
Abstract
Phosphorus (P) loss from land can impair surface water quality. Losses can occur from soil and plant components. While it is known that P losses increase with soil P concentration, it is not known how losses from pasture plants vary with soil P concentration or between different forages. We examined total P and filterable reactive P (FRP) in water extracts of plant shoots, used as a measure of potential P loss to surface runoff, in different forage species relative to soil P concentration in field trials and a glasshouse experiment. The mean total P concentration of 16 forage species in grazed field plots was greater (P < 0.01; LSD05 = 117 mg kg−1) in legumes (3,480 mg kg−1) than for grasses (3,210 mg kg−1). Total plant P concentrations of grasses and legumes increased with soil Mehlich-3 P concentrations in both glasshouse and field trials with concentrations close to 6,000 mg kg−1 in arrowleaf clover at 680 mg kg−1 Mehlich-3 soil P. FRP in water extracts of plant shoots increased relative to plant total P as soil Mehlich-3 P increased, with the greatest concentrations shown by crimson clover and arrowleaf clover. Analysis of water extracts of ryegrass and clover herbage from a field trial showed that while FRP was increasing, phytase-available-P decreased significantly from about 70% of filterable unreactive P at the lowest Mehlich-3 P concentrations, to close to zero at 200 mg kg−1 Mehlich-3 P. The wide variation, and enrichment of FRP in water extracts and total P with increasing Mehlich-3 P among species, indicates that cultivar and site selection and sward management provide a potential option to mitigate P loss to surface waters.
Similar content being viewed by others


Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
Phosphorus (P) is a key limiting nutrient in the eutrophication of many surface waters (Carpenter 2008; Conley et al. 2009; Schindler et al. 2008). Inputs of P to surface water via surface runoff can originate from soil (McDowell and Condron 2004), dung or manure (Smith et al. 2001), and / or fertiliser (Hart et al. 2004). However, it is also possible to leach up to 80% of P from the shoots of dried and freshly cut pasture and crop residues (Sharpley 1981; Timmons et al. 1970; Wendt and Corey 1980). McDowell et al. (2007) showed that by hydrologically separating topsoil from shoots with petrolatum, up to a quarter of total P lost in surface runoff for 30 days after grazing of a ryegrass-white clover pasture to a residual dry matter cover of 1,300 kg ha−1 was released from plant shoots. This P was previously held within vacuoles and exposed by ripping of shoots, or the decomposition of damaged material (McDowell et al. 2007). The remaining three-quarters of P lost in surface runoff came from topsoil either via dissolution or erosion of particles, and potentially, plant roots. However, Henry et al. (2009) found that compared to soil P, root architecture did not affect the P concentration in surface runoff. Mundy et al. (2003) added weight to the hypothesis that recently grazed material was a considerable source of P to runoff by showing about twice as much P was lost in outwash from flood irrigation of a pasture mown to 43 mm compared to uncut pasture.
Most pastures are maintained at or above a soil test P concentration considered optimal for near maximum dry-matter production. This generally means that foliage will contain a minimum of 2,000 mg P kg−1 dry weight, but plant P concentration varies greatly with species (Minson 1990). Typically, pasture mass and P concentration quickly increases with soil test P concentration to an asymptote, after which factors other than soil P availability limit yield (Edmeades et al. 2006; Saunders et al. 1987). Beyond the asymptote, luxury uptake of P by the plant may parallel soil test P concentration (Butler et al. 2007), but it is unknown if this would translate into potential P losses in surface runoff. Water extraction of plant and soil materials have enabled the concentration of plant P in surface runoff to be estimated (e.g., McDowell and Condron 2004; Sharpley 1981). We therefore hypothesize that the concentration of P potentially available to surface runoff (i.e., water extractable) from the plant increases with soil test P concentration.
New Zealand pastures traditionally rely on a mixture of white clover (Trifolium repens L.) and ryegrass (Lolium perenne L.), but research continues to develop new cultivars and species to provide better forage quality and productivity. A similar situation, albeit with largely warm season grasses, and often without legumes, exists in the southern U.S. Work has been done in the southern U.S., and to a lesser degree in New Zealand, that has shown the P concentration and degree of P uptake of grasses and legumes varies with species (Hart and Jessop 1983; Crush et al. 2008; Pederson et al. 2002). Establishing the variation in P concentration of modern pasture species and cultivars grown at a site with one soil test P concentration is the first step in selecting those that do not maintain excessive P concentrations in foliage that may be lost in surface runoff during and after grazing (Caradus et al. 1993; Easton et al. 1997). If low foliage P concentration (but not too low to impair animal nutrition), at low soil test P, can be coupled with the production of large amounts high quality feed, the potential for P loss in runoff may be decreased.
The primary aim of this paper was to determine the relative uptake of P among common and newly developed species/cultivars for forage grazing at one soil test P concentration. We then aimed to determine, for a few of these species, the relative difference in P extracted by water from shoot material with increasing soil test P concentration. Data from glasshouse and field trials (outlined in Table 1 and explained further in detail below) provide information to estimate the relative contribution of different forages to the loss of P in surface runoff and for exploring new pasture systems that minimise the potential for P loss under intensive grazing.
