
Abstract
Soils are frequently exposed to drying and wetting events and previous studies have shown that rewetting results in a strong but short-lived flush of microbial activity. The aim of this study was to determine the effect of the water content during the dry period on the size and duration of the flush and on the rate of recovery. Two soils (a sand and a sandy loam) were maintained at different water contents (WC) 30, 28 and 25 g water kg−1 soil (sand) and 130, 105 and 95 g water kg−1 soil (sandy loam) for 14 days, then rewet to the water content at which respiration was optimal [WC 35 (sand), WC200 (sandy loam)] and maintained at this level until day 68. Ground pea straw (C/N 26) was added and incorporated on day 1. The controls were maintained at the optimal water content throughout the 68 days. Respiration rates during the dry phase (days 1–14) decreased with decreasing water content. The flush of respiration after rewetting peaked on day 15 in the sandy loam and on day 16 in the sand; it was greatest in the soils that had been maintained at the lowest water content [WC25 (sand) and WC95 (sandy loam)]. Cumulative respiration during the remainder of the incubation period in which all soils were maintained at optimal water content increased more strongly in the soils that had been dry compared to the constantly moist control. On the final day of the dry period (day 14), cumulative respiration in the dry soils was 29–65% (sand) and 67–94% (sandy loam) of the constantly moist control whereas on day 68 it was 80–84% (sand) and 86–96% (sandy loam). The greater increase in cumulative respiration in the previously dry soils can be explained by the reduced decomposition rates during the dry period which resulted in higher substrate availability on day 14 compared to the constantly moist control. Microbial community structure assessed by phospholipid fatty acid analyses changed over time in all treatments but was less affected by water content than respiration; it differed only between the highest and the lowest water content. These differences were maintained throughout the incubation period in the sandy loam and transiently in the sand. It can be concluded that the soil water content during the dry phase affects the size of the flush in microbial activity upon rewetting and that microbial activity in previously dried soils may not be fully restored even after 54 days of moist incubation, suggesting that drying of soil can have a significant and long-lasting impact on microbial functioning.
Similar content being viewed by others

Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
Top soils often dry out during the summer, particularly in Mediterranean climates. The soils experience drying and rewetting events as the dry periods may be interrupted by occasional rainfall events.
As a soil dries, water is held in increasingly smaller pores and the water films around aggregates become thinner and disconnected (Ilstedt et al. 2000); decreasing water availability and increasing the energy required to withdraw water. Moreover, substrate and nutrient diffusion are restricted and microbes become substrate-limited (Stark and Firestone 1995). Water availability can be expressed as water potential, with low (more negative) potential indicating high energy requirement. The water potential is the sum of various potentials; in soils, the most important potentials are matric potential (a measure of how strongly the water is held onto soil surfaces) and osmotic potential (a function of the concentration of soluble salts in the soil solution).
Decreasing water content also affects the osmotic potential, because the salt concentration in the remaining soil solution increases as the water content decreases. Hence, microorganisms will also experience decreases in osmotic potential as the soil dries (Chowdhury et al. 2011).
Stressors, such as low water potential, impose a metabolic burden on the surviving microbes due to the need for stress tolerance mechanisms (Harris 1980; Oren 1999; Schimel et al. 2007). In response to low water potential, some microbes accumulate osmolytes which prevent the movement of water out of the cells (Oren 2001).
Rewetting dry soil induces a flush of respiration that usually occurs within a few hours after rewetting and is followed by a rapid decrease in respiration rates (Butterly et al. 2009; Franzluebbers et al. 1994; Kieft et al. 1987; Mikha et al. 2005). The flush of respiration upon rewetting has been explained by increased substrate availability due to release of the osmolytes accumulated during the dry phase, cell lysis and breakdown of aggregates which releases previously protected organic matter (Denef et al. 2001; Fierer and Schimel 2003; Halverson et al. 2000; Sparling et al. 1985). In a soil in which the size and activity of the microbial biomass was manipulated by the addition of different substrates, the size of the flush after rewetting was greatest in the treatment with the largest and most active biomass (Butterly et al. 2009). Drying and rewetting may also change microbial community composition (Butterly et al. 2009; Hamer et al. 2007; Schimel et al. 2007; Wilkinson et al. 2002; Williams 2007), which suggests that some microbial species or groups are more susceptible to drying and rewetting stress than others. For example, fast-growing organisms are more likely to die in dry soil and are more susceptible to rewetting than slow-growing microbes (Bottner 1985; Van Gestel et al. 1993). Thus, after drying and rewetting events, slower growing microbes may dominate (Bottner 1985; Cortez 1989). In a forest soil, rewetting of dry soil induced a shift towards gram-positive bacteria and an increase in bacteria/fungi ratio (Hamer et al. 2007). The effect of single and multiple drying and rewetting cycles on the size of the flush upon rewetting has been studied extensively (e.g., Butterly et al. (2009); Mikha et al. (2005); Van Gestel et al. (1993)), however in most studies, the water content of the soils during the dry phase was reduced to very low levels: air-dry or until the soils did not lose any more water at room temperature.
