
Abstract
This study assessed the ability of biofertilizer inoculants containing Rhizobium leguminosarum bv. trifolii to enhance production of rice (Oryza sativa L.) under actual agricultural conditions in the Nile delta. Large-scale field experiments evaluated 5 rice varieties inoculated with 7 endophytic rhizobial strains during 5 growing seasons, including at sites ranked as the world’s highest in rice production. Inoculation with single strains or multi-strain consortia significantly increased grain yield in 19 of the 24 trials. By combining superior rhizobial inoculants with agricultural extension training, grain yield increased up to 47% in farmers’ fields, with an average increase of 19.5%. Data on rice straw production, harvest index and the agronomic fertilizer N-use efficiency also indicated positive agronomic benefits of rhizobial inoculation. These results establish the merit of deploying our biofertilization strategy using selected rhizobial strains to promote rice production capacity while reducing the need for additional chemical N-fertilizer inputs to maintain agricultural sustainability and acceptable production economy. Technology transfer of this important translational research can significantly help to alleviate hunger and meet the nutritional needs of many people in developing countries.
Similar content being viewed by others


Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
On June 19, 2009, the Food and Agriculture Organization of the United Nations (FAO) announced that “more than ever before, one sixth of humanity is undernourished as world hunger reaches a historic high of 1,020 million people going hungry every day in 2009” (http://www.fao.org/english/dg/2009/index.html). The scope of this study addresses how biofertilizer inoculants developed by agricultural microbiologists can help to remedy this ominous situation on global hunger and malnutrition.
Rice (Oryza sativa) is considered the world’s most important food crop, representing the staple diet for two-thirds of the human population. In response to current reduced availability and soaring prices of rice and other agricultural products, an urgent Emergency Rice Initiative was launched to assist African rice-growing countries in exploring sustainable approaches for seed production and distribution as well as for mineral fertilizer procurement and distribution (IRRI 2008). The goals of that initiative are to: 1) promote an agronomic revolution in rice production and reduce existing yield gaps, 2) accelerate the delivery of new post-harvest technologies, 3) accelerate the introduction of higher yielding rice varieties, 4) strengthen and upgrade the rice breeding and research pipelines, 5) accelerate research on the thousands of rice varieties and disseminate the results so scientists can access the vast reservoir of useful information they contain, and 6) educate a new generation of rice scientists and researchers for the public and private sectors.
An important issue raised by this initiative is the question of whether biofertilization technology can be adequately exploited to augment soil fertility and improve production economics while reducing chemical fertilizer inputs needed to achieve high crop yields, thereby alleviating some of the environmental and economic concerns associated with that agricultural practice. Biofertilization is still under appreciated even after more than a century since discovery that certain N2-fixing microbes can significantly benefit crop plants and be developed into biofertilizer inoculants on an industrial scale.
Our research relates to several goals of that hunger-alleviating initiative based on our success in development of new biofertilization strategies that increase rice production with less chemical fertilizer application (Dazzo and Yanni 2006). This practice is fully compatible with sustainable agriculture, environmental soundness, and enhanced production economy over the long-term worldwide. In earlier work, we described a natural endophytic association between rice and Rhizobium leguminosarum bv. trifolii (the bacterial N2-fixing root-nodule endosymbiont of clovers), the numerical abundance and diversity of local rhizobia strains that colonize endophytic niches within rice roots, the primary infection process in this Rhizobium-rice association followed by dissemination of the bacterial partner within the below-ground and above-ground host plant interior, its various plant growth-promotion responses and their probable underlying mechanisms, the nutritional impacts of the association on rice growth physiology and on grain for human consumption, the strain/variety specificity of the beneficial rhizobia-cereal association including first evidence of its inheritability, and three preliminary biofertilizer inoculation trials of its potential utilization under small-scale experimental field conditions (Biswas et al. 2000a; Biswas et al. 2000b; Chi et al. 2005; Perrine et al. 2001; Prayitno et al. 1999; Reddy et al. 1997; Yanni et al. 1997; Yanni et al. 2001).
These and other independent confirmations of worldwide, natural endophytic rhizobia-cereal associations and their benefit to the plant’s growth physiology (Chaintreuil et al. 2000; Dazzo et al. 2000; Gutierrez-Zamora and Martinez-Romero 2001; Hilali et al. 2001; Jha et al. 2009 ; Lupway et al. 2004; Mano and Morisaki 2008; Matiru and Dakora 2004; Mishra et al. 2008; Peng et al. 2008; Singh et al. 2006; Singh et al. 2009; Sun et al. 2008; Tan et al. 2001) have not lead to a technology that could be utilized directly by farmers, as positive plant-growth benefits obtained in lab, greenhouse and small experimental field plots still need meaningful “translational” research to prove the efficacy of biofertilizer inoculants under a variety of agricultural conditions, and successfully transfer those technologies to everyday use. The objective of this study was to conduct the translational research needed to assess the merit of applying this agricultural biotechnology in a large-scale rhizobial biofertilization program for rice under actual farming conditions, and by so doing, help the recent Emergency Rice Initiative achieve its important hunger-alleviating mission. This study was up-scaled to integrate and deploy our technology directly into numerous farmers’ paddy rice fields in order to determine whether our candidate inoculants of selected endophytic rhizobial strains can perform as efficient, environmental friendly biofertilizers in existing rice agroecosystems without imposing further stress on their production economy and environmental soundness. These field inoculation trials were conducted over wide production areas representing a range of soil textures, salinity, and other pertinent agroecological characteristics in the Nile delta of Egypt, where the world’s highest paddy rice production yields have been produced during the last ten years (record of 10.04 tons / ha recorded during 2008 (http://beta.irri.org/solutions/index.php?option=com_content&task=view&id=250) and (http://beta.irri.org/statistics/index.php?option=com_content&task=view&id=413&Itemid=192).
