
Abstract
Eucalypt plantations have been trialled in recent years as a control measure for shallow groundwater associated with secondary salinity. Uncertainty still remains as to the potential growth and water use of these plantations; these relate mainly to the problems associated with drought stress and accumulated solutes in the root zone resulting from saline groundwater uptake. This study investigates the water balance and identifies water sources of a 21 year old unirrigated Eucalyptus grandis W. Hill ex Maiden (flooded or rose gum) and E. camaldulensis Dehnh. (river red gum) plantation over shallow saline groundwater in the Shepparton Irrigation Region of northern Victoria, Australia. Water sources used by the plantation were identified using a monthly water balance approach, together with investigations of stable isotopes of water (deuterium and oxygen-18), soil water and chloride. We found these trees to be heavily reliant on rainwater, and derive approximately 15% of their transpiration requirements from saline (10,000 mg l−1) groundwater at the capillary fringe. Rainfall at the site is relatively low (465 mm year−1 on average) and groundwater uptake provides a stable water source that leads to a slight extension of the growing period of these trees. There is little potential for recharge, with subsurface water moving into the groundwater depression created by tree water uptake.
Similar content being viewed by others


Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
Many irrigation areas both in Australia and overseas have shallow saline groundwater and are experiencing reduced production and associated problems of salt and waterlogging (Ghassemi et al. 1995). As a result of their evergreen nature and relatively high annual water use, eucalypts have been advocated for manipulating the catchment water balance to control salinity, primarily by reducing recharge (Jenkin 1981; Morris and Thompson 1983; Greenwood 1986; George et al. 1999), but may also have potential in discharge management (Schofield et al. 1989).
Vegetation, and in particular the use of evergreen woody species, has received special interest as a control measure, not only due to its reported ability to enhance water use from groundwater, but also to provide productivity from marginal agricultural land. Several studies have reported a lowering of groundwater when compared with surrounding levels (Heuperman et al. 1984; Engel 1986; Bell et al. 1990; George 1990; Schofield 1990; Bari and Schofield 1992; George et al. 1999; Bari and Ruprecht 2003). The ability of trees to control groundwater in this way depends on, among other things, continued water use and growth.
Control of groundwater discharge using trees and other perennial species has been the subject of interest for some years (Morris and Thompson 1983; Greenwood 1986; Farrington and Salama 1996). Although substantial progress has answered some basic questions as to which species to plant, where to plant, and how many and at what density to plant, serious constraints exist. These include the uncertainty of longer rotations in terms of sustained growth and water use. Trees growing in these conditions are often placed under severe stress due to a lack or over supply of water, and/or the presence of salts in the root zone (Morris and Thompson 1983).
For trees to lower groundwater levels, a water balance analysis implies that, in the absence of appreciable lateral movement of groundwater, total evapotranspiration from the revegetated landscape must exceed gross recharge. Indeed, actual evapotranspiration rates higher than rainfall have been observed in some areas where trees grow over shallow groundwater (Greenwood et al. 1992; Fraser et al. 1995; Khanzada et al. 1998; Benyon et al. 2006). The reduction in groundwater level is often assumed to imply groundwater uptake by trees (for example Biddiscombe et al. 1985; Bari and Schofield 1991; George 1991; Heuperman 1995; Morris and Collopy 1999).
The proportion of tree water derived from groundwater is difficult to measure. However, techniques using stable isotopes of water (deuterium and oxygen-18) can be used to help identify the source of water transpired by vegetation under a variety of environments (Brunel et al. 1990; Walker and Brunel 1990; Ehleringer and Dawson 1992; Thorburn et al. 1993a, b; Mensforth et al. 1994; Brunel et al. 1995; Thorburn and Ehleringer 1995; Farrington et al. 1996; Jolly and Walker 1996; Mensforth and Walker 1996a; Taylor 1998).
A number of these studies have explicitly examined the use of saline groundwater by vegetation (Thorburn et al. 1993a; Mensforth et al. 1994; Fraser et al. 1995; Taylor 1998). As trees use saline water, salts are excluded at the root surface, and solutes may accumulate in the root zone (Prenzel 1979; Stirzaker 2002). The amount of salts that accumulate in the root zone largely depend on the solute concentration of the water used, how much is used, and any processes that may remove salts from the system (ie. leaching or diffusion).
Elevated salt concentrations have been observed under eucalypt plantations in Australia (George 1990; Greenwood et al. 1992; Fraser et al. 1995; Heuperman 1995; Morris and Collopy 1999) and overseas (Khanzada et al. 1998; Mahmood et al. 2001). Several authors have considered the long term sustainability of vegetation in groundwater discharge areas in the face of salt accumulation (Morris and Thompson 1983; Williamson 1986). It is generally assumed that water uptake and discharge from groundwater by trees decreases with increasing watertable depth and salinity (Greenwood 1986). This results in a feedback in which groundwater uptake is reduced, potentially resulting in an equilibrium where root zone salinities do not continue to increase indefinitely. Furthermore, studies undertaken on plantations in some saline discharge areas have not shown evidence of solute accumulation where lateral flows or recharging water provide mechanisms for leaching of solutes from the root zone (Taylor 1998; Benyon et al. 1999), while others report an apparent limit to solute accumulation resulting from watertable fluctuation and redistribution of solutes (Khanzada et al. 1998; Mahmood et al. 2001).
An understanding of the temporal nature of water uptake and the movement of water and salts within the soil profile is both complex and dynamic, and is a crucial component in any study on the sustainability of tree plantations in shallow saline watertable areas. Little research has been documented on the ability of salt-tolerant trees to survive and to lower watertables in groundwater discharge areas over extended periods. In this study, our objective is to quantify different water sources of a mature (21 year old) unirrigated Eucalyptus plantation over shallow saline groundwater by combining signatures of naturally occurring stable isotopes of possible water sources, soil water availability and soil chloride. We relate our findings to the implications for management of plantations for groundwater control in similar regions.
Methods
Study site
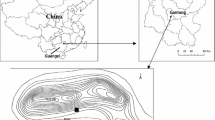
This study was conducted at a 2.4 ha Eucalyptus plantation established in 1976 near the township of Kyabram (Fig. 1a) in an irrigated area in northern Victoria (lat. 36.4333°S, long. 145.2667°E). This study focused on two species within the plantation, namely Eucalyptus grandis W. Hill ex Maiden (flooded or rose gum) and E. camaldulensis Dehnh. (river red gum). The plantation had been irrigated since planting for 6 years until the drought in 1982, and since then, no irrigation water has been applied (Heuperman et al. 1984). Observation bores and deeper piezometers exist throughout the site. The plantation is surrounded by irrigated perennial pasture.

