
Abstract
Land-use type affects gross nitrogen transformation and this information is particularly lacking under varied low temperature conditions. In this study, the effects of land-use type (forest vs. grassland) and temperature (10 vs. 15°C) on gross N transformation rates under aerobic conditions were investigated using the 15N isotope pool dilution technique in the laboratory. Soils were collected from forest and grassland sites in China and Canada. The results showed that gross N mineralization and immobilization rates were significantly higher in forest soils than in grassland soils, while the reverse was true for gross nitrification rates. The higher TC and lower SOCw concentrations in the Chinese soils relative to the Canadian soils were related to the greater gross N mineralization rates and lower gross N immobilization rates in Chinese soils. The greater gross N mineralization rates and lower gross N immobilization rates resulted in much higher inorganic N accumulation and that may increase the risk of NO −3 leaching in the Chinese soils. Increasing temperature significantly increased gross nitrification rates in grassland soils and gross N immobilization rates in forest soils, suggesting that grassland soils maybe more vulnerable to N loss through NO −3 leaching or denitrification (when conditions for denitrification exist) and that conversion of grassland to forest soils may exert less negative effects on the environment by promoting the retention of N and decreasing the production of NO −3 and subsequently the risk of NO −3 leaching under increasing temperature by global warming.
Similar content being viewed by others

Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
Soil N transformations are microbially mediated processes, which are influenced by a number of factors, including the composition and diversity of soil microbial communities, substrate quality and quantity (Compton and Boone 2002; Templer et al. 2003; Grenon et al. 2004). All these factors are influenced by land-use type due to the differences in the quantity and quality of above- and belowground litter addition to the soil, which in turn may lead to changes in soil microbial communities and soil N transformations (Verchot et al. 2001; Patra et al. 2006). In addition, environmental conditions such as temperature also influence soil N transformations by changing microbial activity or community composition associated with a change in substrate affinity and/or ability to access substrates (Nedwell 1999).
Generalizations about land-use type and temperature effects on N cycling are largely based on measurements of net rates of either mineralization or nitrification. However, it is increasingly recognized that net N transformation rates represent only the sum of competing processes, and do not provide adequate information about the rates of individual processes; therefore, increasingly the gross rate of N transformation has been investigated (Hart et al. 1994; Cookson et al. 2002; Booth et al. 2005).
Forests and grasslands are two important land-use types in both Canada and China, especially in western Canada, where the landscape is dominated broadly by forest and grassland uses in the parkland region (Booth et al. 2005), across which the physical and chemical properties of the soil may vary widely due to the different organic matter input from both plant residues and root exudates. For instance, forest soils are typically characterized by high organic carbon (C) concentration and lower pH relative to grassland soils (Booth et al. 2005). Such differences in soil properties are likely to affect the microbially mediated processes such as N transformation (Cote et al. 2000; Chen et al. 2004; Grenon et al. 2004). Despite the availability of 15N isotope tracers (Davidson et al. 1991) and numerical 15N tracing models (Rütting and Müller 2007), the effects of land-use type on gross rates of N transformation are still not well understood (Cookson et al. 2007).
Temperature is one of the most important factors affecting microbial activities in the soil (Joergensen et al. 1990). Increasing incubation temperature will increase microbial activity, and lead to increases in gross N transformation rates (Cookson et al. 2002; Hoyle et al. 2006). However, there are reports that the mineralization process was relatively unaffected by temperature changes (in the range of 3–15°C) as compared to the immobilization process (Andersen and Jensen 2001), and in some cases both gross and net nitrification rates declined with increasing temperature from 15 to 20°C (Stottlemyer and Toczydlowski 1999). Furthermore, little research has been done to quantify the effect of temperature on gross N transformation rates at low temperature (<15°C), a prevailing condition during winter and spring in different parts of the world (Andersen and Jensen 2001; Cookson et al. 2002).
Since northern Canada and China are characterized by a cool growing season, gross N transformation rates under low temperatures are important for our understanding of how land-use types affect N cycling in the cold temperate region. Our questions were: 1) is there any difference in gross N transformation rates across forest and grassland soils in China and Canada? 2) if differences exist, what soil properties (e.g. pH, total C concentration, C/N) are dominant factors that contribute to the differences? and 3) does temperature change in the lower range (10 to 15°C) of temperatures normally found in the northern regions of China and Canada affect gross N transformation rates?
Materials and methods
Site description and soil sampling
Forest and adjacent grassland soils were collected from both China and Canada. The sampling site in China was located in Beian (48°33′N and 126°16′E), in Heilongjiang province. In this region the annual mean temperature is 2°C, and the annual mean precipitation is 440 mm. The sampling site in Canada is located near Linaria (54°12′N and 114°8′W), in Alberta. The site has an annual mean temperature of 3°C and annual mean precipitation of 463 mm (Environment Canada 2007). For each land-use type, three sampling plots (15 by 15 m) were established. After removal of the litter, one composite sample of the top 20 cm of the soil was collected from each plot. Remaining roots and leaf pieces were removed by hand and the soils were left to air-dry at room temperature. Before use, the soil was ground to pass a 2-mm sieve, and stored at 4°C. Soils are abbreviated as FC and GC for forest and grassland soils collected from China, respectively, and FA and GA for forest and grassland soils collected from Alberta, Canada, respectively (Table 1).
