, grain yield increased with N rate both in the 3-year field experiments (Ning et al. 2010; Qiao et al. 2011) and in the pot experiment (Supplementary Table 2). Variance analysis showed significant effect of N, cultivar, and their interaction on TP and PA-P concentration in grains produced from 3 years of field experiments, with cultivar having larger effect (Table 1).
Significant cultivar variations were detected among the cultivars examined. For example, Ningjing2 showed consistently higher concentrations of both TP and PA-P, whereas Wuyujing3 exhibited lower values for the 2 years of field experiments (Table 2). TP concentrations under field conditions were relatively higher, but they fell into the range as reported by Zeng et al. (2010).
N treatment had a negative effect on TP and PA-P, as revealed in both field and pot experiments (Tables 2 and 3). Generally, TP and PA-P concentration showed a clearly decreasing trend as N rate increased. This result is in agreement with our previous finding (Ning et al. 2009), further suggesting the depressing effect of N on P concentrations in japonica rice.
It should be noted Wuyunjing7, the high-yielding cultivar, showed a contrasting trend of grain total P , which was promoted, depressed, and unaffected by increased N rate in the field experiments of 2008, 2009, and 2010 (Table 2). This behavior was absent in pot experiments where Wuyunjing7 exhibited similar grain total P pattern as the other three cultivars (Table 3). This contradictory results may be partially attributed to the uniformity of experimental soil or the changes in growing environments especially the temperature and rainfall.
No significant differences existed between the two contrasting basal/topdressing treatments in field experiments. Similarly, concentrations of TP and PA-P were not significantly different between the two topdressing treatments in pot experiment (Tables 2 and 3). Thus, N rate has more influence on grain P accumulation than application pattern does.
PA-P constitutes the major part of grain P, with an average of 64.4 % (PA-P/Total P) across the five N treatments in the pot experiment (Table 3). Significant cultivar difference in ratio of PA-P/Total P was observed, with Ningjing2 having the highest whereas Ningjing1 being the lowest among the four cultivars. N rate showed no significant effect on this ratio, which is in agreement with former results (Ning et al. 2009). In addition, Pi concentrations were not significantly different among the five N treatments (Table 3).
Response of P uptake and partition to N treatments
To investigate the physiological foundation of the depressing effect of N on grain P concentrations, plants of pot experiments were sampled at heading stage and at maturity, and P in three kinds of sub-samples of stem (including sheath), leaf, and panicle were measured.
At heading stage, N did not show a clear effect on both stem and leaf P concentrations (Table 4), but had a promoting influence on P concentration in the hull and rachis. At maturity, a considerable amount of stem and leaf P was translocated to the grain, and N showed lowering effect for all the three kinds of subsamples (Table 4).
Total P content per plant at heading stage and maturity was calculated by multiplying the P concentration by dry weight (Supplementary Table 1). Results showed TP increased as N rate increased to 90 kg/ha (N9-0 and N6-3) and then decreased as N rate increased to 180 kg/ha (N18-0 and N12-6) for both stages. P uptake during grain filling stage was calculated by subtracting TP content per plant at heading stage from that at maturity. Typically, plant P uptake during grain filling stage consisted of 28.1 % of grain TP. Averaged across the four cultivars, plant P uptake tended to decrease with N rate, especially for the split fertilization treatments. However, cultivar difference was observed. As presented in Fig. 1, application of N led to promotion of P uptake during grain filling for Ningjing2, but had depressing effect in Ningjing1 and Wuyujing3, and no significant effect in Wuyujing7.