Materials and methods
Forage trial to establish the concentration of P according to species and cultivars
A forage trial of 16 grasses, herbs or legumes was established in autumn on a beef farm (annual rainfall c. 1,200 mm) near Matamata, Waikato, New Zealand. The species chosen were: High and Low endophyte ryegrass (Lolium perenne L), Hybrid tetraploid ryegrass (Lolium x hybridum Hausskn.), Italian ryegrass (Lolium multiflorum L.), Cocksfoot (Dactylis glomerata L.), Prairie grass (Bromus willdenowii Kunth.), Timothy (Phleum pratense L.), Tall fescue (Festuca arundinacea Schreb), two Red clovers—G27 and Pawera (Trifolium pratense L.), Lucerne (Medicago sativa L.), White clover (Trifolium repens L.), Sainfoin (Onobrychis viciifolia Scop.), Sulla (Hedysarum coronarium L.), Chicory (Chicorium intybus L.), and Sheeps burnet (Sanguisorba minor Scop.). Plots (7 m by 7 m) were randomised within twelve 8 by 8 grids, each housing four replicates of the 16 species/cultivars. This gave a total of 48 replicates per species/cultivar across a 3-ha paddock. The soil was a well drained Tirau silt loam (New Zealand classification: Typic Orthic Allophanic Soil [Hewitt 1998], similar to a Cryand in US taxonomy) with a Mehlich-3 P concentration of 185 ± 14 mg kg−1. No fertiliser was added to the plots during the trial which were grazed by cattle, deer or horses as part of a wider preference trial. Further details of the preference trial setup can be found in Hunt and Hay (1990). Although it cannot be discounted, the potential for uneven excretal returns or plant growth due to preferential grazing was minimised by sampling regrowth away from obvious dung patches and 3 weeks after sheep had grazed the plots to a common residual dry matter of 1,200 kg ha−1. Samples of shoot material were taken from each plot at monthly intervals for 9 months, dried at 60°C, ground (<1 mm) and total P determined on a bulked sample by a commercial laboratory.
Glasshouse trial of potential P loss from pasture to surface runoff
For the glasshouse trial, soil (c. 300 kg) was taken from the 10–20 cm depth of a site in Arkansas, U.S. that had been in long-term pasture and grazed for >20 years, except in 2008 when the field was used for Soybean and re-sown in pasture the following year. The soil, a Pembroke silt loam (Fine-silty, mixed, active, mesic Mollic Paleudalf), was air-dried, crushed and sieved to pass a 4 mm sieve. Soil was divided into pots, each with about 2 kg of soil and superphosphate (9 g kg−1 total P) added in a suspension to increase Mehlich-3 extractable soil P concentration by 0, 5, 10, 15, 40, 80, 160, 400, or 650 mg kg−1. Once added, soils were watered to field capacity (∼30% soil water content by weight) and left to air dry for 1 week before being wet again. This process was repeated to ensure superphosphate had equilibrated with soil (Sharpley et al. 1984). After a period of 8 weeks, the Mehlich-3 extractable soil P (Mehlich 1984) concentrations achieved were 20, 23, 30, 44, 62, 97, 187, 428, and 679 mg kg−1.
Soil for 5 replicates of each soil P concentration was transferred to 2 L pots (with drainage holes) to achieve a soil bulk density of 1.1 g cm−3. The pots were watered with 0.5 L of deionised water and plants sown at about 30 seeds per pot, equivalent to a sowing rate of 20 to 25 kg ha−1. The plant species chosen were those common in grazed pastures in Arkansas (Arrowleaf clover [Trifolium vesiculosum Savi]; Bermuda grass [Cynodon dactylon L.]; Crimson clover [Trifolium incarnatum L.]; and Tall fescue [Festuca arundinacea, Schreb.]). Pots were watered daily with sufficient deionised water to remain moist until germination and then maintained at 80% of field capacity (c. 50% available soil moisture; asm). After 2 weeks, 200 mL of ¼ strength P-deficient Hoagland solution was provided weekly (i.e., 1 mM Ca(NO3)2, 1.5 mM KNO3, and 0.5 mM MgSO4) (Hoagland and Arnon 1950). Plants were culled to 20 per pot 1 month after germination, in an effort to leave an even space between plants and prevent competition decreasing yields.
After 8 weeks growth, plants were harvested by cutting stems 3 cm above the soil surface. A preliminary experiment was conducted first to determine if fine particles adhered to pasture plants via water splash, insects, dust or detritus would affect or interfere with the concentration of P detected as available for loss in surface runoff. This involved the analysis of filterable reactive P (FRP) in water extracts (see below) of a washed sub-sample of foliage from each pot compared with an unwashed sub-sample. Washing involved spraying deionised water onto plant shoots as a mist and brushing shoots dry with a cloth before harvest. The data indicated that FRP in water extracts was greater (P < 0.05) in unwashed samples than in washed samples (Fig. 1).

Relationship between filterable reactive P in water extracts of pasture shoots before and after washing for Arrowleaf clover, Bermuda grass, Crimson clover and Fescue
The water extraction procedure has been used to determine the potential contribution of ungrazed pasture soil to P losses in surface runoff (McDowell and Condron 2004). However, subsequent work has adapted this test for forage and shown that the determination of FRP in water extracts of plant shoots can predict the potential for P loss from recently (up to 30 days) grazed plant shoot material (McDowell et al. 2007, 2010a). Although it cannot be discounted that pre-washing removes a proportion of plant-derived FRP potentially lost in surface runoff, we present WSP data from samples that had been washed and brushed dry before harvest to remove any potential interference associated with water splash, insects, dust or detritus.
Briefly, the water extraction procedure for plant shoots involved first hand plucking and cutting harvested shoot material into 1-cm long lengths until 5 g of shoot material was obtained. The cut shoots were then added to 100 mL of deionised water and shaken end-over-end for 10 mins. Filterable reactive P was determined by filtering the extract through a 0.45 μm syringe filter and determining P colorimetrically via the method of Watanabe and Olsen (1965). A sub-sample of shoot material was dried at 60°C and dry matter determined. The dried sample was ground (<1 mm) and total P determined by a commercial laboratory. The harvest and extraction of plant material was repeated every 6 weeks until 5 datasets were obtained.