Tree composition in the Chinese forest site mainly consisted of aspen (Populus bonatii Levl.), silver birch (Betula pendula), and linden (Tilia miqueliana Maxim.), and the grassland site was seeded with orchardgrass (Dactylis glomerata L.) and mat bulrush (Scirpus trigueter L.). The Canadian forest site consisted mostly of native aspen (Populus tremuloides Michx.) and the grassland site was seeded with a mixture of tall fescue (Festuca arundinacea Schreb.), orchardgrass (Dactylis glomerata L.), and red clover (Trifolium pratense). Mineral soils in the Chinese and Canadian study sites are Kastanozem (Haplic) and Albic Luvisol, respectively, based on the FAO system of soil classification.
Analysis of soil physical and chemical properties
Soil samples were shaken with deionized water (1:2.5 mass:volume ratio) for 10 min, the mixture was left to sit for 2 min and the pH was then measured using a digital type DMP-2 mV/pH meter (Thermo Orion). Total N and total C concentrations were determined using a CN analyzer (NA Series 2, CE Instruments, Italy). Soil mineral-N (NH +4 -N and NO −3 -N) were extracted with 2 mol L−1 KCl solution (1:2.5 mass:volume ratio) by shaking for 1 h on a rotary shaker and the filtrates were stored at 4°C before analyzed with the steam distillation method (Bremner 1996) within 1 week. Water-soluble organic C (SOCw) and N (SONw) were determined by extracting 10 g of soil with 30 mL of deionized water for 30 min (Burford and Bremner 1975), followed by analysis on the TOC-V Total Organic Carbon Analyzer (Shimadzu Corp, Kyoto, Japan). Water holding capacity (WHC) was determined as described by Fierer and Schimel (2002). Briefly, soil samples were weighed into a funnel, with the bottom of the funnel sealed with a cotton plug to prevent water from leaving the funnel. The sample was then saturated with deionized water for 2 h, before the cotton plug was removed, to allow excess water to be drained freely for about 12 h, WHC was calculated based on the water content of the soil.
Soil incubation and chemical analysis
A total of 48 250-mL flasks were used for the incubation experiment. A 30 g of air-dried soil was placed in each flask. The soils were rewetted to 40% WHC by adding deionized water with an automatic pipette. The flasks were divided into two groups per soil, and each group was pre-incubated for 7 days at either 10 or 15°C. After pre-incubation, they were further divided into two subgroups of 12 flasks each, one sub-group of samples were amended with 1.0 mL of 15NH4NO3 to apply 60 mg N kg−1 with 15N atom% excess of 5%, and the other sub-group of samples were amended with 1.0 mL of NH 154 NO3 at the same concentrations and 15N atom% excess as above. The 15N labeled substrates were added by pipetting solutions uniformly over the soil surface, and the final soil moisture content was adjusted to 60% WHC. The two sub-groups were hence exposed to the same treatment, but the 15N labeling was either on NH +4 or NO −3 (‘inorganic paired’ treatment; Mary et al. 1998). Subsequently, the flasks were incubated in the dark for 15 days. During the incubation, the flasks were opened for 10 min each day to renew the atmosphere inside each flask. The moisture content of the incubated soil samples was maintained by adding water every 3 or 4 days to compensate for the amount of water lost through evaporation.
After applying the 15N-labelled N solution, soils were destructively sampled in replicates, at 0.5 h and 1, 3, 6, 10, 15 d after 15N application. Soil mineral-N was extracted by shaking soil samples with 2 mol L−1 KCl and extracts were obtained as described in the previous section. The extracted soils were dried and fine-ground for the analysis of the concentration and 15N abundance of organic N using a CN analyzer (NA Series 2; CE Instruments, Italy) linked to stable isotope ratio mass spectrometer (Optima-EA; Micromass, Crewe, UK) at the Lethbridge Research Centre of Agriculture and Agri-Food Canada. For the quantitative analysis of N concentrations of NH +4 and NO −3 in the extracts, steam distillation of the KCl extract was used (Bremner 1996). A portion of the extract was steam-distilled with MgO to determine NH +4 concentrations on a steam distillation system (Vapodest 20, C. Gerhardt, Königswinter, Germany), thereafter the sample in the flask was distilled again after addition of Devarda’s alloy to determine NO −3 concentrations. The liberated NH3 was collected in 0.005 mol L−1 H2SO4 solutions (Keeney and Nelson 1982). To prevent isotopic cross-contamination between samples from affecting the measurement, 25 mL of reagent-grade ethanol was added to a distillation flask and steam-distilled for 3 min between sample distillations (Hauck 1982). Nitrogen concentrations were determined by titration with 0.01 mol L−1 NaOH using an automatic potentiometric titrator (719s Titrino, Metrohm, Herisau, Switzerland). The H2SO4 solution containing NH +4 was then evaporated to dryness at 65°C in an oven after adjustment of the solution to pH 3 using 0.05 mol L−1 H2SO4 and analyzed for 15N abundance (Feast and Dennis 1996) using the stable isotope mass spectrometer described above.