a Location of the plantation at Kyabram (▲) in the state of northern Victoria (shaded), and b Location of monitoring sites 1–3 within the plantation and 4 outside the plantation in irrigated pasture
Soil at the sites is known as Lemnos Loam (Skene and Poutsma 1962) and is derived from unconsolidated alluvial parent material. It is a red brown duplex soil (Dr 2.33; Northcote 1979) and is the soil type most widely used for horticulture and irrigated pasture in the Goulburn Valley region of northern Victoria. It consists of a well-drained loamy top soil and a heavy clay B horizon of low permeability. Drainage is low, leading to surface waterlogging in wet seasons.
Profile studies in the unirrigated plantation reveal that heavy clay extends down to about 5–8 m. A poorly developed and highly variable semi-confined aquifer of loamy fine sand exists from 5 to 8 to 9–11 m below the surface (Heuperman et al. 1984). The average watertable depth at the time of plantation establishment is estimated to have been between 1.65 and 2.04 m (Heuperman 1995). The watertable level under the plantation fluctuated during the first 6 years of irrigation, and then fell to about 4–5 m before steadily rising since monitoring commenced in 1982 to about 3.5 m at present (Heuperman 1995).
The plantation’s relatively low impact on piezometric levels, when compared with watertable levels, is evidence of an upward pressure gradient resulting from shallow groundwater uptake by the trees (Heuperman 1995). Watertable salinity under the trees has risen steadily over time, with electrical conductivities between 5 and 15 dS cm−1 higher than under irrigated pasture; no measurable effect has been observed in deeper groundwater (Heuperman 1995).
The region experiences a Mediterranean climate with cold wet winters and hot dry summers, with an annual rainfall averaging 465 mm. Class A Pan Evaporation is approximately 1,600 mm year−1, and exceeds rainfall for over nine months of the year. The climate of the area is not considered suitable for commercial tree growing without irrigation. Daily weather data was recorded by an automatic weather station about 250 m south of the plantation at the Victorian Department of Primary Industries dairy research centre.
Four monitoring locations within and close to the plantation were identified for measurements of groundwater, isotopic analysis and salinity. These include nests of groundwater observation wells and piezometers within the plantation (sites 1–3) and a fourth location (site 4), a piezometer nest 40 m to the east of the plantation, was used to measure groundwater depth and EC outside the plantation (Fig. 1b). Characteristics of the three sampling sites within the plantation are presented in Table 1.
Water balance
A monthly water balance was constructed using data gathered during the intensive monitoring phase described in Vertessy et al. (2000). This included data on rainfall, canopy rainfall interception, stem flow, tree water use and changes in groundwater levels. Unfortunately, reliable soil water data is not available from that study. The focus of the water balance was to provide a preliminary estimate and range of monthly groundwater use by the plantation. Annual groundwater use (G) was estimated as the difference between effective rainfall (EP) and transpiration (Trans) as:
and
where St is stem flow, estimated at 2% of rainfall (Vertessy et al. 2000), E is evaporation at the soil surface/litter layer (assumed to be 10% of EP after McJannet et al. 1996) and T is throughfall (rainfall minus canopy intercepted rainfall) measured at the site (Vertessy et al. 2000). Due to the lack of relief at the site, and the relatively low rainfall, surface runoff was assumed to be zero.
It is assumed that the soil water storage does not change significantly on a 12 monthly time frame, and that trees preferentially use rainfall when available. The basis for this is that the majority of roots exist in the upper soil zone (Vertessy et al. 2000), and that rain water contains far less solutes than groundwater. Furthermore, the low hydraulic conductivity of the heavy clay soils reduces the rate of downward water movement, allowing time for roots to take up recent rain-derived soil water.
Given the limited length of the dataset (July 1994 to April 1996) and in order to explore the relationships between water source and rainfall, a series of fourteen 12 month (i.e. annual) periods was generated, in which progressive annual totals of the water balance, including the estimates of groundwater use, were compared. For example, the first year was generated with data from March 1994 to February 1995, the second year with data from April 1994 to March 1995 and so on up until the final (and fourteenth year) was generated with data from April 1995 until March 1996. We note that these are not ‘independent’ years.
Groundwater fluxes
Saturated flow into the root zone in response to lateral and vertical hydrostatic gradients was estimated from periodic measurements of watertable levels, with the assumption that there is no groundwater recharge under the plantation. This is reasonable given the conclusions given by previous studies of hydrology at the plantation by Feikema (1995) and Heuperman (1995). Water fluxes into the root zone were calculated to provide an estimate of the lateral and vertical supply of water to the roots from surrounding irrigated land. Calculations for fluxes were made from differences in watertable levels using Darcy’s Law, with values for saturated hydraulic conductivity (K s ) measured at the plantation by Feikema (1995). For vertical fluxes, K s = 0.001 m day−1 was used for the material above the aquifer through which most of the flow was occurring, and represented the zone of lowest K s and the greatest limit to flow. For lateral fluxes, K s = 0.01 m day−1 represented the conductive aquifer through which most of the water flow occurs.
Lateral gradients were calculated from periodic measurements of watertable height between piezometer nests at Site 2 (in the plantation) and Outside (in irrigated pasture outside the plantation) and vertical gradients were determined from using measured hydraulic gradients at Site 1. Observations of piezometric and watertable levels were supplemented by prior measurements made by Heuperman (1995). The distance over which the lateral gradients are calculated is 40 m. Given the steep gradient it is assumed that the watertable depth measured at Site 2 approximates the depth near (but not at) the edge of the plantation. Groundwater fluxes in the root zone were also estimated from changes watertable level (ΔH) at Site 1 using a specific yield (S) of 3% for clay (Johnson 1967).
Sampling and measurements
Samples for isotopic, soil water and solute measurements were collected in December 1995 (early summer), March 1996 (early autumn) and September 1996 (early spring). Characteristics of the three sampling sites (Sites 1, 2 and 3) are presented in Table 1. The sites were selected to represent a range of conditions within the plantation in terms of groundwater behaviour and soil profile descriptions given by Vertessy et al. (2000).