TP content per plant at heading and maturity stages as affected by N treatments. CK, control of N fertilizer treatment, no N was applied, whereas P and K fertilizers was applied before transplanting; N9-0, low N rate, equivalent to 90 kg/ha, with all topdressing N at panicle initiation stage; N6-3, low N rate, split topdressing, 60 kg/ha at PI and 30 kg/ha at heading stage; N18-0, 180 kg/ha, all at panicle initiation; N12-6, split topdressing of 180 kg/ha. TP-H: Total phosphorus in whole plant at heading (mg/plant); TP-M: Total phosphorus in whole plant at maturity (mg/plant); TP-U: Phosphorus uptake from heading to maturity (mg/plant)
Effect of N on P harvest index (PHI)
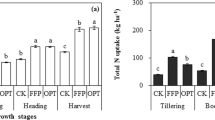
Harvest index (HI) varied significantly with cultivar. Wuyunjing7 had the highest HI of 54.3 % while Ningjing1 showed lowest value of 45.0 %. Cultivars with lower HI do not necessarily have lower P harvest index (PHI). For instance, Ningjing1 had the lowest HI but had highest PHI among the four cultivars.
Both HI and PHI increased with N rate, showing that N promotes the translocation of both dry matter and P from the vegetative organs to the grain. Moreover, PHI of the four cultivars under the five treatments was consistently higher than HI, indicating a larger proportion of P translocation to grain than that of dry matter.
The ratio of the PHI/HI, which describes the proportion of grain P compared to the proportion of grain carbon, was calculated and shown in Fig. 2. This ratio differed significantly among genotypes, ranging from 1.22 for Ninging1 to 1.31 for Wuyujing7. Moreover, PHI/HI of a given cultivar was stable across contrasting N treatments, which serves as further evidence that the ratio of PHI/HI can be used as a criterion in the breeding programs aiming for high P use efficiency (Rose et al. 2010).

Harvest index (HI), P harvest index (PHI), and ratio of PHI/HI of the four cultivars in response to N treatments
Discussion
Positive effect of N on grain P accumulation
P is one of the most limiting mineral nutrients for crop production in many low input agricultural systems. The use of genetically enhanced plants with improved P acquisition efficiency has been proposed as the most sustainable solution to increase crop yields in these systems (Ramaekers et al. 2010). However, as yield increases, the removal of all nutrients including P from soil will also increase (Rengel et al. 1999). Recently, a novel strategy has been proposed to lower ‘mining’ of soil P by reducing the amount of P in rice grains (Lott et al. 2000; Raboy 2001; Dorsch et al. 2003). Large genotypic variations in grain P concentration exists, suggesting the possibility of breeding rice with lower grain P concentration to reduce the depletion of P from rice field (Rose et al. 2010). In this context, one of the potential favorable effects of N on grain P accumulation may be associated with the decreased grain P concentration, which should be helpful for sustainable use of soil P by lowing the rate and degree of P mining.
Another possible beneficial effect of N on grain P accumulation may be that N application also improvePUE. As reported in both our previous (Ning et al. 2010; Qiao et al. 2011) and current study (Supplementary table 2), increased N rate produced higher grain yield at the same P fertilizer rate, thus causing higher agronomic efficiency (∆grain yield /∆ P fertilizer rate). On the other hand, the lower P concentration in both vegetative and reproductive organs meant higher ratio of ∆grain yield /∆ P uptake from soil. Thus physiological efficiency was also improved as N rate increased.
Phytic acid is of particular health concern, because of its contribution to nutritional deficiencies in around one-third of the world’s population, as well as phosphorus pollution in water (Lott et al. 2009). The third potential favorable effects of N on grain P accumulation may be associated with the decreased phytic acid concentration, which should have positive effect on human health. However, unlike wheat, maize, and barley, rice is consumed as milled grains. During polishing, outer layers where most of phytic acid is stored are discard, with little phytic acid remained in the resultant milled rice. This technical consequence is quite different from that of wheat milling, during which a considerable part of phytic acid still exists in the flour fractions due to incomplete separation of endosperm from the outer layer (Liu et al. 2008). The favorable effect of N on human nutrition needs to be further investigated.