Field trial examination of the potential P loss from pasture to surface runoff
To compliment the glasshouse trial, plant shoot samples were taken from three field trials. The first trial (Woodlands) examined P uptake and the potential for P in surface runoff from ryegrass (Lolium perenne L.) and white clover (Trifolium repens L.) monocultures at different rates of Mehlich-3 extractable soil P. The second and third trials (Winchmore irrigation and fertiliser) examined the uptake and release of P from long-term plots of ryegrass and white clover sown as a mixed pasture.
The Woodlands trial was at the Woodlands Research Farm, 15 km east of Invercargill, Southland, New Zealand on a Waikiwi silt loam (New Zealand classification: Firm Brown Soil, similar to a Typic Dystrochrept in US taxonomy). Annual rainfall is 1,000–1,300 mm. The paddock (3 ha) chosen for the trial was flat (<1% slope) and had been in pasture grazed by sheep for at least 20 year. Mehlich-3 extractable soil P concentration was between 130 and 150 mg kg−1 for the previous 5 years. The paddock was sprayed with glyphosate in August, 2008 and again a month later. In mid November 2008, plots, each 2-m wide by 6-m long, were established with a 1-m gap between them with six replicates of the following treatments:
-
Monocultures of either white clover (Trifolium repens L.) or ryegrass (Lolium perenne L.) at sowing rates of 8 kg ha−1 and 24 kg ha−1, respectively.
-
Establishment via direct drill or cultivation to 20-cm depth followed by sowing clover by hand and the use of a direct drill for ryegrass (including on cultivated plots).
-
Application of 10, 35, or 100 kg P as superphosphate ha−1.
This gave a total of 12 treatments and 72 plots arranged in a randomised design. The use of different rates of superphosphate, coupled with the different methods of establishment (i.e. with or without cultivation) gave a wide range of Mehlich-3 extractable soil P concentrations. After establishment, plots were grazed whenever dry matter was estimated to exceed 3,000 kg ha−1. Ten months after plots were established, grab samples (c. 50 g wet weight) were taken of pasture shoots in spring when plants had reached about 10–15 cm height. Soil samples were also taken from the 0–7.5 cm layer, dried, crushed and sieved <2 mm before determination of Mehlich-3 extractable soil P. Plant samples were washed, dried, and analysed for FRP in water extracts and total P as per the glasshouse experiment.
The Winchmore field trials were located at the Winchmore Irrigation Research Station located in Canterbury, New Zealand. Both long-term trials were irrigated and called the “irrigation” and “fertiliser” trial respectively due to a focus on different irrigation rates at the same P fertiliser rate, and on different P fertiliser additions under the same irrigation regime, respectively. The station has an annual rainfall of 740 mm and the soil type is a Lismore stony silt loam formed from moderately weathered greywacke loess (New Zealand classification: Brown Soil [Hewitt 1998], similar to a Typic Dystrochrept in US taxonomy).
The irrigation trial had five treatments: dryland (no irrigation); irrigation applied every 21 days (3w); irrigation applied when topsoil (0–100 mm) moisture reached 10, 15, or 20% (designated as 10%, 15%, and 20% irrigated, respectively). These corresponded to 0% asm (wilting point), 25% asm, and 50% asm (McBride 1994). The 3w, 10%, 15%, and 20% irrigated treatments received on average 6.5, 2.6, 4.2, and 7.7 irrigations per year, respectively, 100 mm at a time (Rickard and McBride 1986). The irrigation trial consisted of 0.09 ha bays, bordered so that each was 9-m wide × 100-m long. There were four replicates of the dryland treatment and five replicates of the irrigated treatments. All treatments received 250 kg ha−1 of single superphosphate (SSP) in winter annually (c. 25 kg P ha−1). Except for winter when no grazing occurred, each treatment was rotationally grazed by a separate flock of sheep with 6, 10, 14, and 18 stock unitsFootnote 1 ha−1 for the dryland, 10%, 15%, 20%, and 3w treatments, respectively.
The fertiliser trial had five treatments, each replicated in 0.09-ha bays, four times: nil (no P applied); 188 kg SSP ha−1 yr−1 (18–19 kg P ha−1) since 1952; 250 kg SSP ha−1 yr−1 (23 kg P ha−1) since 1981; and 376 kg SSP ha−1 yr−1 since 1981. Treatments were grazed by separate flocks of sheep (6, 11, 17, and 17 stock units ha−1 for the nil, 188SSP, 250SSP, and 376SSP treatments, respectively) to prevent carry-over of dung P between treatments. No grazing occurred in winter. Irrigation (100 mm) occurred whenever soil moisture content (w w−1) dropped below 15% (0–100 mm depth). On-average, this occurred 4.3 times per year.
Grab samples of pasture (c. 50 g wet weight) were hand plucked from areas within each plot that had been excluded from grazing by stock for 30 days (approximate height 8–20 cm) to minimise any potential influence from urine or dung deposits. Soil samples were also taken from the 0–7.5 cm layer, dried, crushed and sieved <2 mm before analysis of Mehlich-3 extractable soil P. The plant samples were analysed for FRP in water extracts and total P as per the glasshouse experiment.