Samples of all potential water sources for plantation trees were collected from each site at each sampling time. These consisted of soil water at different depths, groundwater, and irrigation and drainage water from the surrounding paddocks. Rainfall (throughfall) samples were collected at two sites under the canopy at intervals between December 1995 and September 1996. Vegetation (twig) samples were also collected at each sampling time for extraction and analysis of water in the sapwood.
Given the practical difficulties of obtaining branches from tall trees, a single medium-sized tree was selected at each site, and samples were obtained from mid-canopy. At each sampling time, a shotgun was used to remove a branch from each tree, and 10–12 twigs of 1.0–1.5 cm diameter were taken. Twigs were quickly removed from the branch, cut into ∼8 cm lengths, and the bark and phloem scraped away to reduce contamination from isotopically enriched water. Twigs from each tree were divided into two subsamples and immersed in kerosene in an airtight 300 ml glass jar to prevent any isotopic exchange and to prepare for extraction of water by azeotropic distillation using kerosene (Thorburn et al. 1993b).
Soil samples were taken within 2 m from the sampled tree at each site using 5 cm and 10 cm diameter hand augers. Samples were collected at 10 cm intervals down to 40 cm (using a 10 cm auger), at 20 cm intervals to 100 cm and 50 cm intervals (using a 5 cm auger) down to the watertable (or to 5 m at Site 3). For reporting purposes, each depth range is represented by its respective midpoint value. Samples were placed in airtight 500 ml glass jars and analysed for isotopic signature, water content, chloride concentration and matric suction. Water for isotopic analysis was extracted from the soil samples by the azeotropic distillation method outlined by (Revesz and Woods 1990).
All water samples were analysed for deuterium by stable isotope mass spectrometry at the CSIRO Land and Water Adelaide Laboratory by reducing 25 μl of water to H2 over uranium at 800°C. Samples were measured with a V G Micromass mass spectrometer.
Oxygen isotope ratios (δ18O) in water were measured on selected samples using a modification of the Epstein and Mayeda (1953) technique as reported by Socki et al. (1990). Approximately 10 ml of water was evacuated in round sealable flasks for several minutes without letting the samples boil. Between 10 and 15 cm of CO2 was placed over the sample and allowed to equilibrate overnight. The equilibrated CO2 was purified cryogenically to remove N2 and H2O. δ 18O values were measured with a Finnigan MAT 252 mass spectrometer relative to SMOW (Standard Mean Ocean Water). Working standard reference gases had been equilibrated against international standards.
Groundwater samples with EC values greater than 10 dS m−1 were first treated with NaF, placed in an oven at 60°C for 5 h, and then centrifuged to remove the precipitated MgF and CaF, as Mg and Ca may otherwise fractionate during mass spectrometry and contaminate (K. McEwan, pers. comm.). The samples were then distilled to remove NaCl from the solution, and then analysed for isotopic composition in the same manner as other water samples.
Isotopic concentrations are expressed using the standard delta notation for δ 2H and δ 18O relative to V-SMOW (Vienna-Standard Mean Ocean Water) using
where R is the ratio of the heavy to light isotope, i identifies the isotope sample and s the standard. The precision for sampling and analysis is ±1.3‰ for δ 2H and ±0.3‰ for δ 18O for both E. camaldulensis xylem water (Thorburn et al. 1993b) and for soil water (Revesz and Woods 1990).
The source of water in the sampled twigs was identified by comparing the isotopic signatures of possible water sources at each sampling time. The simple mixing model described by Thorburn (1993) was used to investigate the relationship between δ 2H and δ 18O and to determine the proportions of each water source in twig water. If the possible end members for the isotopic composition of tree sap are the δ2H and δ18O values of the groundwater and soil water, then the proportion of groundwater may be calculated using
where X is the δ18O value of the sample, Y is the δ2H value of the sample, and s, g and t refer to soil water, groundwater and tree sap respectively. The proportion of groundwater (G p) in the tree sap is then given by Thorburn and Walker (1993):
Gravimetric moisture content (g g−1) of each soil sample was determined on a 20 to 50 g subsample from its weight loss on drying for 24 h at 105°C. Soil chloride concentrations were determined using the dried samples from moisture content measurements, extracted by shaking for 1 h with 50 ml of 0.1 M Ba(NO3)2 (barium nitrate), and then being allowed to settle for 1 h before titration.
The chloride concentration of each extract was determined by colorimetrical titration with silver nitrate (AgNO3) (Taras et al. 1975) on an aliquot of the clear supernatant using an autotitrator (Orion 960 Autochemistry System). Total chloride was extracted from the dry soil by barium nitrate (BaNO3) and titrated against silver nitrate (AgNO3). Chloride concentration of the soil water (mg l−1) was determined by dividing the total chloride of each sample by its gravimetric moisture content. This transformation allows for computations on a volumetric basis, and for comparison with TDS (mg l−1) of water samples. Osmotic potential (MPa) was estimated from chloride concentrations (mg l−1) in soil water (assuming that all Cl−is present as NaCl, and that NaCl was the main solute contributing to osmotic potential) using
where the constant, 0.000127, is derived by taking the osmotic potential of 1 M NaCl (35.5 g l−1Cl) as 4.5 Mpa (Mensforth 1996).
Soil matric potential was determined for each sample using the filter paper technique (Greacen et al. 1989). Three replicate air dry Whatman No. 42 filter papers were placed in contact with soil in 500 ml air-tight glass sample jars and left to equilibrate at constant temperature for 7 days. The papers were weighed immediately, oven dried at 105°C for 24 h, and reweighed. Using known calibrations, the matric potential can be estimated from the water content of the filter papers (Greacen et al. 1989). Reported values are means of the three replicates in each sample. Total soil water potential reported is the sum of matric and osmotic potentials.
Measurements of groundwater depth in wells and piezometers was made periodically from July 1994 until August 1998. Each piezometer was first bailed and the sample taken a few minutes later. Samples from irrigation water and drainage water (when present) were also collected at numerous sites surrounding the plantation. Measurements of EC (dS m−1) on all rain and groundwater samples were made with a conductivity meter. These were then converted to TDS (mg l−1) by multiplying values of EC (dS m−1) by 640 (Shainberg and Oyster 1978).