Negative effect of N on grain P concentration
Phytate is stored form of phytic acid that chelated with K, and magnesium (Mg). The formation of phytate may be an important physiological process to trap inorganic metals, mainly P, K, and Mg from developing endosperm (Ogawa et al. 1979). Our previous work showed that N application can significantly increase rice yield through elevating panicle number per area and grain number per panicle (Ning et al. 2009). However, when the composition of grains was categorized in detail, a considerable part (about 25 %) of them was imperfect grains like green rice grain and malformed rice grain due to incomplete grain filling (Qiao et al. 2011). High N application increases the sink volume and promotes the intensity of starch and protein synthesis in grains, thereby increasing the requirement of P, K, and Mg. We also examined K and Mg concentrations in grains produced from pot experiments, and found a similarly depressing effect of N fertilization (Supplemental Table 3). We therefore speculate that the reduction in P accumulation may be partially associated with the high occurrence of imperfect rice under high N input. However, more work needs to be conducted to verify this hypothesis.
Mechanism of N effect on grain P accumulation
The reduction in grain P concentration in rice grains may be due to (1) the changes in P absorption from the soil, (2) the dilution effect because of increased grain yield, or (3) the changes in the translocation of P from vegetative organs to grains. Our result showed that N topdressing at both PI and heading stage resulted in increased plant TP concentration at the N rate of 90 kg/ha, whereas decreased plant TP concentration was evident at 180 kg/ha. Further, N showed a decreasing influence on plant P uptake during grain filling stage, especially for the split treatment (Fig. 1). Although the PHI was elevated by N and was higher than HI, the amount of translocated P from vegetative organs is not comparable to that of the biomass. Thus we suggest the combination of depressing effect on plant P uptake during grain filling and the dilution effect of grain yield to be the major mechanism of the decreased grain P by high N.
Singh and Modgal (1978) investigated the effect of N treatments on P uptake and translocation for rainfed upland indica rice, and found that total grain P concentration did not varied significantly with N rate, while total P uptake from soil increased significantly as N rate increased. In the current study with japonica rice in irrigated areas, a clear decrease of grain P concentration in response to increase N rate was observed. However, P uptake from the soil during the grain filling stage varied with cultivar, where Ningjing2 and Wuyunjing7 showed a positive response to N fertilization, while Ningjing1 and Wuyujing3 showed a reverse trend. The mechanism underlying the N effect on grain P concentration may therefore be genotype (indica and japonica rice) and cultivar dependent. Up to now, information is still lacking on the genotypic differences in uptake and translocation to grain. In addition, the soil bio-physical process of N-P interaction after N fertilization and its effect on P accumulation in grains are poorly understood. To evaluate ways of countering the deleterious effect of N on grain filling, for example lowering the ratio of imperfect grains, investigations on the efficacy of increasing P basal fertilization and foliar application are underway in our laboratory.
Abbreviations
- CK:
-
Control of N fertilizer treatment
- CV:
-
Coefficient of variation
- H:
-
High nitrogen rate
- L:
-
Low nitrogen rate
- M:
-
Moderate nitrogen rate
- N:
-
Nitrogen
- P:
-
Phosphorus
References
Batjes NH (1997) A world data set of derived soil properties by FAO-UNESCO soil unit for global modeling. Soil Use Manag 13:9–16
Dorsch JA, Cook A, Young KA, Anderson JM, Bauman AT, Volkmann CJ, Murthy PPN, Raboy V (2003) Seed phosphorus and inositol phosphate phenotype of barley low phytic acid genotypes. Phytochemistry 62:691–706
Frederick FF (2004) Rice proteins. In: Champagne ET (ed) Rice chemistry and technology, 3rd edn. American Association of Cereal Chemists, Inc, St. Paul, pp 143–162