Little is known about the impact of the water content during the dry phase on the size of the flush upon rewetting and how long after rewetting it takes for microbial activity (measured by cumulative respiration) to return to the level of the constantly moist control.
Recovery after stress/disturbance is an important feature in ecosystem sustainability. Recovery can be partial or complete and the length of time required for recovery varies. For example, after soil disinfection, the capacity to decompose glucose and chitin was fully restored after 12 weeks (Wada and Toyota 2007). But after heating of soil to a temperature experienced during forest fire (200°C for 1 h), bacterial density and activity were less than the controls even after 15 weeks incubation (Diaz-Ravina et al. 1996). Bacterial community composition had recovered 56 d after gamma-irradiation, whereas the fungal community did not (McNamara et al. 2007). Substrate-induced respiration recovered within 6 days after rewetting of air-dry soil, but the rate of degradation of the fungicide metalaxyl-M and the insecticide lufenuron was still reduced even after 34 days (Pesaro et al. 2004). Microbial biomass C and N recovered completely after drying and rewetting within 24 d moist incubation in a soil with low soil organic matter content, but had recovered to only 50% of the moist control in a soil with high soil organic matter content (Hamer et al. 2007), which is in contrast to the study by De Nobili et al. (2006) who found that recovery after rewetting of dry soil was more rapid in soil with higher soil organic matter content. Orwin et al. (2006) reported that recovery after drying and rewetting was not correlated with resource availability (soil C, N, P).
The aim of this study was to determine the effect of soil water content during the dry period on the magnitude of the flush of respiration after rewetting and the time for recovery in two soils, a sand and a sandy loam. We tested the following hypotheses: (1) the flush in respiration will be greatest in the treatment with the lowest water content during the dry period; and (2) the time to recovery will be shortest and the extent of recovery greatest in the treatment with the highest water content during the dry period.
Materials and methods
Two soils from Monarto (35° 05′ S and 139° 06′ E) and Mount Bold (38° 11′ S and 138° 69′ E), South Australia differing in texture were used in the study: a sand (sand 91.3%, silt 5%, clay 3.7%, pH 7.7, EC1:5 150 μS/cm, N 0.09%, C 0.69%, water holding capacity 6.7%) and a sandy loam (sand 57.5%, silt 25%, clay 17.5%, pH 5.2, EC 1:5 68 μS/cm, N 0.33%, C 3.65%, water holding capacity 36.4%). After collection, the soils were air-dried and sieved to <2 mm.
The moisture retention curves of the soils were determined with suction and pressure techniques (Klute 1986) and given in Chowdhury et al. (2011).The osmotic potential at a given water content was estimated using the equation of Richard (1954).
Previous experiments had shown that maximal respiration occurred at matric potential −0.03 MPa in the sand (35 g water kg−1 soil) and −0.10 MPa in the sandy loam (200 g water kg−1 soil) (Chowdhury et al. 2011). The air-dry soils were pre-incubated at these optimal water contents for 10 days at 25°C before the experiment was begun. Ten days was chosen on the basis of several experiments with a range of soils (including the two used in the present study) which showed that respiration rates stabilized 7–10 days after rewetting air-dry soil (unpublished data). At the end of the pre-incubation, the soils were dried to the desired water content in a fan forced oven at 25°C. The selection of water contents was based on previous experiments (Chowdhury et al. 2011), ranging from optimal to that at which respiration was decreased by 30–50%. The sand was adjusted to 30, 28 and 25 g water kg−1 soil, the sandy loam to 130, 105 and 95 g water kg−1 soil. The control soils were maintained at the optimal water content (35 and 200 g water kg−1 soil for the sand and the sandy loam, respectively). In the following, the treatments are referred to as WC35, WC30, WC28 and WC25 (sand) and WC200, WC130, WC105 and WC95 (sandy loam).
After adjusting the water content, pea (Pisum sativum) straw (C/N 26, ground and sieved to 0.25–2 mm) was mixed into the soils (2% w/w) to provide a readily available nutrient source. The amended soils were then added into PVC cores and the cores were placed into glass jars (as described below) for respiration measurement (day 0).
Respiration was measured over 68 days. On day 14, water was added to the dried soils to bring the water content of all treatments to the optimal water content. This water content was maintained until day 68.