Calculation and statistical analysis
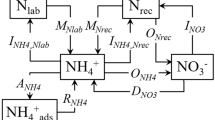
Net mineralization rates were calculated as the difference between inorganic N concentrations in extracts between two sampling dates divided by the number of days in the sampling intervals. Net nitrification rates were calculated in a similar way. Gross N transformation rate was calculated according to the principle of isotopic dilution (Kirkham and Bartholomew 1954) using the FLUAZ model, version 6 (Mary et al. 1998). In the present study, we give the weighted average transformation rates for the interval between d0 and d15 in addition to the individual rates at each sampling interval.
Three-way ANOVAs were used to analyze the effects of land-use type, temperature, country and the interactions of these three factors on gross and net N transformation rates. All data were natural log or arcsine transformed when necessary to meet the assumptions of normality and homoscedasticity; however, non-transformed data are reported in the paper. A one-way ANOVA analysis and Duncan’s multiple range test (DMRT) were used after testing for normal distribution to examine the statistical significance of land-use type effects on gross or net N transformation rates at α = 0.05. Relationships between soil properties and N transformation processes were examined using Spearman’s rank correlation. All statistical calculations were performed using SPSS (SPSS 13.0).
Results
Soil mineral-N concentration
The dynamics of NH +4 and NO −3 in the 10 and 15°C treatments were very similar as shown in Fig. 1. Temperature had little effect on mineral-N concentrations, except for the GC soil, in which the NH +4 concentration decreased and NO −3 concentration increased much faster at 15°C than that at 10°C (P < 0.05), there was little NH +4 left in the GC soil at 15°C by the end of the incubation. In both GA and GC soils, NH +4 concentrations decreased while NO −3 concentrations increased with time, demonstrating the occurrence of net nitrification in the two soils. Whereas in both of the FA and FC soils, NH +4 concentrations increased from day 0 to 15 and NO −3 concentrations remained nearly constant during the entire incubation period.

Changes in ammonium (NH +4 ) and nitrate (NO −3 ) concentrations (mg N kg−1 soil) over time in soils incubated with NH4NO3 at 10°C (closed symbols) and 15°C (open symbols) at 60% WHC. Because NH +4 and NO −3 concentrations were not different after the addition of labeled or unlabeled N, results for the same treatment with labeled or non-labeled N was added were pooled. Vertical bars are standard deviations of the mean (n = 12)
The 15N enrichment in soil mineral-N
The 15N enrichment of ammonium and nitrate pools is shown in Fig. 2. Throughout the incubation at two temperatures a gradual decrease in the 15N enrichment of the NH +4 pool in the 15NH +4 -spiked treatment was observed in all soils (Fig. 2a), indicating low 15N NH +4 input from the mineralization of organic matter that diluted the spiked 15NH +4 . For the GC and GA soils, the increased 15N enrichment in the NO −3 pool over time in the 15NH +4 -spiked treatment was due to nitrification of 15N enriched NH +4 to NO −3 (Fig. 2b). The 15N enrichment of the NO −3 pool in the 15NO −3 -spiked treatment decreased over time as it was slowly diluted by the nitrate produced from nitrification (Fig. 2d).

Changes in 15N abundance of the N components with time when incubated at 10°C (closed symbols) and 15°C (open symbols) at 60% WHC with 15NH +4 (a, b), 15NO −3 (c, d) as a tracer. Vertical bars are standard deviations of the mean (n = 6)
For the FC and FA soils, the isotopic 15N excess of NO −3 pool in both 15N-spiked N treatments remained nearly constant during the entire incubation period (Fig. 2b, d), combined with the data of the dynamics of the nitrate concentration, it suggested that nitrification activity in these two soils was weak. The 15N enrichment of NH +4 in the 15NO −3 -spiked treatment was low and there was no significant change between each sampling time (Fig. 2c).
The temperature effect on the 15N enrichment of mineral N pool was only observed in grassland soils, especially in GC soil (Fig. 2b), where 15N enrichment of the NO −3 pool in the 15NH +4 -spiked treatment was significantly higher at 15°C than at 10°C on most sampling times, indicating that increased temperature promoted nitrification activities in the GC soil.
Nitrogen transformation rates
The gross N mineralization (m), nitrification (n), immobilization (i), NH +4 immobilization (ia), NO −3 immobilization (in) rates in the soils, calculated using the FLUAZ model, are summarized in Table 2. The time-weighted average transformation rates for the interval d0 to d15 and the net N transformations rates are shown in Fig. 3. In further discussion of the land-use type effects described below, the gross N transformation rates mentioned in the text thereafter refers to the weighted average gross N transformation rates between d0 and d15.