Guo JH, Liu XJ, Zhang Y, Shen JL, Han WX, Zhang WF, Christie P, Goulding KWT, Vitousek PM, Zhang FS (2010) Significant acidification in major Chinese croplands. Science 327:1008–1010
Ismail AM, Heuer S, Thomsom M, Wissuwa M (2007) Genetic and genomic approaches to develop rice germplasm for problem soils. Plant Mol Biol 65:547–570
Lia CY, Park DS, Won SR, Hong SK, Ham JK, Choi JK, Rhee HI (2008) Food chemical properties of low-phytate rice cultivar, Sang-gol. J Cereal Sci 47:262–265
Liu ZH, Wang HY, Wang XE, Xu HY, Gao DR, Zhang GP, Chen PD, Liu DJ (2008) Effect of wheat pearling on flour phytase activity, phytic acid, iron, and zinc content. LWT Food Sci Technol 41:521–527
Lott JNA, Ockenden I, Raboy V, Batten GD (2000) Phytic acid and phosphorus in crop seeds and fruits: a global estimate. Seed Sci Res 10:11–33
Lott JNA, Bojarski M, Kolasa J, Batten GD, Campbell LC (2009) A review of the phosphorus content of dry cereal and legume crops of the world. Int J Agric Resour Gov Ecol 8:351–370
Lott JNA, Kolasa J, Batten GD, Campbell LC (2011) The critical role of phosphorus in world production of cereal grains and legume seeds. Food Secur 3:451–462
Ning HF, Liu ZH, Wang QS, Lin ZM, Chen SJ, Li GH, Wang SH, Ding YF (2009) Effect of nitrogen fertilizer application on grain phytic acid and protein concentrations in japonica rice and its variations with genotypes. J Cereal Sci 50:49–55
Ning HF, Qiao JF, Liu ZH, Lin ZM, Li GH, Wang QS, Wang SH, Ding YF (2010) Distribution of proteins and amino acids in milled and brown rice as affected by nitrogen fertilization and genotype. J Cereal Sci 52:90–95
Ogawa M, Tanaka K, Kasai Z (1979) Accumulation of phosphorus, magnesium and potassium in developing rice grains: followed by electron microprobe X-ray analysis focusing on the aleurone layer. Plant Cell Physiol 20:19–27
Qiao JF, Liu ZH, Deng SY, Ning HF, Yang XY, Lin ZM, Li GH, Wang QS, Wang SH, Ding YF (2011) Occurrence of perfect and imperfect grains of japonica rice as affected by nitrogen fertilizer. Plant Soil 349:191–202
Raboy V (2001) Seeds for a better future: ‘low phytate’ grains help to overcome malnutrition and reduce pollution. Trends Plant Sci 6:458–462
Raboy V (2007) Seed phosphorus and low-phytate crops. In: Turner BL, Richardson AE, Mullaney EJ (eds) Inositol phosphates: linking agriculture and environment. CAB International, Oxfordshire, pp 111–132
Raboy V (2009) Seed phosphate. In: Bennetzen JL, Hake SC (Eds) Handbook of maize: its biology. Springer, New York, pp 149–437
Ramaekers L, Remans R, Rao IM, Blair MW, Vanderleyden J (2010) Strategies for improving phosphorus acquisition efficiency of crop plants. Field Crop Res 117:169–176
Rengel Z, Batten GD, Crowley DE (1999) Agronomic approaches for improving the micronutrient density in edible portions of field crops. Field Crop Res 60:27–40
Rose TJ, Pariasca-Tanaka J, Rose MT, Fukuta Y, Wissuwa M (2010) Genotypic variation in grain phosphorus concentration, and opportunities to improve P-use efficiency in rice. Field Crop Res 119:154–160
Singh BK, Modgal SC (1978) Dry-matter production, phosphorus and potassium uptake as influenced by levels and methods of nitrogen application in rainfed upland rice. Plant Soil 50:691–701
Zeng Y, Zhang H, Wang L, Pu X, Du J, Yang S, Liu J (2010) Genotypic variation in element concentrations in brown rice from Yunnan landraces in China. Environ Geochem Health 32:165–177
Acknowledgments
This research was supported by grants from Program for New Century Excellent Talents in University (NCET-10-0472) and National Natural Science Foundation of China (30971733 and 31171485).
Author information
Authors and Affiliations
Corresponding author
Additional information
Responsible Editor: Matthias Wissuwa.
J. Bi and Z. Liu contributed equally to this work.
Rights and permissions
About this article
Cite this article
Bi, J., Liu, Z., Lin, Z. et al. Phosphorus accumulation in grains of japonica rice as affected by nitrogen fertilizer. Plant Soil 369, 231–240 (2013). https://doi.org/10.1007/s11104-012-1561-4
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11104-012-1561-4