Analyses
Pre-incubated sand (40 g) or sandy loam (30 g), was added to PVC cores (diameter 3.7 cm, height 5 cm) with a nylon mesh base (0.75 μm, Australian Filter Specialist) and packed to a bulk density according to their texture (http://www.pedosphere.com/resources/bulkdensity/triangle_us.cfm): 1.55 g cm−3 (sand) and 1.46 g cm−3 (sandy loam). The cores were placed immediately into 1 L glass incubation jars and sealed with gas tight lids equipped with septa to allow headspace sampling. The glass jars were incubated in the dark at 22–25°C. Respiration was quantified by measuring headspace CO2 concentrations at regular intervals using a Servomex 1450 infra-red gas analyser (Servomex Group, Crowborough, England): daily in the first 20 days, every 3–4 days thereafter. After each measurement, the jars were opened to equilibrate the CO2 to ambient concentrations and then resealed. The CO2 evolved from each sample was calculated as the difference between the initial (immediately after resealing of the jars) concentration and that at the end of the measuring interval. The water content was maintained by weighing the cores and adding reverse osmosis water.
The percentage of C remaining of the added pea straw was calculated from cumulative respiration and the amount of C added on day 0. This calculation assumes that only the added pea residues are decomposed and that native SOM does not contribute to the respiration.
The microbial community structure was assessed on days 5, 14, 20 and 68 by phospholipid fatty acids (PLFA) analysis (based on (Frostegård et al. 1993)). Phospholipid fatty acids are components of cell membranes and rapidly dephosphorylated upon cell death; therefore PLFAs represent the living biomass (White 1995). The PLFA patterns provide a coarse measure of microbial community structure. Microbial groups such as bacteria and fungi, but not species or genotypes, differ in PLFA composition of their membranes; the so-called signature fatty acids can be used as a measure of abundance of microbial groups: Gram-positive bacteria i15:0 and i16:0; Gram-negative bacteria 16:1ω7c and 18:1ω7; actinomycetes 10ME-17:0 and 10ME-18:0, and fungi 18:2ω6, 18:1ω9 and 18:3ω6 (Kandeler 2007; Zak et al. 2000; Zelles et al. 1995).
Freeze-dried soil (4 g) was extracted with a one-phase solvent of chloroform, methanol and citrate buffer (1:2:0.8 v/v/v). The lipid-containing phase was collected and dried under a stream of nitrogen at 40°C. The PLFAs were separated from other fatty acids using silicic acid columns (Supelclean LC-Si-SPE Tubes, Supelco). The columns were washed sequentially with chloroform, acetone, and methanol, collecting the the methanol fraction which contains the PLFAs. After alkaline methanolysis the organic phase was collected in dichloromethane. An internal standard (methylnonadecanoate, 19:0) was added to each sample.
The fatty acid methyl esters were separated in a gas chromatograph with a flame ionization detector (GC-FID) (HP 6890) using an SP-2560 fused silica capillary column (75 m, 180 μm × 0.14 μm film thickness; Supelco, Sigma-Aldrich, Australia) with helium as carrier gas. The injector temperature was 250°C and the detector temperature was 260°C. The temperature program was as follows: after an initial temperature of 140°C, the temperature was ramped at 4°C/min to 240°C, and then held for 15 min.
The individual PLFA peaks were identified by comparing retention times with peaks of Supelco 37 standard mixture (Supelco, Bellefonte, PA) and peaks identified by GC-MS (gas chromatograph combined with mass selective detector HP 5973) using the same column and temperature program conditions and carrier gas as described above. Electron energy in electron impact was 70 eV. Mass spectrometer peak identification was based on comparison with the software library NIST02.L. The amounts of individual PLFAs are expressed in μg/g soil.
Statistical analysis
Significant differences between different treatments in a given soil over time in cumulative respiration, sum of PLFAs, sum of bacterial and fungal PLFAs were assessed by 2-way ANOVA and Tukey test with P ≤ 0.05. (GenStat® for Windows 8.0, VSN Int. Ltd, UK, 2005). Regressions between matric or water potential and relative cumulative respiration were calculated in MS Excel.
For statistical comparison of the microbial community structure, PLFA patterns, transformed as log (x + 1), were analysed by Primer E software (Primer-E Ltd, Plymouth Marine Laboratory, Plymouth, UK) and plotted using non-metric multi-dimensional scaling (MDS). The PLFA data were log (x + 1) transformed to balance the contributions of fatty acids by down-weighing the dominant fatty acids and increasing the weighting of rare fatty acids (Clarke and Warwick 2001). MDS plots with a 2D stress < 0.2 are considered to represent a good reflection of the overall structure of the communities. Significant differences in microbial community structure between treatments were determined by PERMANOVA (P ≤ 0.1).
Results
Potentials
During the first 14 days (dry period), the matric potential in the sand ranged from −0.16 to −0.44 MPa and the water potential from −1.04 to −1.62 MPa (Table 1). In the sandy loam the matric potential ranged from −0.10 to −1.30 MPa and the water potential from −0.19 to −1.56 MPa. The osmotic potential was higher in the sand than in the sandy loam, ranging from −0.88 to −1.18 MPa, compared to only −0.09 to −0.26 MPa in the sandy loam.