Enzyme assay
Investigation of organic P fractions in water extracts was based on a modified Turner et al. (2002) method using alkaline phosphomonoesterase (EC 3.1.3.2.), phosphodiesterase (EC 3.1.4.1.), and phytase (EC 3.1.3.8). The Woodlands water extracts were chosen for study as they represented pure monocultures in a field setting. Analysis was conducted on a set of 16 ryegrass or clover plots, each set covered a wide range of Mehlich-3 P concentrations. Before enzyme assays were done, total filterable P in water extracts was determined via persulphate digestion (Patton and Kryskalla 2003) and yielded by difference with FRP, the filterable unreactive P (FURP, largely organic; Turner et al. 2002) fraction. Briefly, for the assays, 0.1 M sodium azide (to prevent microbial growth and P uptake) and the appropriate buffered enzyme mixture (see Turner et al. 2002) were added to a sample of water extract in the ratio of 1:1:9, and incubated overnight at 37°C in inert plastic centrifuge tubes. A blank sample containing no enzyme was also incubated to account for any organic P hydrolysis that may have occurred during incubation. The next morning, FRP was determined in both samples. The difference in FRP concentration was denoted as enzyme liberated P. Calibration curves for P determination also contained each enzyme to account for any P release by the enzyme itself or protein interference. For determination of phosphodiesters, alkaline phosphomonoesterase was also added to fully convert esters to orthophosphate detectable by colorimetry. The concentration of diesters was then calculated as FRP released from the combined alkaline phosphomonoesterase and diesterase mix minus P released from alkaline phosphomonoesterase alone.
Results
Total phosphorus uptake in relation to species, cultivar and soil P concentration
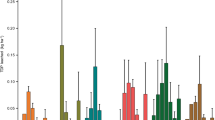
The field trial of 16 different species and cultivars revealed a significant difference (P < 0.01; LSD05 = 324 mg kg−1) in mean total P concentrations between species/cultivars (Fig. 2). The least mean total P occurred in Prairie grass (2,928 mg kg−1), while the greatest occurred in Lucerne (3,770 mg kg−1). Collectively, legumes (inclusive of herbs) had a greater (P < 0.01; LSD05 = 118 mg kg−1) mean total P concentration (3,514 mg kg−1) than grasses (3,236 mg kg−1). This difference agrees with studies of forage harvested on a monthly basis in New Zealand by Reay and Waugh (1983) and Crush et al. (1989) and on an annual basis in the U.S. by Minson (1990) and Pederson et al. (2002). However, the difference between legume and grass total P contrasts to the mean concentration of 2,200 mg kg−1 for legumes and 3,600 mg kg−1 for grasses presented by the U.S. Department of Agriculture—Natural Resources Conservation Service (1992).

Variation in mean total shoot P concentration by species or cultivar (grey grass, white legume/herb) on the beef farm. Error bars represent the standard error of the mean (n = 48). Comparison of means are made via Tukey’s multiple comparison test. Means with the same letter (a or b) are not significant
Concentrations of total P in plant shoots of the glasshouse trial varied from a low of c. 1,000 mg kg−1 at a Mehlich-3 P concentration of c. 25 mg kg−1 to a high of c. 6000 mg kg−1 at a Mehlich-3 extractable soil P concentration of c. 680 mg kg−1 (Fig. 3). In general, total P concentrations in the shoots of white clover and ryegrass were greater (P < 0.01) than those detected in the shoots of the glasshouse forage at similar Mehlich-3 extractable soil P concentrations. However, in general, and commonly reported in the literature (e.g., Minson 1990), total P concentrations tended to increase with soil P availability, as measured by Mehlich-3 extractable soil P concentration.

Variation in plant total P concentrations with soil Mehlich-3 P concentration for the glasshouse trial and in the field in the short-term Woodlands and long-term Winchmore trials. Concentrations for species grown in the glasshouse trial are means established at the end of the trial after 30 weeks. Error bars are not given for clarity
Variation of FRP in water extracts of shoots with soil P and species
The concentration of FRP in water extracts of plant shoots studied increased with Mehlich-3 extractable soil P concentration (Fig. 4a, b). The relationship between FRP in water extracts and Mehlich-3 extractable soil P concentration was best described by a power function (P < 0.05 or better) in both the glasshouse and field trials. Further analysis showed that FRP concentration in water extracts increased, relative to plant total P concentration (i.e. as a quotient), with increasing Mehlich-3 extractable soil P concentration (Fig. 4c, d).

Variation in FRP in water extracts of shoots (left) or FRP in water extracts of shoots as a percentage of total P in shoots (right) amongst species grown at different Mehlich-3 extractable soil P concentrations in the glasshouse trial (top) and in the field in the short-term Woodlands and long-term Winchmore trials (expansion shown in the bottom graph). Error bars represent the standard error of the mean for harvests from each replicate over the duration of the experiment. The lines show the fit of a power function or linear regression at the P < 0.05 (*), <0.01 (**), or <0.001 (***) level of significance
Among species in the glasshouse trial, both clovers (Arrowleaf and Crimson) had a greater FRP concentration in water extracts compared to the two grasses (Bermuda and Tall Fescue) at similar Mehlich-3 extractable soil P concentrations (P < 0.05). The same occurred in the Woodlands field trial (P < 0.05). This data indicated that storage of P in water extractable form, potentially available for loss in surface runoff, is greater on a dry-weight basis for the legumes studied than for the grasses. However, the true runoff potential will be determined by the relative yields of species and whether or not legumes are sown as part of a mixed pasture (Brink et al. 2001).
Analysis of water extracts with phosphatase enzymes classified FURP into phytase-, diesterase- and monoesterase-available P. On average, FURP comprised 14% of total filterable P extracted by water. The percentage of each enzyme class is presented in Fig. 5 as a proportion of FURP for the Woodlands white clover and ryegrass monocultures where the soil Mehlich-3 extractable soil P concentration was considered at least optimal for pasture growth (>40–50 mg kg−1; Beegle et al. 1998; Roberts and Morton 1993). Often, the sum total of phytase- diesterase- and monoesterase-available P was marginally greater than 100% (up to 106%) of FURP due to some activity by enzymes in more than one class (Turner et al. 2002). The data showed a significant decrease (P < 0.05) in the percentage of FURP as phytase-available P with increasing Mehlich-3 extractable soil P concentration (Fig. 5c). The sum of the remaining two classes would, by difference, increase as a result, but individually showed no significant trend (Fig. 5a, b).