Agroecosystems of the Nile delta were ideal for these studies because: 1) their soil fertility (especially N availability) is the major factor limiting rice production, 2) less hazardous and cheaper alternatives to usage of large amounts of chemical fertilizers for high rice production are highly desirable, 3) rice has been rotated successfully with berseem clover (Trifolium alexandrinum) since antiquity and its cumulative ability to increase grain production exceeds the benefit of newly fixed N inputs from mineralization of the previous clover crop residues, 4) the natural rice-rhizobia association has evolved to its present advanced state where it helps to maintain a high inoculum potential of beneficial rhizobia, 5) a large diversity of rice varieties are grown, 6) the rice productivity of some cultivation fields are much lower than the overall national figure and therefore farmers still seek an agroeconomic solution to enhance rice production in an economically and environmentally friendly way, and 7) local rice farmers were willing to share their cultivation fields with us and participate in this scaled-up, mutually beneficial inoculation program. Of special interest to this study was the question of whether biofertilization could increase rice production above Egypt’s national production figure while reducing the need for additional chemical fertilizer inputs. Optimizing biofertilization benefits is especially important for evaluating the production capacity of newly introduced, high N-responsive varieties that currently need 33% more N to maximize grain yield than do the older varieties commonly cultivated in this region before the mid-1980's.
Materials and methods
Rice varieties
Seeds of one Indica and four Japonica rice varieties (Table 1) were obtained from the Rice Research and Training Center, Sakha Agricultural Research Station, Kafr El-Sheikh, Egypt. All are resistant to the rice blast disease caused by the fungus Pyricularia grisea and are more N-responsive (up to 144 kg N/ha) than the older varieties Giza 171 and 172 (96 kg N/ha), which were discontinued because of their overly extended growth period, high water requirements, high susceptibility to blast and lower grain yields.
Bacterial test strains
Seven rice endophyte strains of R. leguminosarum bv. trifolii were used (Table 2), representing the strain diversity of pure culture collections obtained from 2 cycles of isolation from within surface-sterilized field-grown rice roots in the Nile delta (Yanni et al. 1997; Yanni et al. 2001). Each isolate was from a different genotypic group discriminated by its genomic fingerprints and was Nod+ Fix+ on berseem clover in gnotobiotic culture conditions. Their genotypic relatedness based on BOX-PCR and plasmid profiling analyses and to the 16S rRNA sequence of the type strain, fulfillment of Koch’s postulates proving their endophytic colonization ability, and plant growth-promoting activities on several rice varieties in gnotobiotic tube culture were all described previously (Yanni et al. 1997; Yanni et al. 2001).
Preparation of rhizobial inoculants
Inoculants were prepared with peat carrier that had been neutralized from its original pH of 5.0–5.5 using powdered CaCO3. Pure cultures of the tested strains were grown for 72–96 h to a density of 108–109 CFU/ml in yeast extract mannitol (YEM) broth with shaking at 150 rpm and 30°C. Similar culture volumes of either individual strain or balanced mixtures (1:1 volume/volume) of strains were mixed with the peat/CaCO3 carrier to 60% of its water-holding capacity, then cured at room temperature for 24 h and stored in air-tight bags. Samples were plated weekly on YEM agar to enumerate the viable population densities, which were 108–109 CFU/g just before field application.
Field inoculation trials
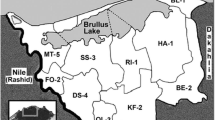
The first field inoculation experiment was conducted at the experimental farm of the Sakha Agricultural Research Station (Kafr El-Sheikh) during the year 2000 rice-growing season (from April/May to September/October). The subsequent 23 experiments were conducted during four consecutive annual rice cultivation seasons (2002–2005) at various paddy rice farms covering 16 ha in 9 counties of the Kafr El-Sheikh Governorate, located in the north-west-central regions of the Nile delta (Fig. 1) hosting approximately 18–25% of the total area used annually for rice cultivation in Egypt. Table 2 lists the identification number assigned to each field inoculation experiment, the rice growing season, the nearest county central city, the name of the cooperating farmer, the previous crop grown in the same field, the rice variety and inoculant strain(s) tested, and the four-times replicated area of the sub-plot. These 24 field experiments plus three previous small-scale field experiments performed at the Sakha Agricultural Research Station, (Yanni et al. 1997;Yanni et al. 2001) represent a total of 27 field inoculation trials for 9 varieties of rice using 7 genotypically distinct, indigenous rhizobial endophyte strains. Soils sampled from the surface to a depth of 30 cm at each experimentation field just before water submergence were classified according to the standard textural triangle (http://www.google.com.eg/search?hl=en&q=soil+triangle+diagram&meta=&aq=0&oq=Soil+triangle) as clay soils at the locations 1, 3, 5, 7, 8, 10, 14, 17, 22–24; as clay-loam at locations 16, 18 and 19; as silty-clay at locations 13, 15, 20 and 21; as silty-clay loam at locations 4, 6, 9 and 12; and as silty-loam at locations 2 and 11. Standard analysis methods (Black et al. 1965; Jackson, 1967) indicated that the soil samples had a pH (v:v 1:2.5) ranging between 7.8 and 8.1, CaCO3 content between 1.4 and 5.4%, and organic matter content between 1.9 to 2.3%. The salinity of top soil (0–15 cm) was <2,500 ppm at locations 3–5, 7–15, 17–24, between 2,501–5,000 ppm at locations 1 and 6, 7,213 ppm at location 16, and 8,609 ppm at location 2.