Respiration rates
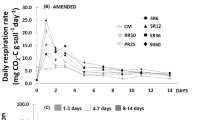
In the sand in the first 24 h after adjusting the soil water content and incorporating the pea straw (day 0), respiration rates decreased with decreasing water content (Fig. 1a). At the lowest water content (WC25) respiration rates were 72% lower than in the control WC35. On day 14, respiration rates had decreased in all treatments and were still lowest at WC25, although the difference between WC25 and WC35 was smaller than on day 1 (32%).

Respiration rates on days 1 to 68 in the sand (a) and the sandy loam (b) maintained at different water contents from day 1 to 14, rewet on day 14 and then maintained at optimal water content [WC35 (sand) and WC200 (sandy loam)] until day 68 (n = 4). Lines indicate standard error. Arrow indicates rewetting
Respiration rates in the sand peaked 2 days after adjusting all treatments to the optimal soil water content (day 16). On day 16, respiration rates were highest in WC25 and WC28, being more than 30% greater than in the constantly moist treatment WC35. After day 16 respiration rates declined in all treatments, with the decline being greatest in WC35. Compared to WC35, respiration rates at WC25 were 82% higher on day 20 and 62% higher on day 32. At the end of the experiment (day 68), respiration rates were low in all treatments.
Respiration rates in the sandy loam on day 1 were also lowest at the lowest water content (WC95), being 45% of that at the highest water content (WC200) (Fig. 1b). Respiration rates decreased from day 1 to day 14, but the decrease was greatest in WC200; thus on day 14, respiration rates at WC95 were 22% higher than in WC200. In contrast to the sand, respiration rates peaked 1 day after rewetting (day 15). On the first (day 15) and second day (day 16) after rewetting, respiration rates were more than two-fold higher in WC95 than in WC200. Respiration rates decreased over time, but, as in the sand, the decrease was greater in WC200. The relative difference between WC200 and WC95 decreased over time, with the respiration rates in WC95 being 70% higher on day 20 but only 10% higher on day 36. On day 68 respiration rates were low in all treatments.
Cumulative respiration
Cumulative respiration in the sand on day 1 decreased with water content; at WC25 it was only 28% of WC35 (Fig. 2a). The relative differences among the treatments remained the same during the 14 day dry period. On day 15 [1 day after adjusting to the optimal water content (35 g water kg−1 soil)], cumulative respiration decreased with water content; in WC25, it was only 28% of that at the highest water content (WC35). On the second day after rewetting, relative cumulative respiration in WC25 had increased, but was still only 36% of WC35. However by day 20, relative cumulative respiration in WC25 had increased to 51% of WC35. The relative cumulative respiration increased over time particularly in WC25, so that on day 68, all treatments that had been exposed to drying had reached 80% of the cumulative respiration of WC35.

Cumulative respiration [expressed in mg CO2-C g soil−1 from the onset of the respiration measurements (day 0) to the given day] on days 1, 14, 15, 16, 20, 32 and 68 in the sand (a) and the sandy loam (b) maintained at different water contents from day 1 to 14, rewet on day 14 and then maintained at optimal water content [WC35 (sand) and WC200 (sandy loam)] until day 68 (n = 4). Lines indicate standard error. Arrow indicates rewetting
Of the C added with the pea straw (assuming no decomposition of native SOM), only 70% remained on day 16 in WC35 whereas 89% remained in WC25. This difference became smaller over time, on day 36, 58% and 71% remained at WC35 and WC25, respectively. By the end of the experiment, the difference was even smaller, 49% and 59% remained at WC35 and WC25, respectively.
To assess respiration per unit biomass, cumulative respiration at given sampling time was divided by the sum of PLFAs at this date. Cumulative respiration per unit PLFA was always highest in WC35 (0.18 mg CO2-C μg PLFA on day 1), but the difference to the dried treatments decreased over time. On day 1 cumulative respiration per unit PLFA in WC25 was only 21% of that in WC35, whereas it was 76% on day 68.
Similar responses were found in the sandy loam: cumulative respiration on day 1 decreased with water content; at WC95, it was only 45% of that at WC200 (Fig. 2b). But unlike the sand, cumulative respiration in the sandy loam increased more strongly in the dry treatments (WC95, WC105 and WC130) than in WC200. Thus, the relative differences between the moist control and the drying treatments decreased during the 14 day dry period. On day 14, cumulative respiration in WC95 was 67% of WC200. The difference to WC200 continued to decrease during the moist incubation. By day 20, cumulative respiration in WC95 was 80% of WC200, and by day 68 it was 86%. In WC105 and WC130, cumulative respiration was less than 10% lower than in WC200.