Relationship between monoesterase-, diesterase- or phytase-available P, as a proportion of filterable unreactive P (FURP), and Mehlich-3 extractable soil P for the Woodlands white clover and ryegrass monocultures. The fit of the exponential equation is significant at P < 0.05
Discussion
Plant P uptake
There are several factors that control the concentration of P in the pasture plants such as climate, management, or the ability of different species to scavenge P from the soil. In general, those species with a large root mass, and a greater number of finer roots to explore small soil aggregates, will compete better than those with a poor root mass or root morphology (Richardson et al. 2009). At low soil test P concentrations, the fibrous root system of grasses concentrated in topsoil, where P is usually enriched, favours uptake of P compared to the tap root system of many legumes (McLaughlin et al. 2005). However, at higher soil test P concentrations well beyond an agronomic optimum for pastures (e.g. >50 Mehlich-3 P kg−1 soil; Beegle et al. 1998), such as that used here, the enhanced ability of grasses to scavenge P may be negated, allowing legumes to support greater total P concentrations in harvested material. It is important to determine how this translates to the potential for P losses in runoff over a range of soil test P (here determined as Mehlich-3 extractable soil P) concentrations, especially as the potential for P loss to surface runoff increases much more if soils are enriched with P beyond an agronomic optimum (Beegle et al. 1998).
At the lower range of Mehlich-3 extractable soil P, total P concentrations measured in the Woodlands ryegrass and white clover monocultures tended to be greater than total P concentrations measured in either the glasshouse trial or the Winchmore long-term trials. In addition to factors such as soil fertility or type and forage species (as noted above), plant total P concentration is known to vary with maturity, nitrogen (N) applications and climate (Marschner 1986). As a plant ages, total P concentration decreases, on average, about 0.03 g kg−1 dry matter day−1 (Whitehead 1966). However, any difference in total P concentration due to age is probably small as plants in both the glasshouse and Woodlands field trial were young and had similar time between harvests or grazing and measurement of FRP in water extracts. When plants are grown as monocultures, the application of N tends to decrease shoot P concentrations (Rosero et al. 1980). The application of N fertiliser to mixed ryegrass/clover pasture has also been shown to decrease herbage P content of the ryegrass component (Mouat et al. 1987). This may explain the lower total P concentration of the ryegrass which received 150 kg N ha−1 during the trial compared to white clover which received no N. However, as the glasshouse trial received only a small amount of N via a ¼ strength Hoagland solution, N application does not explain why the total P concentrations of both Woodlands monocultures were greater at low Mehlich-3 extractable soil P concentration. Finally, plant total P concentrations are known to vary with season, being greatest during periods of rapid growth, especially in spring (Saunders and Metson 1971; Metson and Saunders 1978). The Woodland monocultures were sampled in spring and therefore may have contained a higher proportion of young (high P) leaves than samples from the glasshouse trial.
Variation in FRP in water extracts of plant shoots
Past studies of nutrient uptake have ascribed curvilinear relationships, similar to that seen in Figs. 3 and 4, to increasing root density and the overlapping of depletion zones of individual root hairs decreasing P uptake (Itoh and Barber 1983). However, expressing FRP in water extracts as a proportion of total P indicated that more P was potentially available for loss in surface runoff as Mehlich-3 P increased. Past work on P forms, but not on the potential for loss to surface runoff, would tend to support this finding. For example, at suboptimal to optimal P supply, as measured by soil test P concentration, the concentration of P fractions such as diesters (e.g., lipids, nucleic acids), monoesters (e.g., adenosine triphosphate), and inorganic P increases, whereas beyond the optimal soil test P concentration, inorganic P increases (Hart and Jessop 1982). Comparing white clover and lotus, Hart and Jessop (1983) found that white clover preferentially stored P as inorganic P at the expense of lipid-P and ester-P and continued to do so with increasing soil test P concentration. The additional P is stored in highly vacuolated tissues (Kakie 1969; Marschner 1986). The exceptions to this rule are grains and seeds which may store excess P in monoester form, specifically phytate (Michael et al. 1980).
In addition to changes in FRP in water extracts with changing Mehlich-3 extractable soil P concentration, variation was also evident in P fractions within the 10–25% (average 14%) of total filterable P in water extracts that was FURP. Work by Turner et al. (2002) and others has suggested that diesters and monoesters may be available to algae, whereas phytate-P is probably not. The decrease in phytase-available P and subsequent increase in FRP, which is highly algal available, and the sum of diester- and monoester-P, suggests that algal available P increases at a greater rate beyond, compared to below soil test P concentrations optimal for pasture growth.
Management considerations
The power function describing the increase in FRP in water extracts with Mehlich-3 extractable soil P concentration also indicates that most change in FRP in water extracts, as with total P, occurs at low soil test P concentration (Fig. 4). However, much work has shown that the potential for P loss from cropping and pasture soils increases with soil test P concentration (e.g., McDowell and Sharpley 2001). Many have shown that this increase is exponential or occurs at a greater linear rate beyond a threshold in soil test P concentration (Koopmans et al. 2002). If an aim is to keep soil test P concentration within the agronomic optimum, but still decrease P losses, then the focus may need to shift to plant P losses.