Results
Climate during the sampling period
Climate leading up to the first sampling period in December 1995 was relatively dry. The last major rain event before sampling occurred on 6/11/95 (7.4 mm) with a minor event on 21/11/95 (1.2 mm). The end of summer, prior to the second sampling period in March 1996, was wetter and cooler than usual. Significant rainfall events occurred on 1/1/96 (19.4 mm), 24/1/96 (24 mm), 8-9/2/96 (14.2 mm), 28/2/96 (15.2 mm) and at the time of sampling on 6/3/96 (11.4 mm).
Rainfall was significantly higher than the average during the winter months leading up to the sampling time in September 1996. Major rainfall events occurred on 1/8/96 (11 mm), 7/8/96 (7.7 mm), 18-19/8/96 (13.6 mm), 23/8/96 (9.8 mm), and 27/8/96 (7.6 mm), with some scattered minor events (<2 mm). The total rain falling during the sampling period (December 1995–September 1996) was 443 mm. Annual rainfall for 1996 was 483 mm (long term mean 465 mm).
Water balance
Results from the monthly water balance for the Kyabram plantation are given in Fig. 2, in which residual water represents the monthly cumulative surplus or deficit after tree transpiration has been subtracted from rainfall. A positive residual (surplus) implies that rainfall exceeded transpiration, and while it does not imply that no soil or groundwater was used during a particular month, it does suggest that there was no net usage of soil and/or groundwater for that month. A negative residual (deficit) indicates that rainfall was lower than transpiration for that month, suggesting that trees used a total net amount of water from soil moisture and/or groundwater.

Monthly rainfall and residual water in the soil profile and average watertable level under the plantation. Residual water (mm) is water left over after the subtraction of tree transpiration from effective rainfall. A positive residual occurs when total rain exceeds tree water use for that month
Table 2 lists water budget values corresponding to the average, the lowest, and the highest rainfall for the fourteen 12 month periods generated using data from 1994 to 1996. The soil moisture within the profile under the plantation is assumed to be constant over each 12 month period. Effective rainfall is slightly over half of the actual rainfall. Over all 12 month periods, soil water storage was assumed to be identical at the beginning and end of the annual period, and effective rainfall was not sufficient to meet the trees’ water requirement, resulting in estimated groundwater use of between 5 and 68% (average 26%).
A linear relationship applied to the effective rainfall as a percentage of transpiration (to provide a measure of how much of the plantation’s water use can be derived from rainfall) and actual rainfall is given in Fig. 3. It is acknowledged that the linearity of this relationship is due to the high dependence of the separate 12 monthly periods used in the analysis. While the assumptions of the water balance made earlier restrict the applicability of this relationship, it provides a crude estimate of the rainfall required to meet the trees’ water requirements under present site conditions. Using the linear relationship in Fig. 3, an annual rainfall of ∼556 mm is required to meet the plantations annual water requirements. Using the long-term average rainfall for the area (465 mm), rainfall will (on average) meet ∼84% of the plantation’s water requirements.

Effective rainfall (total rainfall less canopy interception and soil evaporation) as a fraction of transpiration, plotted against actual rainfall at the Kyabram plantation for 14, 12 month periods between 1994 and 1996
Water fluxes
Maximum rises in stored groundwater of up to 0.5 mm day−1 (water depth equivalent) were calculated from watertable changes under the plantation at Site 1 (Fig. 4a). This peak flow reflects the difference between the flow rate into the root zone from the saturated zone and groundwater extraction by tree roots. Maximum net rates of reduction in stored groundwater of up to 0.2 mm day−1 were estimated. These generally occurred during summer in periods with low rainfall. Assuming an inflow rate of 0.5 mm day−1, peak extraction of groundwater by trees estimated using this method would be about 0.7 mm day−1.

a Change in groundwater depth (water depth equivalent) and b Vertical and lateral saturated groundwater fluxes into the plantation root zone with time. Vertical gradients are calculated at Site 1 (over a distance of 6.5 m), and lateral gradients are calculated between Site 2 and Outside (over 40 m)
Water fluxes into the plantation root zone laterally from surrounding irrigated land and vertically from the saturated zone beneath the plantation are shown over time in Fig. 4b. Both lateral and vertical gradients were positive (laterally and upwards) towards the plantation root zone at all times. Vertical hydraulic gradients under the plantation ranged between 0.12 and 0.25 m m−1 (upwards) with higher gradients during periods of low rainfall and high evaporation. These gradients led to estimated fluxes into the root zone of between 0.12 and 0.25 mm day−1. Lateral hydraulic gradients were lower, between 0.01 and 0.09 m m−1, leading to estimated lateral fluxes into the root zone in the range 0.1–0.9 mm day−1.
Stable isotopes
Values of δ2H and δ18O for rain water ranged between −57 and 7.2‰ (mean −33‰) and from −8.3 to 0.95‰ (mean −5.2‰) respectively. Although the maximum and minimum occurred during summer and winter respectively, there was no seasonal trend. The value of δ2H of rainfall is probably best related to the ambient air temperature at the time of rainfall (see Dawson 1993).
The isotopic composition of groundwater was uniform spatially, with depth and through time (Fig. 5). At Site 1, mean groundwater values for δ2H (between 4.0 and 10 m) were −29.5 ± 0.9, −29.2 ± 0.5 and −29.2 ± 1.0 for the three sampling times (±95% confidence intervals). At Site 2, mean groundwater values for δ 2H (between 3.5 and 9.0 m) were −30.1 ± 0.8, −29.4 ± 1.3 and −28.7 ± 0.8, and at Site 3, mean groundwater values for δ2H (between 5.0 and 9.0 m) were −28.6 (at 9.0 m), −29.6 (at 9.0 m) and −28.2 ± 1.9 for the three sampling times. Most of the profiles show a slight minimum in δ2H between 1 and 3 m (Fig. 5). This appears to be common in δ2H profiles of field soils (Allison et al. 1983) and values are similar to isotopic groundwater signatures measured elsewhere (for example Mensforth 1996).

Plots of δ2H profiles for Sites 1 (a–c), 2 (d–f) and 3 (g–i) at December 1995, March 1996 and September 1996. Vertical lines and ‘tree’ symbols indicate deuterium values for twig water. Horizontal dashed lines indicate watertable level
While more pronounced in summer and particularly at Sites 1 and 2, there was a trend across all sites and all sampling times of decreasing δ2H to a depth of 1–3 m, with values below these depths remaining constant or increasing slightly, to approach that of the groundwater. δ2H values of upper soil (0–0.1 m) had the greatest fluctuation of 37.2‰ (ranging between −3.3 and −40.5‰) during the sampling period. Evaporation during summer (December 1995–March 1996) enriched surface soil water at all sites, while rain and low evaporation during winter and spring led to relatively uniform deuterium profiles in the upper soil by September 1996.