Of the C added, 63% remained on day 14 in WC200 whereas 75% remained in WC95. Even more pronounced than in the sand, the differences between the moist and the dried soil decreased over time. On day 32, 53% of the added C had been decomposed in WC200, compared to 60% in WC95. On day 68, 36% remained in WC200 and 45% in WC95.
Cumulative respiration per unit PLFA was lower in the sandy loam than in the sand (0.04 mg CO2-C μg PLFA on day 1 in WC200), and, except for day 1, there were no significant differences in this parameter among the treatments (data not shown).
Correlation between cumulative respiration and matric or water potential
When analysed separately, cumulative respiration (in percentage of the constantly moist control) in the sand and the sandy loam was positively correlated with matric and water potential, i.e. cumulative respiration decreased with decreasing matric or water potential. However the relationship was strongest on day 1 (r 2 = 0.91 in both soils). On day 32, the correlation coefficient was 0.51 in the sand and 0.73 in the sandy loam. On day 68, there was no correlation between matric or water potential and cumulative respiration in the sand, whereas in the sandy loam cumulative respiration was correlated with both potentials (r 2 = 0.77). To assess if the relationship between relative cumulative respiration and potentials can be applied across both soils, the data of the two soils was combined. Relative cumulative respiration was weakly correlated with matric potential on day 1 but not on the other days (r 2 = 0.17) (Fig. 3). However, it was correlated with water potential at all sampling times (r 2 = 0.56–0.73) with no clear trend over time.

Relationship between cumulative respiration (% of constantly moist control) in the sand and the sandy loam and matric potential (a) and water potential (b) on days 1, 14 and 32 (n = 4).
Microbial community structure
On day 20 (6 days after rewetting), the sum of PLFAs was 4.3 (sand) and 15.0 μg g−1 soil (sandy loam). The abundance of bacterial fatty acids was 1.8 (sand) and 6.5 μg g−1 soil (sandy loam) and that of fungi was 1.9 (sand) and 3.8 μg g−1 soil (sandy loam). In both soils, the sum of total, bacterial and fungal PLFA did not change over time and there were no clear trends among the treatments (data not shown). The Permanova results of the PLFA patterns indicate that the microbial community structure differed between the highest and the lowest water content on days 14 and 20 in the sand and up to day 68 in the sandy loam (Fig. 4). In all treatments and both soils, microbial community structure on day 68 differed from that on day 14 .

Non-metric multi-dimensional scaling plots of the microbial community structure assessed by phospholipid fatty acid analysis on days 5, 14, 20 and 68 in the sand (a) and the sandy loam (b) maintained at different water contents from day 1 to 14, rewet on day 14 and then maintained at optimal water content [WC35 (sand) and WC200 (sandy loam)] until day 68. Symbols represent means of 3 replicates for a given water content and sampling day, numbers indicate sampling days. Symbols close to each other indicate similar community composition whereas communities represented by symbols that are far apart from another differ in composition
Discussion
This study showed that microbial activity is reduced by low water content and recovers after rewetting, but it remained below that of the moist control 54 days after rewetting. Whereas microbial activity during the dry phase and after rewetting was strongly affected by water content, microbial community structure differed only between the lowest and the highest water content.
Respiration during the dry phase and flush after rewetting
In the first 14 days, the decrease in respiration rates was greatest in the moist control which is probably due to the strong depletion of labile substrates in the first few days after residue addition compared to the soils with lower water content where the initial respiration rates were lower (Fig. 1). Thus, the smaller difference among treatments between respiration rates on day 14 compared to day 1 are due to the smaller decrease in respiration rates in the soils with lower water content and does not necessarily indicate adaptation of the microbes to low water content. Indeed, the relative difference in cumulative respiration between treatments was similar on days 1 and 14 in the sand indicating the lack of adaptation (Fig. 2). However in the sandy loam, the relative difference between the dry soils and the moist control was greater on day 1 (29–55% lower) than on day 14 (6–33% lower). This suggests that the microbes in the sandy loam were able to adapt to the lower water content.
In agreement with our first hypothesis, the magnitude of the rewetting flush was greatest in the soils with lowest water content (Fig. 1). This may be explained by the substrate availability upon rewetting which is likely to be greatest in the treatment with the lowest water content because: (i) microbes have accumulated more osmolytes during the dry period compared to the soils with the higher water content and these osmolytes are released/and or metabolised upon rewetting; and (ii) very dry aggregates may be more vulnerable to breakdown upon rewetting because the influx of water is greater due to the strong gradient in potential towards the centre of the aggregates.
The rewetting flush occurred on the first day after rewetting in the sandy loam but was delayed by 2 days in the sand (Fig. 1). At WC30 and 28 in the sand, this may be due to the lower water potential compared to WC130 and WC103 in the sandy loam (Table 1). However, although the WC25 in the sand and WC95 in the sandy loam had approximately same water potential (−1.62 and −1.56 MPa), the response to rewetting was delayed only in the sand. And this delay occurred although a higher percentage of the added C remained on day 14 in the sand (92%) than in the sandy loam (75%).