Gross N mineralization (A), net N mineralization (B), gross nitrification (C), net nitrification (D), gross N immobilization (E) and ammonium immobilization rates (F) over a 15-day incubation period as affected by temperature and land-use type. Different letters indicate significant differences among soils at P < 0.05 for each incubation temperature. Vertical bars are standard deviations of the mean (n = 3)
There was a main effect of land-use type on all N transformation rates (P < 0.001), and effects for country on nitrification and immobilization rates. The temperature effect (P < 0.05) was only found for gross nitrification and immobilization rates (Table 3). There were no interactions among land-use type, temperature and country for each N transformation rate, except the interaction of land-use type and country on gross and net nitrification rates.
Gross N mineralization rates for all soils generally attained the highest values during the time interval d0 to d1, decreased subsequently and reached the lowest values towards the end of the incubation (Table 2). Our data suggest that a flush of gross N mineralization may have occurred at the beginning of the incubation after the addition of the 15N-labeled fertilizer solution. For all soils studied, temperature had no effect on gross N mineralization rates in all time intervals (Table 2). Gross and net N mineralization rates were significantly higher in the forest soil than in the grassland soil, regardless whether they were Chinese or Canadian soils at both 10 and 15°C incubations (Fig. 3A, B). No significant differences were observed between the Chinese and Canadian grassland soils for the gross or net N mineralization rates, while net N mineralization rates were significantly higher in the Chinese than in the Canadian forest soil when incubated at 15°C. Gross N mineralization rates were about 1.1- to 5.0-fold that of net N mineralization rates at the two temperatures, with the difference much greater at 15 than at 10°C.
The gross nitrification rates in grassland soils (GC and GA) first increased with time and then decreased, with the highest rates occurred between d6 and d10. In contrast, the gross nitrification rates in forest soils (FC and FA) fluctuated and remained nearly constant during the whole incubation period (Table 2). When incubation temperature was raised from 10 to 15°C, nitrification activity significantly increased in GC and GA soils in most time intervals. However, gross nitrification rates were not significantly different between the two forest soils (Table 2, ANOVA data not shown). The land-use type had opposite effects on gross and net nitrification rates in comparison with gross and net N mineralization rates (Fig. 3C, D). As shown in Fig. 3, the gross and net nitrification rates in the GC soil were significantly higher than that in the FC soil at both temperatures, the same phenomenon was also found in Canadian soils but significant only at 15°C. There was no difference between FC and FA soils for the gross and net nitrification rates at both temperatures, but the difference between GC and GA soils was significant, with the highest rate occurred in GC, followed by GA. The magnitude of rates of gross vs. net nitrification was very similar, irrespective of incubation temperature.
Based on results from the FLUAZ model, NH +4 immobilization occurred in all four soils, whereas there was no NO −3 immobilization in GC and FA soils, and a small amount of NO −3 immobilization in FC and GA soils (Table 2). The proportion of the NH +4 immobilized ranged from 65 to 100% of the total mineral N immobilization in all time intervals investigated, indicating that ammonium was the main form consumed by microbes in the soil. In all soils, NH +4 immobilization rates were considerably greater during the first day after addition of the NH4NO3 solutions in relation to the immobilization rates observed during the rest of the incubations (Table 2).
The temperature effects on gross N immobilization and NH +4 immobilization rates were more pronounced in the two forest soils than in the two grassland soils (Table 2). Gross N immobilization and NH +4 immobilization rates were greatly affected by land-use type, being significantly higher in the forest than in the grassland soil at 15°C, regardless of the origin of the soil. Gross N immobilization and NH +4 immobilization rates were significantly higher in the Canadian than in the Chinese soils regardless of land-use type (Fig. 3E, F). In all soils, the average gross N immobilization rates were lower than or nearly equal to the average gross N mineralization rates, resulting in net N mineralization during incubation of these soils.
In this study, the ratio of gross nitrification to NH +4 immobilization rates (N/IA) was used to assess the relative dominance of NH +4 consumptive processes (Fig. 4). For the FC, FA, GA soils, gross nitrification rates were significantly lower than NH +4 immobilization rates, resulting in an N/IA ratio less than 1. While for the GC soil, the gross nitrification rate was much higher than gross NH +4 immobilization rate, with the N/IA ratio being 4.2 and 8.9 at 10 and 15°C, respectively.