Often the soil test P concentration for near maximum yield reflects the P requirements of the least P-efficient species in a mixed sward. For example, to reach within 97% of near maximum yield, white clover (Trifolium repens) commonly requires an Olsen P concentration at least 25 mg L−1, whereas ryegrass (Lolium perenne) may only require an Olsen P concentration <15 mg L−1 (Gillingham et al. 2008). There is potential to decrease P losses to surface runoff if plant species are chosen that avoid excessive P concentrations in foliage. Work of Hart et al. (1981) indicates that among legumes grown at a similar soil test P concentration, white clover stores greater quantities of P in foliage. Further decreases in P losses, and savings in P fertiliser, could be achieved if this species reaches the near maximum yield at low soil test P concentrations.
An alternative strategy has been to sow grass and legumes in a paddock or field as monocultures in a ratio of about 60% legume to 40% grass. This strategy has been shown to improve the liveweight gain of lambs and milk solids production of dairy cows by about 10% compared to a mixed sward (Cosgrove et al. 2003, 2006). Given the different P requirements of grasses and legumes, it may be possible to place the low-P grasses in areas of the catchment that exhibit a low soil test P concentration but produce the most runoff, while sowing the legume in “safe” (i.e., low runoff producing) areas. McDowell et al. (2010b) showed that by introducing conventional cultivation for ryegrass establishment in this system, soil P was decreased by redistributing it within the plough layer—thereby decreasing the potential for P loss from the soil by about 20–40%.
A caveat to this idea is that while the spatial separation of monocultures to account for hydrology (e.g. runoff) could decrease P loss, the design of such pasture systems needs to balance environment with agronomic performance. For instance, while runoff losses from tall fescue may be less than from a hybrid ryegrass at the same soil test P concentration (Fig. 2), recent research has indicated that the soil P requirement of tall fescue is much greater than originally thought (e.g. Miles and Manson 1995), and greater than ryegrass (Mackay et al. 2010). Furthermore, while maintaining low soil test P concentrations, especially in runoff-producing areas, places emphasis on selecting P-efficient pasture species, the quantity of P taken up and stored in shoots should be no more than required for animal nutrition. The P requirements of cattle vary for a number of reasons such as the breed of cattle (McDowell 1996), how available P is in the feed, and the effects of disease or parasites (Ternmouth 1990). Karn (2001) reviewed the P requirements of grazing cattle and indicated that a response to P supplementation only occurred when total P in the forage was <10–20 g kg−1. Although the recommended P concentration for lactating dairy cattle is greater (30 g kg−1; National Research Council 2001), this recommendation incorporates a considerable safety margin. Environmental scientists (e.g., Wu et al. 2000) contend that this P is not efficiently utilised and the excess P, being excreted in the faeces, causes a P loss problem when deposited either as dung or manure in a runoff producing area. Clearly, there is room to design a pasture system that would better match feed requirements of the animal and decrease the potential for environmental impact.
Conclusions
Findings generated by this combined glasshouse, plot, and long-term field research into P uptake and potential for loss to surface runoff by forages, will inform improved pasture management strategies and decrease the impact of P on surface water quality. At the same Mehlich-3 extractable soil P concentration (∼185 mg kg−1), the above ground biomass of legumes tended to maintain a greater total P concentration than exhibited by a range of grasses. In a pot trial, as Mehlich-3 P concentration increased to nearly 700 mg kg−1, P uptake by the forages increased, particularly for crimson clover. When forages were cut to simulate grazing, FRP in water extracts increased as a function of soil P and plant P concentration. This variation in FRP with species and soil P indicates a potential to manage forages and their diversity. One way would be to avoid sowing species that have a high potential to lose P to runoff in pastures that regularly produce runoff such as areas adjacent to streams. Conversely, high-P forage species in areas less hydrologically active and prone to runoff, can be managed to provide P requirements of grazing animals.
As P runoff and related water quality issues continue, forage species management in grazed pastures becomes another mitigation option available to farmers and land managers. While careful management of pastures will be needed to achieve these water quality goals, cultivar selection, site selection and sward management will provide low cost mitigation options.
Notes
One stock unit is equivalent to one ewe at 54 kg live-weight.
References
Beegle DB, Sharpley AN, Graetz D (1998) Interpreting soil test phosphorus for environmental purposes. In: Sims JT (ed) Soil testing for phosphorus: environmental uses and implications. Soil Coop Ser Bull No 389, Univ Delaware, Newark, DE, pp 31–40
Brink GE, Pederson GA, Sistani KR, Fairbrother TE (2001) Uptake of selected nutrients by temperate grasses and legumes. Agron J 93:887–890
Butler TJ, Muir JP, Provin TL (2007) Phosphorus fertilization of annual ryegrass and comparison of soil phosphorus extracts. J Plant Nutr 30:9–20