The delayed response to rewetting in the sand may be explained by three factors: (i) a microbial biomass which was 3–4 fold lower than in the sandy loam; this would not only limit the concentration of osmolytes released from the cells upon rewetting, but also the capacity of the surviving microbes to utilise the substrates immediately; (ii) fewer small pores than in the sandy loam, which may still contain water when the large pores are already drained, allowing microbes within them to survive in otherwise dry soil; and (iii) the lower osmotic potential which could have resulted not only in reduced water availability but also ion toxicities and element imbalances.
Recovery of respiration during moist incubation
Although respiration rates decreased over time during the moist incubation in all treatments, the decline was less in the previously dried soils (Fig. 1), presumably due to increased substrate availability as decomposition rates in the first 14 days were lower. On day 14, at least 10% more of the C added with the pea straw was still available at the lowest water content compared to the moist control. At the end of the experiment (day 68), after the previously dried soils had been incubated for 54 days at optimal water content, cumulative respiration had recovered to between 80 and 96% of the moist control in both soils, with the recovery being greater in the sandy loam (Fig. 2). By then, respiration rates were very low and similar in all treatments, therefore a full recovery of the dry treatments is unlikely even if the moist incubation were to be continued for longer.
The second hypothesis (i.e. the time to recovery will be shortest and the extent of recovery greatest in the treatment with the highest water content during the dry period) was based on the premise that a lower water stress during the dry period would allow microbes to recover more rapidly and to a greater extent than microbes exposed to more severe drying stress. However, this hypothesis is only supported by the cumulative respiration in the sandy loam where WC130 reached similar values as in the moist control after 20 days whereas cumulative respiration in WC95 was still 14% lower than the moist control on day 68 (Fig. 2). In the sand, cumulative respiration in all dried treatments was similar on day 68 (17–20% lower than the control) and the rate of increase in cumulative respiration was greater in WC25 (increasing from 32% of the control on day 15 to 80% of the control on day 68) than in WC30 (increasing from 67% of the control on day 15 to 84% of the control on day 68). This suggests that in the sand, the impact of water stress in all drying treatments was similar; indeed, water potential varied only between −1.25 and −1.62 MPa. In contrast, water potential in the sandy loam varied from −0.31 to −1.56 MPa. Despite the smaller differences in water potential between the drying treatments in the sand, the amount of C added remaining on day 14 in WC25 was 10% lower than in WC30, indicating that the microbes were less active during the dry period. A similar difference in substrate utilisation between WC95 and WC130 was recorded in the sandy loam. Thus, although the greater rate of recovery in WC25 can be attributed to the higher substrate availability, a stronger increase in cumulative respiration in WC95 should also have occurred in the sandy loam. However, whereas only 75% of the added C remained on day 14 in WC95 in the sandy loam, 92% remained in WC25 in the sand. This greater percentage of the C remaining from the added residues in the sand may have included a greater amount of easily available compounds which could lead to higher recovery rates in the sand.
Although differential substrate availability can help explain the results of the present experiment, the importance of substrate availability in recovery after drying and rewetting is controversial. Whereas De Nobili et al. (2006) found that recovery after rewetting of soil was more rapid in soil with higher soil organic matter content, the opposite was reported by Hamer et al. (2007) and in the study by Orwin et al. (2006) recovery after drying and rewetting was not correlated with resource availability (soil C, N, P).
Response of the microbial community structure to water content
Whereas microbial activity (respiration rates and cumulative respiration) differed among the treatments in both soils, the abundance of bacteria or fungi was not affected by differential extent of drying in either the sand or the sandy loam, and microbial community structure differed only between the highest and the lowest water content (Fig. 4). This suggests that, in contrast to microbial activity, community structure is less sensitive to water content. Moreover, differences in response to drying and subsequent recovery cannot be explained by differences in community structure. However, it should be noted that PLFA is only a relatively coarse measure of community structure because it does not provide information on species or genotype level. For this, DNA-based methods are required. It is possible that such methods could reveal changes in relative abundance of certain species, e.g. within bacteria, fungi or functional groups such as nitrifying bacteria.
In many previous studies, microbial communities in soils exposed to one or several drying and rewetting cycles differed from those in constantly moist soils (Butterly et al. 2009; Gordon et al. 2008; Hamer et al. 2007; Schmitt and Glaser 2011), but not always (Butterly et al. 2009; Griffiths et al. 2003). Of these studies, most used fatty acid analysis, only Griffiths et al. (2003) assessed bacterial community structure by DNA-based methods. In the studies that reported changes in community structure, the soils were dried to very low water contents—lower than in the present study. Therefore, the difference in community structure between the highest and the lowest water content here is in agreement with those previous studies. Interestingly, these differences in community structure between the highest and the lowest water content in the present study remained throughout the recovery period in the sandy loam although microbial community structure changed over time in all treatments (Fig. 4). Similarly, Pesaro et al. (2004) found that bacterial and archaeal community structure in dried and rewetted soil differed from that in the constantly moist control even after 34 d of moist incubation.