Kafr El-Sheikh Governorate (white fill) in the Nile delta where the 24 field inoculation trials were performed. Shown are the abbreviated name and number of field experiments conducted in each county, a satellite image of the area location within the entire Nile Delta (insert), and the borders surrounding the Kafr El-Sheikh Governorate. County labels are BL Baltem, BE Beila, DS Desouk, FO Fowa, KF Kafr El-Sheikh, MT Metobas, QL Qalein, RI El-Read, SS Sidi-Salem. The borders of the working area are approximately at 31° 34’ 02” N, 31° 01’ 45” S, 31° 14’ 52” E and 30° 31” 08’ W
Rice nurseries (1/12th of each test field area) were established adjacent to the experimental field area approximately 30–35 days before manually transplanting the young seedlings. The soil was plowed three times in perpendicular directions, remains of the previous crop were removed and the soil was leveled using a laser leveler machine. Phosphate at 36 kg phosphorus pentoxide (P2O5) / ha was broadcasted as calcium super-phosphate (15% P2O5) on the dry soil surface. Nile tributary water was then introduced to a height of five to ten cm followed by further soil leveling under water. Rice seeds were enclosed within cloth bags and submerged in water for two days, then removed and left to germinate for one to two days depending on the variety. The emerged seedlings were uniformly broadcasted to a density of 115 g (dry seed basis) / m2 during a calm period (without wind) on the levee-enclosed nursery area covered with water to a depth of 3 cm. The herbicide Benzthiocarb “Saturn 50%” [S-(4-chlorobenzyl)-N, N diethylthiocarbamate] was applied at the rate of 4.8 L/ha four days later. Irrigation was then stopped for 3-4 days, followed by 3 to 4 cycles of irrigation and surface drainage until the seedlings were well established, followed by concomitant increase in the head of standing water proportionally with plant growth.
The experimental test fields were prepared using protocols similar to those followed for the nursery. Seedlings of 28–30 days age were gently uprooted from the nursery and manually transplanted to the experimental field sub-plots in groups of 2 or 3 seedlings at the corners of 20 x 20 cm squares or 15 x 20 cm rectangles (250,000 or 330,000 seedling groups/ha, respectively, according to the rice variety). Rhizobial inoculants contained individual strains or consortia containing two or more strains in 16 field trials of farmer's sub-plots (20 m2 each) and in 8 field trials of larger sub-plots (52.5 m2 each), depending on the availability of land area. The experimental design was the split-plot with N-fertilization rates as the main-plot treatments and inoculation or non-inoculation as the sub-plot treatments. Four replicates were used for each sub-plot treatment. Each of the main and sub-plot treatments and the replicates were randomly distributed to assist sound field experimental layout and exclude possible sources of experimental errors that may originate from uncontrolled factors normally accompanying field experimentation. The rest of the field area was used by the land owner farmer to cultivate the same rice variety by using his own traditional field management practice or by following the recommendations provided by the Agricultural Research Center, Egypt (http://www.arc.sci.eg/) for rice farming practices. Typically, the farmer’s practice would include mistakes such as poor field leveling, irregular transplantation with some unused land areas, no inoculation, application of excessive quantities of N-fertilizers, improper timing of their application, uneven distribution of the fertilizer in the sub-plot area, insufficient pest management practices, insufficient or excessive irrigation water supplied and/or inefficient drainage system. Inoculation was by manual broadcast of 720 g/ha peat-based inoculum (109 colony-forming units / g) on the flooded water surface one or two days after transplantation of the rice seedlings. This was done from outside the field plot during a windless calm period at sunset to avoid dispersal of the inoculum to uninoculated plots and to minimize exposure of the inoculant populations to adverse effects of direct sunlight. Potassium sulphate (48% K2O) was applied to all test sub-plots at the rate of 240 kg/ha at one month after transplantation. Manual weeding and different pesticides for fungal and insect pests were applied as common field treatment when necessary. Water supply was continued regularly to maintain a standing head starting at 3 cm, then subsequently increased to 12–15 cm as the plant height increased, and finally stopped 15 days before harvest. To compare the growth response to inoculation vs. application of N-fertilizer, replicate main plots were fertilized with 48, 96 or 144 kg N [applied as urea (46% N)] / ha in two equal doses, the first at 15 days and the second at one month after transplantation. The total fertilizer-N amounts for the two doses represent 1/3, 2/3 and the full amount of fertilizer-N recommended by the Agricultural Research Center, Egypt for maximum grain yield of the rice varieties tested. Grain and straw yields were measured at harvest using the entire sub-plot area (no sub-sampling). The harvest index [% of grain yield / (grain plus straw yields)] and the agronomic fertilizer N-use efficiency (kg grain yield / kg fertilizer-N applied) were calculated. Samples for yield comparison were taken from uninoculated crop varieties cultivated during the same period in the adjacent field areas managed by the farmers without direct supervision from the research and technology teams. Ten field areas (12 m2 each) randomly distributed to represent the whole field area managed independently by the farmer were harvested. Mean of grain yield of these samples was considered the farmer’s yield in the adjacent field area. Data obtained from each experiment were statistically analyzed as a split-plot design experiment with three N-fertilization doses as the main-plot treatments, the inoculated and no-inoculated control as sub-plot treatments, with four replicates. The mean differences were compared to their corresponding least significant differences at the 95% confidence level.