The ratio of gross nitrification to gross ammonium immobilization rate (N/IA ratio) over a 15-day incubation period as affected by temperature and land-use type. Different letters indicate significant differences among soils at P < 0.05 for each incubation temperature. Vertical bars are standard deviations of the mean (n = 3)
Discussion
The temperature effect
Soil temperature is a major factor controlling microbial metabolism (Joergensen et al. 1990). Increasing incubation temperature is expected to increase microbial activity and increase gross (Cookson et al. 2002; Hoyle et al. 2006) as well as net N transformation rates (Hoyle et al. 2006). In our study, temperature did not affect gross N mineralization rates, but significantly affected gross N immobilization and gross nitrification rates in most incubation time intervals (Table 2), consistent with Andersen and Jensen (2001), who reported that immobilization process may be very sensitive to temperature change whereas the mineralization process seemed to be relatively unaffected within the temperature range of 3 to 15°C. Nitrification has also been reported to be more sensitive to temperature change than mineralization (Van Schöll et al. 1997), as nitrite oxidizers appear to be more sensitive than NH +4 -N oxidizers to low temperature (Tyler and Broadbent 1960). However, it is interesting to note that gross N immobilization rates in forest soils increased significantly in most incubation time intervals when the temperature was raised from 10 to 15°C, whereas gross N immobilization rates in grassland soils were not affected by the change of temperature in our study (Table 2). This may be due to the influence of N immobilization by soluble organic carbon concentration (Magill and Aber 2000), while the effects of temperature change on the size of the labile C pool in the soil was dependent on the type of organic matter (Hoyle et al. 2006), the decomposition and release of available C in forest soils maybe more sensitive than grassland soils to temperature changes, thus increasing temperature from 10 to 15°C was likely to significantly increase the amounts of soluble organic carbon in forest soils and consequently promoted N immobilization in the soil; however, the underlying mechanism needs to be further studied. Support for our assumption was given by Zaman and Chang (2004) and Cookson et al. (2007) who found that the temperature effects on the amounts of soluble organic carbon depended on the land-use type. The lack of obvious response in gross nitrification rates of forest soils to increasing temperature relative to grassland soils maybe due to the rather low nitrification activity in forest soils (Table 2). A lack of gross nitrification response in forest soils to temperature changes in a similar range (5–20°C) has also been reported by Stottlemyer and Toczydlowski (1999).
The land-use type effect
Nitrogen transformations are microbially mediated processes and it has been well established that the quality of organic matter input, a factor associated with land-use type, can affect the microbial community, soil physical and chemical properties and ultimately processes of soil N transformations (Cote et al. 2000; Chen et al. 2004; Grenon et al. 2004). This study also found that there were significant differences in gross and net N transformation rates between the two types of land-uses.
The initially high and then declined over time of the gross N mineralization rates reflected the decrease in the quantity of the decomposable substrate. Our study showed that gross and net N mineralization rates in the forest soils were more than double those in the grassland soils (Fig. 3A, B), consistent with those of McKinley and Blair (2008), who also found that gross and net N mineralization in forest soils were greater than those in grassland soils. Uri et al. (2008) reported that annual net N mineralization in the surface 10 cm forest soil layer was approximately two times that in grassland soils. We found that gross and net N mineralization correlated well with total carbon concentrations (TC) across different soils (Table 4), indicating that TC was an important factor affecting nitrogen mineralization in soils (Accoe et al. 2004; Booth et al. 2005). Organic substrates are sources of energy for the microbes (Zaman et al. 1998; Schimel and Bennett 2004). A higher amount of TC in the soil can therefore support greater microbial activities and greater mineralization potential (Gaillard et al. 1999). In this study, the greater TC concentrations in forest than in grassland soils might have contributed to the observed differences in gross and net N mineralization rates among four soil types.
Both gross and net nitrification rates were higher in grassland than in forest soils regardless of the country of origin, with the Chinese grassland soil exhibited the strongest nitrification potential among all soils investigated (Fig. 3C, D). This is probably related to the higher and closer to neutral pH in grassland than in forest soils (Table 1), as there were positive correlations between gross and net nitrification rates and soil pH (Table 4). The increases of nitrification rates with pH, similar to those reported by Bååth and Anderson (2003), might have reflected structurally and functionally different microbial communities under different soil pH conditions across forest and grassland soils (Zogg et al. 1997; Cookson et al. 2007). It is generally thought that the occurrence of nitrification in acid forest soils was dominant by acid-tolerant autotrophs despite the significant potential for heterotrophic nitrification by fungi (De Boer and Kowalchuk 2001), whereas autotrophic nitrifier are considered to be highly sensitive to pH, it has been reported that the autotrophic ammonia-oxidizing bacteria generally do not grow below pH 5.0–5.5 and grow faster in neutral or slightly alkaline media (De Boer and Kowalchuk 2001). In our study, the GC soil had pH of 6.43, a near neutral environment, which is about 1 unit higher than forest soils (pH = 5.08 to 5.29), thus the GC soil had the highest nitrification potential which may be associated with the greater activities of nitrifiers.
The land-use effect on the immobilization of inorganic N was similar to gross N mineralization (Fig. 3), with rates in forest soils higher than that in grassland soils regardless the country of origin. However, the country effect on gross N immobilization rates was in contrast with that on gross N mineralization rates, with the gross N immobilization rates significantly higher in the Canadian than in the Chinese soils, and gross N mineralization rates significantly higher in the Chinese than in the Canadian soils, regardless of land-use type. Correlation analysis indicated that gross N immobilization rates were positively correlated with SOCw (P < 0.01) but no obvious correlation was found between gross N immobilization rates and TC (Table 4), indicating that the immobilization of N in the soil was mainly controlled by the availability of energy. Others had also found that microbial consumption of N was influenced by soluble organic carbon concentration and microbial mineralization of N was influenced by total carbon concentration (Hoyle et al. 2006), which further suggests the importance of the availability of C for microbial immobilization of N (Compton and Boone 2002). For example, Gibbs and Barraclough (1998) have previously demonstrated that the addition of a labile fraction of soil organic matter to the soil did not affect gross N mineralization, but markedly increased N immobilization, and they proposed that the bacteria that participate in N immobilization could only utilize simpler (or more labile) forms of C substrates and were incapable of utilizing more complex organic substrates, but the bacteria that participate in N mineralization could utilize a range of organic matter type and was not affected by substrate quality. Therefore, it is likely that the higher concentrations of SOCw in the Canadian soils contributed to the higher gross N immobilization rates relative to the Chinese soils despite the lower TC concentrations in the Canadian soils.