Caradus JR, Hay MJM, MacKay AD, Thomas VJ, Dunlop J, Lambert MG, Hart AL, van den Bosch J, Wewala S (1993) Variation within white clover (Trifolium repens L.) for phenotype plasticity of morphological and yield related characters, induced by phosphorus supply. New Phytol 123:175–184
Carpenter SR (2008) Phosphorus control is critical to mitigating eutrophication. Proc Natl Acad Sci USA 105:11039–11040
Conley DJ, Paerl WH, Howarth RW, Boesch DF, Seitzinger SP, Havens KE, Lancelot C, Likens GE (2009) Controlling eutrophication: nitrogen and phosphorus. Sci 323:1014–1015
Cosgrove GP, Hyslop MG, Anderson CB, Litherland AJ, Lambert MG (2003) Integrating novel forage management into sheep farm systems. Proc N Z Grassl Assoc 65:75–81
Cosgrove GP, Burke JL, Death AF, Lane GA, Fraser K, Pachero D (2006) The effect of clover-rich diets on cows in mid-lactation: production, behaviour and nutrient use. Proc N Z Grassl Assoc 68:267–274
Crush JR, Evans JPM, Cosgrove GP (1989) Chemical composition of ryegrass (Lolium perenne L.) and prarie grass (Bromus willdenowii Kunth) pastures. N Zeal J Agric Res 2:461–468
Crush JR, Boulesteix-Coutilier ARL, Ouyang L (2008) Phosphate uptake by white clover (Trifolium repens L.) genotypes with contrasting root morphology. NZ J Agric Res 51:279–285
Easton HS, MacKay AD, Lee J (1997) Genetic variation for macro- and micro-nutrient concentration in perennial ryegrass (Lolium perenne L.). Aust J Agric Res 48:657–666
Edmeades DC, Metherell AK, Waller JE, Roberts AHC, Morton JD (2006) Defining the relationships between pasture production and soil P and the development of a dynamic P model for New Zealand pastures: a review of recent developments. NZ J Agric Res 49:207–222
Gillingham AG, Morton JD, Gray MH (2008) Pasture responses to phosphorus and nitrogen fertilisers in east coast hill country: 2. Clover and grass production from easy slopes. NZ J Agric Res 51:85–97
Hart AL, Jessop D (1982) Concentration of total, inorganic, and lipid phosphorus in leaves of white clover and stylosanthes. NZ J Agric Res 25:69–76
Hart AL, Jessop D (1983) Phosphorus fractions in trifoliate leaves of white clover and lotus at various levels of phosphorus supply. NZ J Agric Res 26:357–361
Hart AL, Halligan G, Haslemore RM (1981) Analysis of the response of pasture legumes to phosphorus in a controlled environment. NZ J Agric Res 24:197–201
Hart MR, Quin BF, Nguyen LM (2004) Phosphorus runoff from agricultural land and direct fertilizer effects: a review. J Environ Qual 33:1954–1972
Henry A, Kleinman PJA, Lynch JP (2009) Phosphorus runoff from a phosphorus deficient soil under common bean (Phaseolus vulgaris L.) and soybean (Glycine max L.) genotypes with contrasting root architecture. Plant Soil 317:1–16
Hewitt AE (1998) New Zealand soil classification. Landcare research science series 1. Manaaki Whenua, Lincoln
Hoagland DR, Arnon DI (1950) The water-culture method for growing plants without soil. California Agricultural Experimental Station Circular 347, the College of Agriculture, University of California-Berkeley, CA
Hunt WF, Hay JM (1990) A photographic technique for assessing the pasture species performance of grazing animals. Proc N Z Grassl Assoc 51:191–196
Itoh S, Barber SA (1983) A umerical solution of whole plant nutrient uptake for soil-root systems with root hairs. Plant Soil 70:403–413
Kakie T (1969) Phosphorus fractions in tobacco plants as affected by phosphate application. Soil Sci Plant Nutr 15:81–85
Karn JF (2001) Phosphorus nutrition of grazing cattle: a review. Anim Feed Sci Technol 89:133–153
Koopmans GF, McDowell RW, Chardon WJ, Oenema O, Dolfing J (2002) Soil phosphorus quantity—intensity relationships to predict increased phosphorus loss in overland and subsurface flow. Chemosphere 48:679–687
Mackay AD, Gillingham A, Smith C, Budding P, Phillips P, Clarke-Hill W, Johnstone P (2010) Evaluation of the effects of grass species, irrigation, nitrogen fertiliser application and soil compaction on the response of modern dairy pastures to phosphorus fertiliser. Proc N Z Grassl Assoc 72:153–158
Marschner H (1986) Mineral nutrition of higher plants. Academic, London
McBride SD (1994) Pasture yield responses to irrigation in Canterbury. Proc N Z Grassl Assoc 56:165–168
McDowell LR (1996) Feeding minerals to cattle on pasture. Anim Feed Sci Technol 60:247–271
McDowell RW, Condron LM (2004) Estimating phosphorus loss from New Zealand grassland soils. NZ J Agric Res 47:137–145
McDowell RW, Sharpley AN (2001) Approximating phosphorus release from soils to surface runoff and subsurface drainage. J Environ Qual 30:508–520
McDowell RW, Nash DM, Robertson F (2007) Sources of phosphorus lost from a grazed pasture receiving simulated rainfall. J Environ Qual 36:1281–1288
McDowell RW, Sharpley AN, Simmons T (2010a) The potential for different plants to lose phosphorus in surface runoff. 4th International Symposium on Phosphorus Dynamics in the Soil-Plant Continuum. 19–23 September, 2010, Beijing, China. CD-ROM
McDowell RW, Knowler K, Cosgrove GP, Littlejohn R (2010b) The potential for a best management practice to decrease phosphorus loss from pastures while improving profitability. Proc N Z Grassl Assoc 72:171–176
McLaughlin MR, Sistani KR, Fairbrther TE, Rowe DE (2005) Overseeding common Bermudagrass with cool-season annuals to increase yield and nitrogen and phosphorus uptake in a hay field fertilized with swine effluent. Agron J 97:487–493
Mehlich A (1984) Mehlich-3 soil test extractant: a modification of Mehlich-2 extractant. Comm Soil Sci Plant Anal 15:1409–1416
Metson AJ, Saunders WMH (1978) Seasonal variations in chemical composition of pasture I. Calcium. Magnesium, potassium, sodium, and phosphorus. NZ J Agric Res 21:341–355