Results
Grain yield
Biofertilization significantly increased paddy rice grain production in nineteen (79.2%) of the 24 field experiments, as compared to the corresponding uninoculated controls receiving the same experimental research practices (Table 3). The overall statistically significant increases in grain yield resulting from inoculation for all 24 field trials ranged from 0.268 to 1.499 ton/ha (mean±std. dev. of 0.737 ± 0.281 ton/ha). The average increases in grain production in three other trials where differences were not statistically significant ranged from 0.108 to 0.582 ton/ha. In two experiments (locations 5 and 12) overall averages for the non-inoculated treatments were higher than the inoculated ones but the differences were not statistically significant (all three rates of fertilizer-N application combined).The benefit of inoculation represented by statistically significant increases in grain yield as compared to the corresponding uninoculated counterpart at each N-dose applied is highlighted in Table 3 and in Fig. 1a (in Online Resource 1). Figure 1b-d (in Online Resource 1) indicate the benefit of inoculation expressed as the means of overall increases in paddy grain yield under each N-fertilization dose tested.
Grain yield increased concomitantly with increased application of fertilizer-N in all 24 field trials. Statistically significant increases in grain yield resulted from inoculation plus application of 48, 96 and 144 kg N/ha in 16, 19 and 18 (66.7, 79.2 and 75%) of the 24 experiments, respectively. Interestingly, inoculation significantly increased grain yield with application of the intermediate doses of 48 or 96 kg fertilizer-N/ha in 9 of the 24 field trials where the increases were close or equal to the yield obtained with the full recommended dose of fertilizer-N without inoculation (Tables 3 and 4, Fig. 1b-d in Online Resource 1). This result indicated that rhizobial inoculation could reduce the inputs of chemical N-fertilizer needed to achieve maximum rice grain production. Especially interesting were the results of field trials in experiments # 2, 6 and 9 where inoculation significantly increased grain yield even when no statistically significant increases in yield were obtained by application of the maximum recommended dose of fertilizer-N without inoculation (Table 3).
Certain consortia of rhizobial strains performed better than single rhizobial strains as rice biofertilizer inoculants (Table 3). The best performing biofertilizers were consortia containing the endophyte strains E11 + E12, which increased grain yield by an overall mean that was 8.2% higher than uninoculated, as compared to an overall mean increase of 5.4% for single strains or consortia that did not contain strains E11 and E12.
Biofertilization increased productivity in some of the experiments to levels that exceeded the Egyptian average grain production figure (Table 3). Production capacity increased by inoculation in 7 of the 24 experiments (trials #8, 10, 15, 19, 22–24) when it was not achieved by chemical N-fertilizer application alone. In those experiments, the mean of grain yields with application of 48 kg N/ha was 8.185 tons/ha without inoculation, and increased to 8.597 tons/ha with inoculation. The means of grain yield with application of 96 and 144 kg N/ha were 8.846 and 9.569 without inoculation and increased to 9.605 and 10.231with inoculation, respectively. Increases in response to inoculation for the other 17 experiments are indicated in Table 3. Grain yield exceeded the national production figure in 33% of the inoculation experiments with 48 kg fertilizer-N/ha, 62.5% with 96 kg fertilizer-N/ha, and in 87.5% with the full recommended dose of 144 kg N/ha. In each of these cases, the increase in grain yield resulting from inoculation was significant at the 95% confidence level. The overall mean of grain yield for all 24 field tests was 9.155 ton/ha without inoculation, and increased to 9.700 ton/ha with inoculation. The corresponding overall mean of grain yield with application of 96 kg N/ha was 9.836 without inoculation and increased to 10.582 with inoculation, and with application of 144 kg N/ha was 10.467 without inoculation, increased to 10.986 ton/ha with inoculation..
Table 4 compares the grain yields obtained by the farmers using their conventional practices versus the researcher yields on adjacent experimental fields obtained with and without biofertilization. Mean grain yield was 13.6% higher using researcher’s treatments without inoculation and increased up to 47% with an average of 19.5% with inoculation. This result emphasizes the value-added benefit of incorporating proper agricultural extension activities when using rhizobial biofertilizers to enhance rice grain production.
Straw yield
Table 5 reports the response of straw production to inoculation and fertilizer-N applications for the 24 field trials. Inoculation increased the overall straw production in 19 of the 24 field experiments, with 14 trials (58.3% of the total) being statistically significant where the increases ranged between 0.417 to 1.037 ton/ha, mean of 0.740 ton/ha. The overall means (± std. dev.) of straw production for all the 24 experiments without inoculation were 12.348 ± 2.728 ton/ha with application of 48 kg N/ha, increased to 13.560 ± 2.764 with 96 kg N/ha, and increased further to 14.617 ± 2.754 with application of 144 kg N/ha. The corresponding figures for straw yield following inoculation were 12.743 ± 2.605, 14.033 ± 2.563 and 14.758 ± 2.419 tons/ha, respectively. Significant increases in straw yield due to inoculation with application of 48, 96 and 144 kg N/ha were obtained in 8, 14, and 7 of the 24 experiments, respectively. Detrimental lodging (bending over of top-heavy stalks due to a disproportional increase of vegetative growth response to excessive application of mineral N fertilizers) did not occur in any of the 24 experiments because the tested rice varieties received no more than the recommended amount of N-fertilizer and were of the short stature type that does not typically exhibit this adverse growth response.