In the four soils studied, the gross NH +4 immobilization rates in the first day after addition of the NH4NO3 solution were considerably higher than those in the rest of the incubation, consistent with the result of Accoe et al. (2005). The large amount of NH +4 -N added to the soil might have stimulated NH +4 immobilization. In our soils, the lack of or the small amount of NO −3 immobilization (Table 2) may be related to the lower metabolic energy cost when NH +4 –N was assimilated than assimilatory reduction of NO −3 –N by microbes, and there was a preferential uptake of NH +4 when both inorganic N forms were present in the soil (Recous et al. 1990).
Many studies had reported that gross rates of mineralization and nitrification generally exceed the net rates by an order of magnitude in forest and grassland ecosystems when measured in situ (Davidson et al. 1991; Hart et al. 1994; Stark and Hart 1997; Verchot et al. 2001; Compton and Boone 2002). Whereas in our study, gross N mineralization rates were only 1.1–5.0 times greater than the net rates, and no obvious difference was observed between gross and net nitrification rates. This may have been caused by sieving and rewetting of air-dried soils that stimulated microbial activity, but gross N immobilization rates increased at a lower rate compared to gross nitrification and gross N mineralization rates (Pulleman and Tietema 1999).
The ratio of gross nitrification to gross ammonium immobilization rate
Microbial nitrification and immobilization of NH +4 are two consumption processes of NH +4 in the soil, the ratio of gross nitrification to gross ammonium immobilization rate (N/IA) could be used as an effective indicator for assessing the relative importance of the two consumption processes (Hoyle et al. 2006). For the GC soil, the importance of nitrification as an NH +4 -N consuming process is evident, as gross nitrification rates were 4–9 times higher than ammonium immobilization rates (Fig. 4). In contrast, the ratio of N/IA in the FC, FA, GA soils was less than 1 (Fig. 4), suggesting that immobilization was the dominant process controlling the NH +4 consumption. Microbial immobilization of NH +4 was more favourable under high SOCw concentration (Seely and Lajtha 1997; Jaeger et al. 1999). Increases in soluble organic carbon have also previously been shown to contribute to increased microbial immobilization of N in forest soils (Magill and Aber 2000; Cookson et al. 2007). In this study, SOCw concentrations in the FC, FA, GA soils were much higher than that in the GC soil (Table 1), leading to the high immobilization of NH +4 in these three soils (Fig. 3F). The N/IA ratio reflected the rates of inorganic N being nitrified or immobilized by soil microorganisms, as a result, it may be used to determine the risk of increasing NO −3 losses within a soil, and the impact of land-use type on N retention versus loss pathways (Tietema and Wessel 1992; Stockdale et al. 2002). In this study, the N/IA ratios for grassland soils were larger than that for forest soils (Fig. 4), indicating the greater potential for NO −3 loss in grassland soils, especially in the GC soil.
Implications of differences in gross N transformation rates in cold temperate region
Few studies have examined the effects of land-use type on gross N transformation rates at low soil temperatures (<15°C) typical of cold temperate regions (Andersen and Jensen 2001; Cookson et al. 2002). Forests and grasslands are two important land-use types in northern Canada and China (Booth et al. 2005), where low temperatures are normally experienced. Our findings that higher gross mineralization and immobilization rates but lower gross nitrification rates in forest soils than grassland soils indicate that the forest soils had a higher potential to release and conserve available nutrients such as NH +4 -N and had a lower risk for N loss by leaching. The temperature change from 10 and 15°C did not affects gross N mineralization rates, but increased gross nitrification rates in grassland and gross immobilization rates in forest soils, indicating that conversion from grassland to forest soils when temperature increases by global warming may exert less negative effects on the environment by promoting the retention of N and decreasing the risk of NO −3 leaching. However, a more comprehensive evaluation of land-use type effects on the environment, particularly on the greenhouse gas emissions needs to be further investigated.
In this study, the TC concentration in the FA soil was lower than that in the FC soil and was comparable between the GA and GC soils, but SOCw concentrations in the FA and GA soils were significantly higher than that in the FC and GC soils, respectively (Table 1). This could be related to the input of more easily decomposed organic matter in the Canadian soils, which is dependent on the input of plant residues and root exudates from different tree and grass species (Booth et al. 2005). The differences in TC and SOCw concentrations between the Chinese and Canadian soils likely lead to the differences in gross mineralization and gross immobilization rates of N. Higher gross N mineralization and lower gross N immobilization rates contributed to the higher inorganic N concentration (Table 1) and greater N availability to nitrifiers in the Chinese as compared to the Canadian soils. The greater inorganic N in the Chinese soil could provide higher N availability to plants but could also pose greater risk of leaching loss during heavy rainfall events.