Twig water δ 2H varied little temporally and spatially, ranging between −18.5 and −28.3‰ (Fig. 5). Deuterium in twig water of E. grandis at Site 1 most closely followed changes in groundwater, reaching a minimum by late summer and increasing until early spring. Here, the temporal variation of twig water was most positively correlated with groundwater (R 2 = +0.84) and most negatively (R 2 = −0.88) with upper soil water (0–0.3 m depth) for the three observations.
Deuterium in twig water of E. grandis at Site 2 and E. camaldulensis at Site 3 was highest by the end of summer, and decreased until early spring. Temporal variation of twig water at Sites 2 and 3 both showed positive correlations with upper soil water (R 2 = +0.90 and +0.56 respectively) and negative correlations with groundwater (R 2 = −0.58 and −0.73 respectively).
The Local Meteoric Water Line (LMWL) for Kyabram does not differ significantly from the Global Meteoric Water Line (GMWL) (Craig 1961). The LMWL is included in the individual δ2H–δ18O plots (Fig. 6). Plots of deuterium (δ2H) against oxygen-18 (δ18O) are shown in Fig. 6. All δ2H and δ18O values from tree sap, soil water, groundwater and surface water lie to the right of the LMWL, indicating that the samples were evaporatively enriched relative to rainfall. At Site 1, twig (tree) water was most similar to soil water from depths of 0.25 and 0.35 m at sampling in December 1995. Twig water was similar to soil water at 0.15 m depth at Site 2 at this time. Unfortunately, twig water at Site 3 was very similar in isotopic composition to soil water deeper than 0.25 m and groundwater, making it impossible to distinguish between them as possible water sources.

Plots of δ2H and δ18O for twig, rain, surface, soil and groundwater samples at Site 1 (a–c), 2 (d–f) and 3 (g–i). (irrig irrigation water; drain drainage water; S15 soil at 15 cm depth; GW groundwater; Tree twig water, • = recent rainfall; dashed line represents local meteoric water line)
At the sampling time in March 1996, the composition of twig water at all three sites had a closer relationship with water higher in the profile. Twig water at Site 1 was closest to soil water at 0.15 m depth, whereas twig water from trees at Sites 2 and 3 fell between soil water from 0.05 and 0.15 m depth. While the majority of profiles are typical for a dry area, profiles in winter showed little difference in deuterium profiles, preventing identification of possible water sources.
Soil water, matric potential and chloride
Electrical conductivity of rainfall appeared to be lower during the winter months with a weighted average of 0.18 dS m−1 over the monitoring period. Assuming the majority of salts exist as NaCl, then the average salt concentration is 118 mg l−1 TDS, (∼72 mg l−1 Cl−) and equates to 5.8 kg ha.year−1 NaCl (13.9 kg year−1 over the plantation area of 2.4 ha).
Figure 7 shows soil matric and osmotic potentials for Sites 1, 2 and 3 at the three sampling times. Vertical lines indicate minimum tree twig water potentials measured during summer (February 1995) and spring (October 1995) by Morris et al. (1998). The water potential of the twig, as opposed to that of the leaf, more closely represents the suction exerted by the roots, and we assume that trees are unable to extract appreciable amounts of water from regions where the total soil water potential (matric and osmotic) is below that of the tree twig water potential. The soil profile was dominated by matric potential at all sites and at all times, with the osmotic potential component generally being very small (Fig. 7). Osmotic potentials (approximately −1 MPa with soil solution salinity approaching 10,000 mg l−1) were significant enough to decrease appreciably the total soil water potential near the watertable at Sites 1 and 2 (and possibly also at Site 3, although no measurements were made in this part of the profile).

Matric, osmotic and total soil water potential profiles for Sites 1 (a–c), 2 (d–f) and 3 (g–i) at each sampling period (December 1995, March 1996 and September 1996). The Total soil water potential ( ) is the sum of matric (
) is the sum of matric ( ) and osmotic (
) and osmotic ( ) potentials at each depth interval. Straight vertical lines indicate approximate minimum leaf water potentials (Morris et al. 1998)
) potentials at each depth interval. Straight vertical lines indicate approximate minimum leaf water potentials (Morris et al. 1998)
In December 1995, all three profiles showed low matric potentials near the soil surface. Recent rainfall increased upper soil matric potentials at the end of summer (March 1996). By the end of spring, the upper soil profiles were relatively wet, with soil water potentials approaching zero. Nonetheless, a zone of low soil water potential (−1.5 to −4 MPa) was observed at all sites at all times between the upper soil and groundwater. At the end of winter, this dry zone peaked at −2.8, −3.0 and −1.9 MPa at Sites 1, 2 and 3 respectively.
Soil water variation was greatest near the soil surface due to the influences of rainfall and evapotranspiration (Fig. 8). While the rest of the profile at Site 1 appeared to be driest by the end of summer, there was little change with time in the deep profiles at Sites 2 and 3.

Soil gravimetric soil moisture (a–c) and chloride mass (d–f) profiles for Sites 1, 2 and 3 at the three sampling times. Note different scales for soil chloride for each site
Soil chloride profiles provide evidence of two salt bulges in the profile (Fig. 9). The first, with a peak between 0.5 and 1.0 m at all sites appears to develop towards the end of summer, and one may assume that this peak may disappear by the start of the summer, only to develop again towards the end of summer as before. This salt bulge peaked at 300–400 mg kg−1 at Site 1, approximately 300 mg kg−1 at Site 2, and approximately 240 mg kg−1 at Site 3. The solute concentration developed during summer, and was still evident to the same magnitude by early spring (September 1996). Unfortunately, due to the lack of additional sampling, it is not known whether the upper solute bulge was subsequently reduced to the same magnitude as observed in early summer (December 1995).

Profiles of solution chloride for sites 1, 2 and 3 at three sampling times. Values above the watertable are soil solution and below the watertable are groundwater EC, assuming all salts are present as Cl
The second bulge occurred lower down the profile, in the zone of seasonal watertable fluctuation (Fig. 9). The bulge is not evident in the profile of Site 3 because the watertable is lower than 5 m depth. At Sites 1 and 2, this bulge peaks at 1,000 and 1,500 mg kg−1 respectively.