Harvest index
The overall means of harvest indices recorded for all three levels of N-fertilizer applications were higher with inoculation in 19 of the 24 field trials compared to corresponding uninoculated controls, but were statistically significant in only 5 experiments (Table 1 in Online Resource 1). When considering the three different levels of N-fertilization, significant increases due to inoculation were recorded in 3, 3 and 5 experiments with application of 48, 96 and 144 kg N/ha, respectively. In 6 experiments, increasing the N-fertilization rate significantly decreased the harvest index, indicating that unlike for inoculation, chemical N-fertilization increased straw production more so than it increased grain production, consistent with results reported above on straw yield. In the remaining 18 experiments, N fertilizer applications of up to 144 kg N/ha did not significantly lower the harvest index, indicating that the recommended N-dose used in those experiments was within the acceptable range that balanced vegetative and reproductive growth.
Agronomic N-use efficiency
This metric helps to evaluate the mineral fertilization strategy for field crops and reflects their agronomical, economical and environmental efficiencies in using that resource input for grain production. As anticipated, overall means of the agronomic N-use efficiency for the inoculated and non-inoculated sub-treatments decreased sharply in all the 24 field experiments with increased application of fertilizer-N. Twenty-three of these overall mean differences represent statistically significant increases due to inoculation (Table 2 in Online Resource 1). Inoculation enhanced N-use efficiency in 18, 17 and 13 experiments with application of the ascending fertilizer doses of 48, 96 and 144 kg N/ha, respectively.
Discussion
Our previous results of three, relatively small field inoculation trials (Yanni et al. 1997; Yanni et al. 2001) combined with the extensive up-scaled field inoculation program of 24 experiments reported here indicate that inoculation of rice with biofertilizers containing certain rhizobia can effectively and significantly enhance rice production while reducing the need for additional fertilizer-N inputs. For this program to be highly representative of the rice agro-ecosystem in the northern Nile delta and helpful for testing the biofertilization technology used, it was conducted in cooperation with 20 participating farmers whose expertise in rice farming varied considerably, using 7 genotypically distinct, endophytic rhizobial test strains, 5 blast-resistant high N-responsive Indica and Japonica rice varieties, 5 rice growing seasons, and covered a total of 16 hectares of farmers fields with variable soil characteristics distributed among 9 county geographic locations (Tables 1 and 2, Fig. 1). Results obtained in this transitional research program documented the benefits of biofertilization to produce higher rice grain yields, thereby offsetting land and water usage requirements plus need for additional fertilizer and labor costs, providing significant help to sustainable rice production over the long term. This study represents an example of how deployment of well-selected biofertilizers should be an integral component of sustainable nutrient management for rice production in an environmentally safe way when soil fertility (especially available N) is a limiting factor. Thus, in addition to its well-known use as a micro-partner in the N2-fixing root-nodule symbiosis with legumes, rhizobia deserve serious consideration as a microbial biofertilizer that can significantly enhance rice production.
Unlike the well-known suppression of symbiotic nitrogen fixation resulting from application of N-fertilizers, the degree of benefit to rice by inoculation with rhizobia was enhanced rather than suppressed with application of N-fertilizer, consistent with a mechanism that involved increased sequestration and utilization of N from available soil N pools rather than from de novo biological nitrogen fixation. Previous evidence for that mechanism was supported by studies indicating that inoculation of rice with endophytic rhizobia enhanced seedling growth vigor, modulated the levels of phytohormone growth regulators, and developed an expansive root architecture with increased biomass, cumulative length and biosurface area and enhanced ability to accumulate various macro- and micro-nutrients including N, P, K, Ca, Mg, Zn, and Mo (Biswas et al. 2000b; Chi et al. 2005; Yanni et al. 1997; Yanni et al. 2001). Cases where inoculation increased grain production despite an inability of the maximum recommended dose of fertilizer-N to do so indicate that the benefits of rhizobial inoculation on rice grain production extended beyond its alleviation of N-limitation. Thus, this up-scaled experimentation program did not include treatments without added chemical fertilizer-N nor N-fertilization doses above the recommended ones, primarily because prior work already showed that rice biofertilization performs much better when combined with intermediate doses of fertilizer-N (Yanni and Abd El-Fattah 1999; Yanni et al. 1997; Yanni et al. 2001), and also because the farmers would not have cooperated nor provided the land for this work knowing well that the outcome in both cases would be catastrophic to their production economy.
Saline soils are very common in northern coastal regions of the Nile delta. There, submerged rice is predominantly grown as an economically staple cereal crop and also because soil reclamation can be facilitated by water percolation through the soil profile that washes the high salt content downwards to groundwater and then to drainage canals. In this work, the high soil salinity in location #2 near the northern city of Baltem most likely lowered grain yield of the rice variety Giza 178 obtained in the non-inoculated treatment as compared to productivity of the same variety in field experiments #3 – #5 at other locations with less soil salinity (Table 3, Fig.1). Interestingly, the mean increase in paddy rice grain yields resulting from inoculation was highest at the high-saline location for experiment #2, suggesting that inoculation of rice with certain endophytic rhizobial strains may also help to alleviate the adverse effects of salinity stress on its grain production. A similar benefit has been previously found for the root nodule symbiosis between Bradyrhizobium spp. (Cajanus) with pigeon pea and Rhizobium loti with chickpea (Rao and Sharma 1995; Subbarao et al. 1990).