Relationship between cumulative respiration and water potential
Cumulative respiration was positively correlated with matric and water potential in both soils, however, the relationship became less strong over time after rewetting which also shows microbial activity recovered in the previously-dried soils (Fig. 3). When the data of both soils was combined, cumulative respiration was not related to matric potential, whereas it was positively correlated with water potential (matric + osmotic) throughout the experiment. The poor relationship to matric potential can be explained by the relatively low osmotic potential in the sand, particularly at low water contents. This shows that it is important to consider water potential and not just matric potential when comparing the effect of water stress on microbes in different soils.
Conclusions
This study showed that the flush in respiration after rewetting of dried soil is greatest in soils that had been exposed to the lowest water content which is most likely due to greater substrate availability upon rewetting. The results also suggest that, even after 54 days at optimal water content recovery after drying may not be complete, particularly in soils that have been exposed to very low water potentials. This indicates that drying of soil can have a significant and long-lasting impact on microbial function. Both extent of stress and recovery after rewetting is soil type dependent. In finer-textured soils, the fraction of microbes located in small pores may be exposed to less severe water stress than those in larger pores and can, therefore, recover more rapidly after rewetting. This needs to be verified using a greater number of soils.
This study also highlighted the importance of considering water potential rather than water content or matric potential when comparing water stress between soils.
In this study, the abundance of broad groups (e.g. bacteria or fungi) was little affected by the extent of drying, suggesting that the large differences in respiration were due to modulation of the activity per cell and not changes in community structure. However, it can not be ruled out that microbial community structure on a finer scale, e.g. genotypes, may change in response to drying or during the following moist period.
References
Bottner P (1985) Response of microbial biomass to alternate moist and dry conditions in a soil incubated with C-14-labeled and N-15-labelled plant material. Soil Biol Biochem 17:329–337
Butterly CR, Bunemann EK, McNeill AM, Baldock JA, Marschner P (2009) Carbon pulses but not phosphorus pulses are related to decreases in microbial biomass during repeated drying and rewetting of soils. Soil Biol Biochem 41:1406–1416
Chowdhury N, Marschner P, Burns R G (2011) Response of microbial activity and community structure to decreasing soil osmotic and matric potential. Plant Soil. in press.
Clarke KR, Warwick RM (2001) Change in marine communities: an approach to statistical analysis and interpretation. Primer-E, Plymouth
Cortez J (1989) Effect of drying and rewetting on mineralization and distribution of bacterial constituents in soil fractions. Biol Fertil Soils 7:142–151
De Nobili M, Contin M, Brookes PC (2006) Microbial biomass dynamics in recently air-dried and rewetted soils compared to others stored air-dry for up to 103 years. Soil Biol Biochem 38:2871–2881
Denef K, Six J, Bossuyt H, Frey SD, Elliott ET, Merckx R, Paustian K (2001) Influence of dry-wet cycles on the interrelationship between aggregate, particulate organic matter, and microbial community dynamics. Soil Biol Biochem 33:1599–1611
Diaz-Ravina M, Prieto A, Bååth E (1996) Bacterial activity in a forest soil after soil heating and organic amendments measured by the thymidine and leucine incorporation techniques. Soil Biol Biochem 28:419–426
Fierer N, Schimel JP (2003) A proposed mechanism for the pulse in carbon dioxide production commonly abserved following the rapid rewetting of a dry soil. Soil Sci Soc Am J 67:798–805
Franzluebbers K, Weaver RW, Juo ASR, Franzluebbers AJ (1994) Carbon and nitrogen mineralization from cowpea plant parts decomposing in moist and in repeatedly dried and wetted soil. Soil Biol Biochem 26:1379–1387
Frostegård A, Bååth E, Tunlid A (1993) Shifts in the strucutre of soil microbial communities in limed forests as revealed by phospholipid fatty acid analysis. Soil Biol Biochem 25:723–730
Gordon H, Haygarth PM, Bardgett RD (2008) Drying and rewetting effects on soil microbial community composition and nutrient leaching. Soil Biol Biochem 40:302–311
Griffiths RI, Whiteley AS, O’Donnell AG, Bailey MJ (2003) Physiological and community responses of established grassland bacterial populations to water stress. Appl Environ Microbiol 69:6961–6968
Halverson LJ, Jones TM, Firestone MK (2000) Release of intracellular solutes by four soil bacteria exposed to dilution stress. Soil Sci Soc Am J 64:1630–1637
Hamer U, Unger M, Makeschin F (2007) Impact of air-drying and rewetting on PLFA profiles of soil microbial communities. J Plant Nutrit Soil Sci 170:259–264
Harris R F (1980) Effect of water potential on microbial growth and activity. In Water potential relations in soil microbiology. pp 23–95. Soil Science Society America, Madison.