Soil solution chloride was combined with calculations made from groundwater EC measurements to give the chloride solution profiles shown in Fig. 9. High and low watertable levels are also shown. The high salt bulge noted earlier in Fig. 8 can be seen to occur in the fluctuating watertable zone. In general, the salt bulge at the watertable is one order of magnitude higher than the bulge at about 1 m depth.
Identifying water sources
Data from isotopic analyses, soil water, soil matric potential and soil chloride concentration were examined to define regions in the profile likely to contribute most to tree water uptake at each site and at each time. Chloride accumulation, available soil water and isotopic signature with depth are provided in Table 3. Based on isotopic signatures, water potential and chloride distribution, each profile was compartmentalised into two sections; upper soil and groundwater, and each soil compartment was assigned a representative value for deuterium and oxygen-18 (Table 4). This allows the calculation of the percentage of groundwater found in twig water in each case using Eqs. 5–7. Most of the water was taken up in the depth range 0.2–2.0 m and groundwater in December 1995, from 0 to 0.8 m and groundwater in March 1996, and between 0 and 2 m in September 1996.
All sites were similar to one another in the relative proportions of groundwater used at each sampling time (Table 5). Most groundwater was used in December 1995, with no groundwater being used in September 1996. Site 3 showed the greatest variation in the proportion of groundwater used, with about 80% of transpiration derived from groundwater in December 1995, while no groundwater was used in September 1996.
Discussion
Water balance
The water balance investigation confirmed the hypothesis that the unirrigated trees at Kyabram were using groundwater to help meet their annual water requirements. Despite a relatively low LAI of between 1.2 and 1.8, canopy interception was approximately 30% at the site (Vertessy et al. 2000) and effective rainfall (rain less canopy interception and soil evaporation) represented about 52% of the total rainfall during this period. Effective rainfall was estimated to account for about 74% (range 32–95%) of the plantation’s annual water use over the 2 year period between June 1994 and May 1996. At no stage did annual rainfall completely account for plantation water use. These results support existing evidence from the watertable drawdown that the groundwater is used by the plantation (Heuperman 1995), with groundwater accounting for between 5 to 68% of annual transpiration (Table 2).
Assuming trees preferentially use rainfall when available, a linear relationship relating transpiration requirements to rainfall (Fig. 3) suggests that, based on the long term average rainfall of 465 mm/yr, approximately 84% of transpiration is met by rainfall. Groundwater contributes an average of approximately16% to transpiration annually over this period. Using annual transpiration rates measured by Morris et al. (1998) groundwater uptake rates are estimated to be 50 mm year−1 (0.14 mm day−1 on average). This annual groundwater uptake rate is low relative to higher annual uptake rates in the region reported in lighter soil types by Benyon et al. (2006) of between 120 and 380 mm year−1 for E. grandis, but is similar to uptake rates in heavier soils, where little or no groundwater was used (Benyon et al. 2006).
Within the region with clay soils where the hydraulic conductivity is similar (0.003 mm day−1), slightly higher annual uptake rates of 170–220 mm year−1 have been estimated for E. camaldulensis (Morris and Collopy 1999). However that plantation is younger (7 years) and water table shallower (0.7–3.0 m BNS) and groundwater uptake rates could be expected to decrease as the plantation draws down the watertable over time, as the ability of trees to utilise groundwater will decrease with increasing groundwater depth (Thorburn et al. 1995), particularly in heavy textured soils.
Low hydraulic conductivities of soil in the unsaturated zone (Feikema 1995) in particular limit the rate of water uptake from groundwater, which is closely related to the depth of the watertable (Thorburn et al. 1995). Watertable depth undergoes an annual cycle, and is highly dependent on plantation transpiration rates. Times when soil water storage declined or groundwater was used coincided with times of low rainfall and high evaporative demand.
Water fluxes
Lower hydraulic conductivity of the medium above the more conductive aquifer (Feikema 1995) resulted in lateral fluxes into the root zone being about twice those of vertical fluxes. While the uncertainty associated with these calculations restricts further detailed analysis, lateral boundaries of the plantation (6,900 m2 to 10 m depth) is less than 30% of the ground area (24,000 m2), and so the vertical flux is likely to contribute greater volume of water to the root zone. Therefore, most of the groundwater directly entering the root zone appears to enter from beneath the plantation rather than laterally from under surrounding pasture. However, it is the lateral flow within the deeper more conductive aquifer that provides a significant amount of water that will ultimately flow up into the root zone from directly below the plantation.
Changes in the volume of stored groundwater derived from changes in watertable level suggest that inflow into the plantation root zone (laterally and vertically) is in the order of 0.5 mm day−1 (August 1995; Fig. 4). Assuming that the inflow rate is reasonably constant throughout the year, lowering of the watertable at a peak rate of 0.2 mm day−1 equates to groundwater extraction by trees of approximately 0.7 mm day−1. Estimates of transpiration rates for the site rarely exceeded 1.5 mm day−1 (Vertessy et al. 2000), and so the groundwater uptake may account for up to approximately 50% of transpiration flux at any given time.
The observed rises in the watertable result from lateral and vertical inflow rates exceeding groundwater extraction by trees during the cooler, wetter months. Times when groundwater uptake by trees exceeded inflow rates was confirmed by a lowering in watertable levels during the summer months when rainfall is low and evaporation is high.
Water sources and availability
Profiles of matric suction and soil chloride identified two zones of water uptake; upper soil rain-derived water (0–1.0 m) and groundwater (including groundwater-derived soil water). Between these zones, matric potential is low, suggesting that little opportunity exists for downward leaching of water and solutes from the root zone. This region of low total soil potential (Fig. 7) is not evident in the profiles of moisture content (Fig. 8). This is presumably a reflection of the heavy clay soil under the plantation, in which matric suction increases steeply at the dry end of the soil moisture characteristic curve. This highlights the importance of measuring soil matric potential in addition to soil water content when identifying the water available to plants. Ideally, leaf water potentials should be measured at the same that soil water potentials were measured. However, we present the minimum leaf water potentials only, and they provide a means (when used in conjunction with other techniques) to help identify zones from which trees are possibly extracting water.