The statistically significant increases in straw yield resulting from inoculation are consistent with earlier studies showing that inoculation with certain endophytic strains of rhizobia can significantly increase rice shoot height, leaf area and photosynthetic capacity (Biswas et al. 2000a; Biswas et al. 2000b; Chi et al. 2005; Yanni et al. 1997; Yanni et al. 2001). Straw biomass production was more responsive to chemical N-fertilization than to rhizobial biofertilization, and in contrast, biofertilization boosted rice grain production more so than it increased straw biomass production. This differential result is reflected in the harvest index (Table 1 in Online Resource 1) and can be considered as the preferred, positive benefit of inoculation (more grain production for human consumption rather than excessive vegetative biomass for animal consumption and wasteful burning). It inevitably leads to the hypothesis that the rhizobial interaction with rice intensifies the plant’s reproductive physiology in ways that increase grain production.
The plant responses to inoculation and N-fertilization worked together synergistically in supplying its demand for N and other nutrient requirements without decreasing the harvest index values in most cases. This result is consistent with our previous work (Yanni et al. 1997; Yanni et al. 2001) showing that rhizobial inoculation and N-fertilization contributed to rice vegetative growth and grain yield in parallel, with the single exception where an excessive N-fertilization dose (above the recommended) resulted in detrimental lodging that decreased grain yield (Yanni et al. 1997). The values of agronomic N-use efficiency obtained in most experiments in this program indicated that rhizobial inoculation can benefit grain production by counter-balancing the tradeoff in reduced agronomic N-use efficiency with increasing fertilizer-N doses, providing the desired result that rhizobial inoculation can reduce the need for additional fertilizer-N application to achieve higher grain yield.
Comparison of rhizobia with other biofertilizer candidates for rice
Previous field inoculation studies on rice have used N2-fixing cyanobacteria, Azospirillum and Azotobacter as biofertilizer inoculants (Arora 1969; Gupta et al. 1989; Jack and Roger 1977; Omar et al. 1993; Rajarmamohan et al. 1978; Roger and Kulasooriya 1980; Shahaby et al. 1993; Subrahmanyan et al. 1965; Venkataraman 1966; Yanni and Abd El-Fattah 1999). However, the performance of rhizobial inoculants differs from those other plant growth-promotive rhizobacteria in two major ways. First, the appropriate rhizobia can benefit both the legume and the cereal host as in the rice-berseem clover crop rotation under which this research program was performed, whereas the other inoculants are only used for the cereal crop. In Egypt, 67% of rice cultivation is done in rotation with berseem clover, and so that rotation can help to perpetuate the populations of clover rhizobia involved in both beneficial plant-microbe interactions. Second, rhizobial inoculants benefit rice at each stage of development, beginning with seed germination and extending through grain maturity (Biswas et al. 2000a; Biswas et al. 2000b; Dazzo and Yanni 2006; Dazzo et al. 1999; Yanni et al. 1997; Yanni et al. 2001), whereas other biofertilizer inoculants benefit rice only during a narrow time interval at its maximum “N-limitation stress period” when an external source of N is required (Yanni and Abd El-Fattah 1999 and references herewith) at the tillering stage (15 to 40 days post transplanting).
Bio-safety of inoculants
It is important to evaluate the biosafety characteristics of biofertilizer inoculants for all of the plant species grown in the same crop rotation. In earlier gnotobiotic culture studies (Yanni et al. 1997; Yanni et al. 2001), we found that some natural rice endophyte strains of clover rhizobia were active root nodulators and efficient N-fixers on berseem clover but inhibitory to rice growth, whereas other natural rice endophyte strains of rhizobia were efficient PGPR+ on rice but lethal pathogens on berseem clover. Since all these various types of endophytic rhizobia-plant interactions exist in nature, it is important to screen for, identify and exclude possible detrimental plant/strain combinations beforehand to avoid catastrophic inoculation outcomes under field conditions. All of the 7 strains of rhizobia used in this study were pre-tested under gnotobiotic conditions and found to be Nod+ Fix+ on berseem clover while PGPR+ on rice (Yanni et al. 2001).
Final comments
The results of this translational research indicate that environmentally friendly rhizobial biofertilizer inoculants can now be recommended with sufficient supporting data to improve agriculture economy and sustainable agro-ecosystem maintenance of increased rice grain production where the benefits of such biotechnology are most urgently needed, e.g., the important hunger-alleviating mission of the Emergency Rice Initiative. However, because rice production often remains “N-responsive” even when combined with rhizobial inoculation, further efforts must be exerted to reasonably eliminate the need for additional fertilizer-N inputs to obtain higher rice yields. Perhaps the solution to achieve that ultimate goal is to critically formulate biofertilizers containing mixed inoculant consortia that include our best-performing rhizobial strains plus highly selected free-living N-fixers like cyanobacteria, “associative” diazotrophs like azospirilla, and other plant-growth promoting rhizobacteria like Pseudomonas. We are currently pursuing this hypothesis.