Michael B, Zink F, Lantzsch HJ (1980) Effect of phosphate application on phytin-P and other phosphate fractions in developing wheat grains. Z Pflanzenernaehr Bodenkd 143:369–376
Miles N, Manson AD (1995) Effects of soil acidity and phosphorus on the yield and chemical composition of tall fescue. Comm Soil Sci Plant Anal 26:843–860
Minson DJ (1990) Forage in ruminant nutrition. Academic, San Diego
Mouat MCH, Hay MJM, Crush JR, Nes P, Dunlop J, Hart AL (1987) Effects of nitrogen and phosphorus applications on pasture growth and composition on a recent alluvial soil with low phosphorus retention. NZ J Exp Agric 15:143–146
Mundy GN, Nexhip KJ, Austin NR, Collins MD (2003) The influence of cutting and grazing on phosphorus and nitrogen in irrigation runoff from perennial pasture. Aust J Soil Res 41:675–685
National Research Council (2001) Nutrient requirements of dairy cattle. 7th rev. edn. Natl Acad Sci, Washington, DC
Patton CJ, Kryskalla JR (2003) Methods of analysis by the U.S. Geological Survey National Water Quality Laboratory: evaluation of alkaline persulfate digestion as an alternative to Kjeldahl digestion for the determination of total and dissolved nitrogen and phosphorus in water. U.S. Geological Survey, Water Resources Investigations Report 03-4174. U.S. Geological Survey, Branch of Information Services, Federal Center, Denver, CO
Pederson GA, Brink GE, Fairbrother TE (2002) Nutrient uptake in plant species of sixteen forages fertilized with poultry litter: nitrogen, phosphorus, potassium, copper and zinc. Agron J 94:895–904
Reay PF, Waugh C (1983) Element composition of ryegrass and white clover leafblades—seasonal variation in a continuously stocked pasture. NZ J Agric Res 26:341–348
Richardson AE, Hocking PJ, Simpson RJ, George TS (2009) Plant mechanisms to optimise access to soil phosphorus. Crop Past Sci 60:124–143
Rickard DS, McBride SD (1986) Irrigated and non-irrigated pasture production at Winchmore, 1960 to 1986. MAF Technical Report no. 21, Winchmore Irrigation Research Station. Ministry of Agriculture and Forestry, Wellington, New Zealand
Roberts AHC, Morton JD (1993) Fertiliser use on New Zealand dairy farms. New Zealand Fertiliser Manufacturers’ Association, Auckland
Rosero OR, Tucker RE, Mitchell GE, Schelling GT (1980) Mineral utilization in sheep fed spring forages of different species, maturity and nitrogen fertility. J Anim Sci 50:128–136
Saunders WMH, Metson AJ (1971) Seasonal variation of phosphorus in soil and pasture. NZ J Agric Res 14:307–328
Saunders WMH, Sherrell CG, Gravett IM (1987) A new approach to the interpretation of soil tests for phosphate response by grazed pasture. NZ J Agric Res 30:67–77
Schindler DW, Hecky RE, Findlay DL, Stainton MP, Parker BR, Paterson MJ, Beaty KG, Lyng M, Kasian SEM (2008) Eutrophication of lakes cannot be controlled by reducing nitrogen inputs: Results of a 37-year whole-ecosystem experiment. Proc Natl Acad Sci 105:11254–11258
Sharpley AN (1981) The contribution of phosphorus leached from crop canopy to losses in surface runoff. J Environ Qual 10:160–165
Sharpley AN, Gray C, Jones CA, Cole CV (1984) A simplified soil and plant phosphorus model. II. Prediction of labile, organic and sorbed P. Soil Sci Soc Am J 48:805–809
Smith KA, Jackson DR, Withers PJA (2001) Nutrient losses by surface run-off following the application of organic manures to arable land. 2. Phosphorus. Environ Pollut 112:53–60
Ternmouth JH (1990) Phosphorus and beef production in northern Australia. 3. Phosphorus in cattle: a review. Trop Grassl 24:159–169
Timmons DR, Holt RF, Latterell JJ (1970) Leaching of crop residues as a source of nutrients in surface runoff water. Water Resour Res 6:1367–1375
Turner BL, McKelvie ID, Haygarth PM (2002) Characterisation of water-extractable soil organic phosphorus by phosphatase hydrolysis. Soil Biol Biochem 34:27–35
U.S.Department of Agriculture—Natural Resources Conservation Service (1992) Agricultural waste management field handbook. Natl. Eng. Handb. Part 651. NRCS, Washington, DC http://www.wsi.nrcs.usda.gov/products/W2Q/AWM/handbk.html. Accessed 5 May 2010
Watanabe FS, Olsen SR (1965) Test of an ascorbic acid method for determining phosphorus in water and NaHCO3 extracts from soil. Soil Sci Soc Am Proc 29:677–678
Wendt RC, Corey RB (1980) Phosphorus variations in surface runoff from agricultural lands as a function of land use. J Environ Qual 9:130–136
Whitehead DC (1966) Data on the mineral composition of grassland herbage from the Grassland Research Institute, Hurley, and the Welsh Plant Breeding Station, Aberystwyth, Technical Report 4, Grassland Research Institute
Wu Z, Satter LD, Sojo R (2000) Milk production, reproductive performance, and fecal excretion of phosphorus by dairy cows fed three amounts of phosphorus. J Dairy Sci 83:1028–1041
Acknowledgements
W.F. Hunt and R.J.M. Hay, Department of Scientific and Industrial Research, Grasslands Division, for management of the pasture species preference trial at Matamata. Funding for the remainder of this project was sourced from the International Science and Technology Linkages Fund administered by the New Zealand Foundation for Research, Science and Technology (FRST) and FRST project C10X1006.
Author information
Authors and Affiliations
Corresponding author
Additional information
Responsible Editor: Tim Simon George.
Rights and permissions
About this article
Cite this article
McDowell, R.W., Sharpley, A.N., Crush, J.R. et al. Phosphorus in pasture plants: potential implications for phosphorus loss in surface runoff. Plant Soil 345, 23–35 (2011). https://doi.org/10.1007/s11104-010-0687-5
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11104-010-0687-5