While rainfall is the major source of water for transpiration, direct use of rainfall by trees is not indicated by the stable isotope data. The reason for this is presumably that rainwater would have mixed with isotopically enriched upper soil water, creating water with a non-distinctive δ2H value. Deuterium profiles at Site 2 in March 1996 (data not shown) indicate that rainwater mixed with or displaced the existing soil water (in the 0–0.1 m depth zone) to give a similar δ2H value to the rain water itself. The δ2H value of rain water in the soil would have been further altered by subsequent evaporation prior to sampling. Hence, recharged soil water from recent rain water may well have been a source of water for the trees, even though there was no clear isotopic signature of rain in the tree xylem water.
The δ2H and δ18O profiles in the soil are reasonably constant over time, and so the profile could be considered as two compartments, and single isotope data can provide information on relative water uptake from these compartments (Walker and Richardson 1991; Thorburn 1993). Water in the upper soil zone is derived from rainfall, while lower soil water (with limited leaching) consists of groundwater-derived water moving upwards in response to tree water extraction.
The relatively low groundwater use at the end of summer, when water use is higher, suggests that the soil imposes a large resistance to uptake of water from shallow groundwater (at 3.5 m depth). Studies on plantations on lighter soil types have reporting higher groundwater uptake rates approaching 2 mm day−1 (e.g Thorburn and Walker 1993; Thorburn et al. 1995; Benyon et al. 2006), while those on heavier textured soils generally report uptake rates between 0 and 0.5 mm day−1 (e.g. Morris and Collopy 1999; Benyon et al. 2006).
Low groundwater uptake may result from solute concentrations in the vicinity of the watertable (which approach 10,000 mg l−1) where osmotic potentials are relatively low. The osmotic potentials are generally less negative than the matric potentials, and limited groundwater uptake results from low hydraulic conductivity of soil at the capillary fringe. The soil itself does not appear to pose a mechanical barrier to root growth, because roots have been found much deeper under the plantation (Vertessy et al. 2000). While the osmotic potentials may not be low enough to induce severe limitations on available water, the measured solute concentrations in the capillary fringe are quite high (∼25% of seawater), and other effects, including those associated with salt toxicity (unregulated salt uptake), low oxygen concentrations and nutrient imbalances. Growth rates for E. camaldulensis and E. grandis have been observed to decline by approximately 3 and 10% in a nearby field research trial with imposed root zone salinity (1.5 dS m−1; ∼1,000 mg l−1) similar to that at Kyabram (unpublished data).
Stable isotope analyses suggest trees were using most groundwater-derived water during early summer (13–80%), when rainfall was low and groundwater relatively shallow after spring. A wetter than usual summer resulted in trees using relatively small amounts of groundwater in early autumn (3–11%). The enriched deuterium composition of tree water by the end of summer suggests that trees must have been using appreciable amounts of upper soil water. It is unlikely that trees were using no groundwater in early spring because i) trees use rain water preferentially when available; ii) effective rainfall was greater than transpiration during this period; and iii) the watertable was rising.
Combining results of the proportion of groundwater used by trees with transpiration rates measured by Morris et al. (1998), estimates of groundwater uptake rates were 0.2–1.2 mm day−1 in December 1995 and 0.02–0.1 mm day−1 in March 1996. The peak uptake rate estimated using the water fluxes of 0.7 mm day−1 falls within this range. This range is similar to uptake rates reported for similar soil textures in the literature (e.g. Cramer et al. 1999; Morris and Collopy 1999).
The application of stable isotopes in this study was limited by the small range in δ values between the different water sources. Variation in δ2H of water sources amounted to ∼30‰ in summer and only ∼10‰ in winter. Small differences in the isotopic composition of different waters impose a limitation of the technique in south-eastern Australia, where variation between rainfall and groundwater may be small (Simpson and Herczeg 1991) by comparison with studies in the USA, where differences of up to 80‰ may occur (Ehleringer et al. 1991). Evaporation of water resident in soils is the main process leading to the different isotopic composition of possible water sources (Thorburn and Walker 1993) and allowed sufficient differentiation during periods of high evaporation to estimate groundwater uptake in this study.
Salinity and groundwater level
Watertable levels at the Kyabram plantation generally fall from August to March, although the extent and timing of the rise in watertable observed in autumn varies from year to year. At Site 1, the greatest fall in the watertable coincided with relatively high percentage of groundwater being used (Table 5). There was a lower rate of decline during March 1996, when lateral and vertical hydraulic gradients (and therefore influxes) were large and trees were using proportionally less groundwater, and when the watertable was at its lowest since July 1995. The watertable rose during periods of little or no groundwater use, during periods when rainfall accounted for nearly all of transpiration requirements.
Chloride solution profiles at both Sites 1 and 2 identified a small chloride bulge peaking at between 0.5 and 1.0 m. A second, and more prominent zone of chloride accumulation was observed in the zone of watertable fluctuation (Fig. 9). The study by Vertessy et al. (2000) identified zones of solute concentration coinciding with zones of high root length density. The authors inferred that regions of high solute concentration were a result of water extraction by roots in that zone.
Sustainability of plantations over shallow saline groundwater
The presence of shallow groundwater at this site provides a limited yet reliable water source and has facilitated greater growth rates than would otherwise have been observed (in this rainfall limited region) had groundwater been considerably deeper (>6 m). There are no data available for comparison within the region of growth rates without groundwater access or supplemental irrigation, but benefits of shallow groundwater in increasing growth has been observed in other studies. For example, Dawson (1993) showed that box elder trees (A. negundo) were initially restricted to using only surface water sources, with years of little radial growth correlating with years of lower rainfall and lower stream runoff. As the trees gained access to groundwater, their radial growth increased.
The longer term availability of water is the major factor affecting growth and survival of plantation trees in this region. The relatively low annual transpiration and the absence of a definite summer peak in transpiration suggest that water availability is limiting water use by unirrigated trees at this site (Morris et al. 1998; Feikema 2000). This is related to increased resistance in the root zone with a heavy soil, resulting from a low and saline watertable in combination with low rainfall. This appears to be the case with the limited ability of trees to utilise groundwater, and may present a serious constraint to growth during periods when atmospheric conditions are otherwise sufficient to allow growth. Thus, longer term growth, or the sustainable tree density of plantation trees, at Kyabram now largely depend on rainfall at the site.