Abbreviations
- bv:
-
biovar
- CFU:
-
colony forming units
- FAO:
-
Food and Agriculture Organization
- IRRI:
-
International Rice Research Institute
- PGPR:
-
plant growth-promoting rhizobacteria
- YEM:
-
yeast extract mannitol
References
Arora S (1969) The role of algae in the availability of phosphorus in paddy fields. Riso 18:135–138
Black A, Evans DD, Ensmingers FE, White JI, Clark FE, Dinaver RC (1965) Methods of soil analysis. II- Chemical and microbiological properties. No.9, Series of Agronomy, American Society of Agronomy Inc., Madison, Wisconsin, USA
Biswas JC, Ladha JK, Dazzo FB, Yanni YG, Rolfe BG (2000a) Rhizobial inoculation influences seedling vigor and yield of rice. Agron J 92:880–886
Biswas JC, Ladha JK, Dazzo FB (2000b) Rhizobia inoculation improves nutrient uptake and growth of lowland rice. Soil Sci Soc Amer J 64:1644–1650
Chaintreuil C, Giraud E, Prin Y, Lorquin J, Ba A, Gillis M, de Laudie P, Dreyfus B (2000) Photosynthetic bradyrhizobia are natural endophytes of the African wild rice Oryza breviligulata. Appl Environ Microbiol 66:5437–5447
Chi F, Shen SH, Cheng HP, Jing YX, Yanni YG, Dazzo FB (2005) Ascending migration of endophytic rhizobia, from roots to leaves, inside rice plants and assessment of benefits to rice growth physiology. Appl Environ Microbiol 71:7271–7278
Dazzo FB, Yanni YG (2006) The natural Rhizobium-cereal crop association as an example of plant-bacteria interaction. In: Uphoff N et al (eds) Biological approaches to sustainable soil systems. Taylor and Francis, Boca Raton, pp 109–127
Dazzo FB, Yanni YG, Rizk R, deBruijn F, Squartini A, Mateos P, Martinez-Molina E, Biswas J, Ladha JK, Weinman J, Rolfe B, Vega-Hernandez M, Hollingsworth RI (1999) The natural beneficial association between Rhizobium leguminosarum bv. trifolii and rice and its exploitation in sustainable agriculture. Proceedings of the "International Symposium on Nitrogen-Fixation and Crop Production", Cairo, Egypt, May 11–13, 99
Dazzo FB, Yanni YG, Rizk R, de Bruijn FJ, Rademaker J, Squartini A, Corich V, Mateos P, Martínez-Molina E, Velázquez E, Biswas JC, Hernandez RJ, Ladha JK, Hill J, Weinman J, Rolfe BG, Vega-Hernández M, Bradford JJ, Hollingsworth RI, Ostrom P, Marshall E, Jain T, Orgambide G, Philip-Hollingsworth S, Triplett E, Malik KA, Maya-Flores J, Hartmann A, Umali-Garcia M, Izaguirre-Mayoral ML (2000) Progress in multinational collaborative studies on the beneficial association between Rhizobium leguminosarum bv. trifolii and rice. In: Ladha JK, Reddy PM (eds) The quest for nitrogen fixation in rice. IRRI, Los Banos, pp 167–189
Gupta A, Sharma BM, Kerni S (1989) Comparative efficiency of Azospirillum, Azotobacter and blue-green algae in submerged rice culture. Res Develop Reporter 6:169–171
Gutierrez-Zamora M, Martinez-Romero E (2001) Natural endophytic association between Rhizobium etli and maize (Zea mays). J Biotechnol 91:117–126
Hilali A, Prevost D, Broughton W, Hartmann A (2001) Effets de l’inoculation avec des souches de Rhizobium leguminosarum biovar trifolii sur la croissance der le dans deux sols der Maroc. Can J Microbiol 41:590–593
Jack V, Roger PA (1977) Diminution des fonts de semis dues ‘a la sulfatore’ duction, par un pre’traitement des grains de riz avec des cyanophyc’ees. Cabiers ORSTOM Serie Biologie 12:101–107
Jackson ML (1967) Soil chemical analysis. Prentice-Hall, New Delhi
Jha B, Thakur MC, Gentia I, Albrecht V, Stoffels M, Schmid M, Hartmann A (2009) Isolation, partial identification and application of diazotrophic rhizobacteria from traditional Indian rice cultivars. Eur J Soil Biol 45:62–72
Lupway N, Clayton G, Hanson K, Rice W, Bierderbeck V (2004) Endophytic rhizobia in barley, wheat, and canola roots. Can J Pl Sci 84:37–45
Mano H, Morisaki H (2008) Endophytic bacteria in the rice plant. Microbes Environ 23:109–117
Matiru VN, Dakora FD (2004) Potential use of rhizobial bacteria as promoters of plant growth for increased yield in landraces of African cereal crops. African J Biotechnol 3:1–7
Mishra RPN, Singh RK, Jaiswal HK, Singh MK, Yanni YG, Dazzo FB (2008) Rice-rhizobia association: evolution of an alternate niche of beneficial plant-bacteria association. In: Ahmad I, Pichtel J, Hayat S (eds) Plant-bacteria interactions: strategies and techniques to promote plant growth. Wiley-VCH Verlag GmbH, Weinheim, pp 165–194
Omar MNA, Hussein KRF, Zaher EA, Abou-Zeid MY (1993) Effect of inoculation with Azospirillum brasilense on yield and some chemical constituents of rice grain. In: Hegazi NA, Fayez M, Monib M (eds) Nitrogen fixation with non-legumes. American University in Cairo Press, Cairo, pp 493–494
Peng G, Yuan Q, Zhang W, Tan Z (2008) Rhizobium oryzae sp. nov., isolated from the wild rice Oryza alta. Int J System Evol Microbiol 58:2158–2163