References
Accoe F, Boeckx P, Busschaert J et al (2004) Gross N transformation rates and net N mineralization rates related to the C and N contents of soil organic matter fractions in grassland soils of different age. Soil Biol Biochem 36:2075–2087
Accoe F, Boeckx P, Videla X et al (2005) Estimation of gross nitrogen transformations and nitrogen retention in grassland soils using Fluaz. Soil Biol Biochem 69:1967–1976
Andersen MK, Jensen LS (2001) Low soil temperature effects on short-term gross N mineralization-immobilization turnover after incorporation of a green manure. Soil Biol Biochem 33:511–521
Bååth E, Anderson TH (2003) Comparison of soil fungal/bacterial ratios in a pH gradient using physiological and PLFA-based techniques. Soil Biol Biochem 35:955–963
Booth MS, Stark JM, Rastetter E (2005) Controls on nitrogen cycling in terrestrial ecosystems: a synthetic analysis of literature data. Ecol Monogr 75:139–157
Bremner JM (1996) Nitrogen-total. In: Sparks DL (ed) Methods of soil analysis. Part 3. Chemical methods. Soil Science Society of America, Madison, pp 1085–1121
Burford JR, Bremner JM (1975) Relationships between the denitrification capacities of soils and total water soluble and readily decomposable soil organic matter. Soil Biol Biochem 7:389–394
Chen CR, Xu ZH, Mathers NJ (2004) Soil carbon pools in adjacent natural and plantation forests of subtropical Australia. Soil Sci Soc Am J 68:282–291
Compton JE, Boone RD (2002) Soil nitrogen transformations and the role of light fraction organic matter in forest soils. Soil Biol Biochem 34:933–943
Cookson WR, Cornforth IS, Rowarth JS (2002) Winter soil temperature (2–15 °C) effects on nitrogen transformations in clover green manure amended or unamended soils: a laboratory and field study. Soil Biol Biochem 34:1401–1415
Cookson WR, Osman M, Marschner P et al (2007) Controls on soil nitrogen cycling and microbial community composition across land use and incubation temperature. Soil Biol Biochem 39:744–756
Cote L, Brown S, Pare D et al (2000) Dynamics of carbon and nitrogen mineralization in relation to stand type, stand age and soil texture in the boreal mixed wood. Soil Biol Biochem 32:1079–1090
Davidson EA, Hart SC, Shanks CA et al (1991) Measuring gross nitrogen mineralization, immobilization, and nitrification by 15N isotopic pool dilution in intact soil cores. J Soil Sci 42:335–349
De Boer W, Kowalchuk GA (2001) Nitrification in acid soils: microorganisms and mechanisms. Soil Biol Biochem 33:853–866
Environment Canada (2007) Canadian climate data online 1993–2007. PhysicsWeb. http://www.climate.weatheroffice.ec.gc.ca/climateData/canada_e.html. Accessed 27 December 2007
Feast NA, Dennis PE (1996) A comparison of methods for nitrogen isotope analysis of groundwater. Chem Geol 129:167–171
Fierer N, Schimel JP (2002) Effects of drying-rewetting frequency on soil carbon and nitrogen transformations. Soil Biol Biochem 34:777–787
Gaillard V, Chenu C, Recous S et al (1999) Carbon, nitrogen and microbial gradients induced by plant residues decomposing in soil. Eur J Soil Sci 50:567–578
Gibbs P, Barraclough D (1998) Gross mineralization of nitrogen during the decomposition of leaf protein I (ribulose 1, 5-diphosphate carboxylase) in the presence or absence of sucrose. Soil Biol Biochem 30:1821–1827
Grenon F, Bradley RL, Titus BD (2004) Temperature sensitivity of mineral N transformation rates, and heterotrophic nitrification: possible factors controlling the post-disturbance mineral N flush in forest floors. Soil Biol Biochem 36:1465–1474
Hart SC, Nason GE, Myrold DD et al (1994) Dynamics of gross nitrogen transformations in an old-growth forest: the carbon connection. Ecology 75:880–891
Hauck RD (1982) Nitrogen-isotope ration analysis. In: Page AL (ed) Methods of soil analysis. Part 2. Chemical and microbiological properties. Soil Science Society of America, Madison, pp 735–779
Hoyle FC, Murphy DV, Fillery IRP (2006) Temperature and stubble management influence microbial CO2-C evolution and gross N transformation rates. Soil Biol Biochem 38:71–80
Jaeger CH, Monson RK, Fisk MC et al (1999) Seasonal partitioning of nitrogen by plants and soil microorganisms in an alpine ecosystem. Ecology 80:1883–1891
Joergensen RG, Brookes PC, Jenkinson DS (1990) Survival of the soil microbial biomass at elevated temperatures. Soil Biol Biochem 22:1129–1136
Keeney DR, Nelson DW (1982) Nitrogen-inorganic forms. In: Page AL (ed) Methods of soil analysis. Part 2. Chemical and microbiological properties. Soil Science Society of America, Madison, pp 643–698