Ilstedt U, Nordgren A, Malmer A (2000) Optimum soil water for soil respiration before and after amendment with glucose in humid tropical acrisols and a boreal mor layer. Soil Biol Biochem 32:1594–1599
Kandeler E (2007) Physiological and biochemical methods for studying soil biota and their function. In Soil Microbiology, ecology, and biochemistry. Ed. E A Paul. pp 53–84. Elsevier
Kieft TL, Soroker E, Firestone MK (1987) Microbial biomass response to a rapid increase in water potential when dry soil is wetted. Soil Biol Biochem 19:119–126
Klute A (1986) Water retention: laboratory methods. In: Klute A (ed) Methods of soil analysis, Part 1. Soil Science Society of America, Madison, pp 635–660
McNamara NP, Griffiths RI, Tabouret A, Beresford NA, Bailey MJ, Whiteley AS (2007) The sensitivity of a forest soil microbial community to acute gamma-irradiation. Appl Soil Ecol 37:1–9
Mikha MM, Rice CW, Milliken GA (2005) Carbon and nitrogen mineralization as affected by drying and wetting cycles. Soil Biol Biochem 37:339–347
Oren A (1999) Bioenergetic aspects of halophilism. Microbiol Molec Biol Rev 63:334–348
Oren A (2001) The bioenergetic basis for the decrease in metabolic diversity at increasing salt concentrations: implication of the functioning of salt lake ecosystems. Hydrobiologia 466:61–72
Orwin KH, Wardle DA, Greenfield LG (2006) Context-dependent changes in the resistance and resilience of soil microbes to an experimental disturbrance for three primary plant chronosequences. Oikos 112:196–208
Pesaro M, Nicollier G, Zeyer J, Widmer F (2004) Impact of soil drying-rewetting stress microbial communities and activities and on degradation of two crop protection products. Appl Environ Microbiol 70:2577–2587
Richard LA (1954) Determination of the properties of saline and alkali soils. United States Department of Agriculture Handbook 60. Washington, DC, pp 7–53
Schimel JP, Balser TC, Wallenstein M (2007) Microbial stress response physiology and its implications for ecosystem function. Ecology 88:1386–1394
Schmitt A, Glaser B (2011) Organic matter dynamics in a temperate forest soil following enhanced drying. Soil Biol Biochem 43:478–489
Sparling GP, Whale KN, Ramsay AJ (1985) Quantifying the contribution from the soil microbial biomass to the extractable P levels of fresh and air-dried soils. Aust J Soil Res 23:613–621
Stark JM, Firestone MK (1995) Mechanisms for soil moisture effects on the activity of nitrifying bacteria. Appl Environ Microbiol 61:218–221
Van Gestel M, Merckx R, Vlassak K (1993) Microbial biomass responses to soil drying and rewetting: the fate of fast- and slow-growing microorganisms in soils from different climates. Soil Biol Biochem 25:109–123
Wada S, Toyota K (2007) Repeated applications of farmyard manure enhance resistance and resilience of soil biological functions against soil disinfection. Biol Fertil Soils 43:349–356
White DC (1995) Chemical ecology: possible linkage between macro- and microbial ecology. Oikos 74:177–184
Wilkinson SC, Anderson JM, Scardelis SP, Tisiafouli M, Taylor A, Wolters V (2002) PLFA profiles of microbial communities in decomposing conifer litter subject to moisture stress. Soil Biol Biochem 34:189–200
Williams MA (2007) Resonse of microbial communities to water stress in irrigated and drought-prone tallgrass prairie soils. Soil Biol Biochem 39:2750–2757
Zak DR, Pregnitzer KS, Curtis PS, Holmes WE (2000) Atmosperic CO2 and the composition and function of soil microbial communities. Ecol Appl 10:47–59
Zelles L, Rackwitz R, Bai QY, Beck T, Beese F (1995) Discrimination of microbial diversity by fatty acid profiles of phospholipids and lipopolysaccharides in differently cultivated soils. Plant Soil 170:115–122
Acknowledgments
This study was funded by the Australian Research Council. Nasrin Chowdhury received an Endeavour Australia postgraduate scholarship. Petra Marschner thanks Alan Robson for introducing her to mycorrhiza all these years ago, she has been fascinated by soil biology ever since.
Author information
Authors and Affiliations
Corresponding author
Additional information
Responsible Editor: Hans Lambers.
Rights and permissions
About this article
Cite this article
Chowdhury, N., Burns, R.G. & Marschner, P. Recovery of soil respiration after drying. Plant Soil 348, 269–279 (2011). https://doi.org/10.1007/s11104-011-0871-2
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11104-011-0871-2