Perrine FM, Prayitno J, Weinman J, Dazzo FB, Rolfe BG (2001) Rhizobium plasmids are involved in the inhibition or stimulation of rice growth and development. Austr J Plant Physiol 28:923–937
Prayitno J, Stefaniak J, McIver J, Weinman JJ, Dazzo FB, Ladha JK, Barraquio W, Yanni YG, Rolfe BG (1999) Interactions of rice seedlings with bacteria isolated from rice roots. Austr J Plant Physiol 26:521–535
Rajarmamohan RV, Charyulu PBBN, Nayas DN, Ramakrishna C (1978) Nitrogen fixation by free-living organisms in tropical rice soil. In: Dobereiner J, Burris RH, Hollaender A, Franco AA, Scott DB (eds) Limitation and potential for biological nitrogen fixation in the tropics. Plenum, New York, p 354
Rao DLN, Sharma PC (1995) Alleviation of salinity stress in chickpea by Rhizobium inoculation or nitrate supply. Biologia Plantarum 37:405–410
Reddy PM, Ladha JK, So R, Hernandez R, Dazzo FB, Angeles OR, Ramos MC, de Bruijn FJ (1997) Rhizobial communication with rice: induction of phenotypic changes, mode of invasion and extent of colonization. Plant Soil 194:81–98
Roger PA, Kulasooriya SA (1980) Blue-green Algae and Rice. International Rice Research Institute, Philippines
Shahaby AF, Amin G, Khalafalla GM (1993) Response of rice and tomato seedlings to inoculation with diazotrophs and their culture filtrates. In: Hegazi NA, Fayez M, Monib M (eds) Nitrogen fixation with non-legumes. American University in Cairo, Cairo, pp 375–376
Singh RK, Mishra RPN, Jaiswal HK, Kumar V, Pandey SP, Rao SB, Annapurna K (2006) Isolation and identification of natural endophytic rhizobia from rice (Oryza sativa L.) through rDNA PCR-RFLP and sequence analysis. Curr Microbiol 52:117–122
Singh MK, Kushwaha C, Singh RK (2009) Studies on endophytic colonization ability of two upland rice endophytes, Rhizobium sp. and Burkholderia sp. using green fluorescent protein reporter. Springer Science and Business Media, LLC 2009, published online 30 May 2009
Subbarao GV, Johansen C, Kumar Rao JVDK, Jana MK (1990) Response of the pigeonpea-Rhizobium symbiosis to salinity stress: variation among Rhizobium strains in symbiotic activity. Biol Fert Soils 9:49–53
Subrahmanyan R, Relwani LL, Manna GB (1965) Fertility build-up of rice field soils by blue-green algae. Proc Indian Acad Sci Series B 62:252–277
Sun L, Qiu F, Zhang X, Dai X, Dong X, Song W (2008) Endophytic bacterial diversity in rice (Oryza sativa L.) roots estimated by 16S rDNA sequence analysis. Microbial Ecol 55:415–424
Tan Z, Hurek T, Gyaneshwar P, Ladha JK, Reinhold-Hurek B (2001) Specific detection of Bradyrhizobium and Rhizobium strains colonizing rice (Oryza sativa) roots by 16S–23S ribosomal DNA intergenic spacer-targeted PCR. Appl Environ Microbiol 67:3655–3664
Venkataraman GS (1966) Algalization. Phykos 5:164–174
Yanni YG, Abd El-Fattah FK (1999) Towards integrated biofertilization management with free living and associative dinitrogen fixers for enhancing rice performance in the Nile Delta. Symbiosis 27:319–331
Yanni YG, Rizk RY, Corich V, Squartini A, Ninke K, Hollingsworth SP, Orgambide G, de Bruijn F, Stoltzfus R, Buckley D, Schmidt T, Mateos PF, Ladha JK, Dazzo FB (1997) Natural endophytic association between Rhizobium leguminosarum bv. trifolii and rice roots and assessment of its potential to promote rice growth. Plant Soil 194:99–114
Yanni YG, Rizk RY, Abd El-Fattah F, Squartini A, Corich V, Giacomini A, de Bruijn F, Rademaker J, Maya-Flores J, Ostrom P, Vega-Hernandez M, Hollingsworth RI, Martinez-Molina E, Mateos P, Velazquez E, Wopereis J, Triplett E, Umali-Garcia M, Anarna JA, Rolfe BG, Ladha JK, Hill J, Mujoo R, Ng PK, Dazzo FB (2001) The beneficial plant growth-promoting association of Rhizobium leguminosarum bv. trifolii with rice roots. Austr J Plant Physiol 62:845–870
Acknowledgments
This work was supported by projects BIO2-001-017 and BIO5-001-015 (Contract/Agreement Award No. 27 and 015, respectively) of the US-Egypt Science and Technology Joint Fund. We thank the numerous rice farmers listed in Table 2 and our field experimentation experts for their cooperation in this study.
Author information
Authors and Affiliations
Corresponding author
Additional information
Responsible Editor: Peter A.H. Bakker.
Electronic supplementary material
Below is the link to the electronic supplementary material.
Rights and permissions
About this article
Cite this article
Yanni, Y.G., Dazzo, F.B. Enhancement of rice production using endophytic strains of Rhizobium leguminosarum bv. trifolii in extensive field inoculation trials within the Egypt Nile delta. Plant Soil 336, 129–142 (2010). https://doi.org/10.1007/s11104-010-0454-7
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11104-010-0454-7