Kirkham D, Bartholomew WV (1954) Equations for following nutrient transformations in soil utilizing tracer data. Soil Sci Soc Am Proc 18:33–34
Magill AH, Aber JD (2000) Variation in soil net mineralization rates with dissolved organic carbon additions. Soil Biol Biochem 32:597–601
Mary B, Recous S, Robin D (1998) A model for calculating nitrogen fluxes in soil using 15N tracing. Soil Biol Biochem 30:1963–1979
McKinley DC, Blair JM (2008) Woody plant encroachment by Juniperus virginiana in a semiarid native grassland promotes rapid carbon and nitrogen accrual. Ecosystems 11:454–468
Nedwell DB (1999) Effect of temperature on microbial growth: lowered affinity for substrates limits growth at low temperature. FEMS Microbiol Ecol 30:101–111
Patra AK, Abbadie L, Clays-Josserand A et al (2006) Effects of management regime and plant species on the enzyme activity and genetic structure of N-fixing denitrifying and nitrifying bacterial communities in grassland soils. Environ Microbiol 8:1005–1016
Pulleman M, Tietema A (1999) Microbial C and N transformations during drying and rewetting of coniferous forest floor material. Soil Biol Biochem 31:275–285
Recous S, Mary B, Faurie G (1990) Microbial assimilation of ammonium and nitrate in soil. Soil Biol Biochem 22:597–602
Rütting T, Müller C (2007) 15N tracing models with a Monte Carlo optimization procedure provide new insights on gross N transformations in soils. Soil Biol Biochem 39:2351–2361
Schimel JP, Bennett J (2004) Nitrogen mineralization: challenges of a changing paradigm. Ecology 85:591–602
Seely B, Lajtha K (1997) Application of a 15N tracer to simulate and track the fate of atmospherically deposited N in the coastal forests of the Waquoit Bay watershed Cape Cod, Massachusetts. Oecologia 112:93–402
Stark JM, Hart SC (1997) High rates of nitrification and nitrate turnover in undisturbed coniferous forests. Nature 385:61–64
Stockdale EA, Hatch DJ, Murphy DV et al (2002) Verifying the nitrification to immobilisation ratio (N/I) as a key determinant of potential nitrate loss in grassland and arable soils. Agronomie 22:831–838
Stottlemyer R, Toczydlowski D (1999) Nitrogen mineralization in a mature boreal forest, Isle Royale, Michigan. J Environ Qual 28:709–720
Templer P, Findlay S, Lovett G (2003) Soil microbial biomass and nitrogen transformations among five tree species of the Catskill Mountains, New York, USA. Soil Biol Biochem 35:607–613
Tietema A, Wessel WW (1992) Gross nitrification transformations in the organic layer of acid forest ecosystems subjected to increased atmospheric nitrogen input. Soil Biol Biochem 24:943–950
Tyler KB, Broadbent FE (1960) Nitrite transformations in Californian soils. Soil Sci Soc Am Proc 24:279–282
Uri V, Lõhmus K, Kund M et al (2008) The effect of land use type on net nitrogen mineralization on abandoned agricultural land: silver birch stand versus grassland. For Ecol Manag 255:226–233
Van Schöll L, Van Dam AM, Leffelaar PA (1997) Mineralization of nitrogen from an incorporated catch crop at low temperatures: experimental and simulation. Plant Soil 188:211–219
Verchot LV, Holmes Z, Mulon L et al (2001) Gross versus net rates of N mineralization and nitrification as indicators of functional differences between forest types. Soil Biol Biochem 33:1889–1901
Zaman M, Chang SX (2004) Substrate type, temperature, and moisture content affect gross and net mineralization and nitrification rates in agroforestry systems. Biol Fertil Soils 39:269–279
Zaman M, Cameron KC, Di HJ et al (1998) Nitrogen mineralisation rates from soil amended with dairy pond waste. Aust J Soil Res 36:217–230
Zogg GP, Zak DR, Ringelberg DB et al (1997) Compositional and functional shifts in microbial communities due to soil warming. Soil Sci Soc Am J 61:475–481
Acknowledgments
Funding for this work was provided by the China Opportunity Fund of University of Alberta and National Natural Science Foundation of China. The authors would like to thank Agriculture and Agri-Food Canada Lethbridge Research Centre for sample analysis and technical assistance.
Author information
Authors and Affiliations
Corresponding author
Additional information
Responsible Editor: Paul Bodelier.
Rights and permissions
About this article
Cite this article
Lang, M., Cai, ZC., Mary, B. et al. Land-use type and temperature affect gross nitrogen transformation rates in Chinese and Canadian soils. Plant Soil 334, 377–389 (2010). https://doi.org/10.1007/s11104-010-0389-z
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11104-010-0389-z