The dry zone observed under the plantation throughout the year is evidence that leaching does not normally occur at this site. While this is partly related to the lack of water entering the system from above, it is largely a function of the heavy clay soil. Even under flooded conditions, heavy clay soils may limit the effects of leaching and the subsequent benefits of increased transpiration and water status (Akeroyd et al. 1998; Feikema 2000). Heavy soil textures play an important role at Kyabram, where periodic leaching that would greatly enhance tree growth is prevented. Periodic leaching is an important process for long term tree survival in many natural ecosystems (for example, Mensforth 1996).
Groundwater at Kyabram has made a significant contribution to tree growth in what is a relatively dry environment. The availability of a perennial water source such as groundwater allows plants the opportunity to shift their water use to a more stable source (White et al. 1985). Trees at Kyabram appear to be opportunistic in using fresh rainwater when it is available, and permanent groundwater when rainfall is insufficient to meet transpiration demand.
Measurements of matric potential show a region of relatively low matric potential (ie. dry zone) flanked by two regions (an upper and lower) of higher potentials (Fig. 7). During periods of sufficient rain to maintain high matric potential in the upper profile, trees preferentially use rain-derived soil water. As this upper soil zone dries out during periods of little or no rain, the matric potential can fall so that it has a lower total hydraulic potential than the lower soil zone. In this situation, given that rooting depth is sufficient, trees may preferentially use water from the lower (groundwater-derived) zone during periods of little or no rain.
This is similar to other studies reporting the ability of Eucalyptus species to adapt readily and exploit changing water sources (Thorburn et al. 1993a; Mensforth et al. 1994; Costelloe et al. 2008). This rapid adaptation mechanism can, in large part, be attributed to the deep root system observed under the trees. This rooting system appears to be typical of Eucalyptus spp., and allows them to be opportunistic in their search for, and utilisation of, water and nutrients (Knight 1998).
The accumulation of solutes in the root zone, while possibly one of the most discussed processes likely to limit plantation growth and water use, may play a minor role in decreasing groundwater uptake by the plantation. Zones of chloride accumulation (with relatively low peaks) were evident at ∼0.6 m and at the watertable with relatively high peaks in concentration (Fig. 9). While solute storage in the root zone has increased over the life of the plantation (Heuperman 1995), an equilibrium may have been reached, whereby solute exclusion at the root interface is counteracted by mechanisms moving solutes within, and out of, the root zone. The zone of chloride accumulation appears to identify zones of current maximum water uptake, and is capable of relatively rapid dissipation and reformation in response to root activity linked to changes in watertable depth (Vertessy et al. 2000).
The presence of zones of lower salinity within the profile may be important for tree growth. Thompson (1988) suggested that the presence of zones of low salinities (<4 mg l−1 ECe), even when surface soil salinities were relatively high, gave E. camaldulensis the ability to survive in what were relatively saline soils. Temporary reprieve of stressful environments (e.g. leaching of solutes, see Jolly et al. 1993) may allow some eucalypts to survive in otherwise marginal areas, where chloride in the soil solution can reach up to 30,000 mg l−1 (Costelloe et al. 2008).
There have been isolated cases in which active roots have been found in the permanently saturated zone. Roots of E. globulus and E. cladocalyx have been found extending well into an unconfined aquifer by 3–5 m below the highest annual water level (Greenwood et al. 1992) where they survived prolonged waterlogging because of an oxygen content above the threshold for root metabolism. Heuperman (1995) also observed live roots to depths greater than 10 m (about 5 m below the highest annual water level) and acknowledged that the ability for tree roots to ‘by-pass’ saline layers would provide a mechanism to access deeper, less saline groundwater in the saturated profile. Roots have been observed below the watertable at Chowilla (Akeroyd et al. 1998).
Solute accumulation in the root zone at Kyabram exerts its major influence on tree growth towards the end of summer, when trees are increasingly reliant on a decreasingly available (i.e.deeper) groundwater resource; it is being lowered and solutes accumulate in the capillary fringe. During this time, resulting decreases in osmotic potential in the capillary fringe above the watertable are sufficient to reduce appreciably the total soil water potential.
Commercial plantation establishment in areas receiving less than 600 mm rainfall annually is normally not considered viable, unless trees have access to additional water sources such as groundwater to supplement their transpirational needs. The zone of predominant tree water extraction can change from deep to surface soil layers, depending on recent precipitation events (White et al. 1985; Mensforth and Walker 1996b; Chimner and Cooper 2004). However, there is evidence that the response of trees to changing root zone conditions may be unexpected; for example, where E. largiflorens does not use any relatively fresh water in surface soils in preference to shallow saline groundwater, or where stream side E. camaldulensis relies heavily on saline groundwater (Thorburn 1993).
While rates of groundwater uptake at Kyabram are low, and may not provide much scope for increasing discharge from shallow groundwater on heavy textured soils within the region, they may provide a means of utilising land that may otherwise be unsuitable for agricultural crops. The opportunity to enhance groundwater discharge by pumping groundwater and then irrigating plantations may also have limited benefits, as it is likely that salts will accumulate in the absence of leaching.
Conclusions
This study incorporated complementing techniques, including a water balance and stable isotopes, to investigate tree water sources used by unirrigated plantation trees. In relation to the initial hypotheses, unirrigated trees do use appreciable amounts of groundwater. While average uptake rates are low, they may comprise up to 80% of transpiration at times, and therefore provide a stable water source when rainfall is low. Groundwater becomes increasingly less available towards the end of summer, when the watertable depth and solute concentrations increase. This results in water being scarce in late summer-early autumn before autumn rain arrives. The main conclusions from this study are:
-
1.
There are two main sources of water for the plantation (rain and groundwater), with rainfall providing about 84% of the annual tree transpiration (at age 22 years).
-
2.
Upper soil water (from rainfall) is preferred by plantation trees when it is available.
-
3.
The dry zone at 1–2 m depth between the upper and lower water sources is subject to little or no leaching by infiltrating rain.
-
4.
Groundwater uptake by unirrigated plantation trees was estimated to be 0.2–1.2 mm day−1 in December 1995 and 0.02–0.1 mm day−1 in March 1996. Annual average groundwater uptake from the water balance study was approximately 50 mm year−1.
-
5.
Seasonal rise in the watertable observed under the plantation is explained by lateral and vertical fluxes into the root zone along hydrostatic gradients, and not by vertical recharge down through the plantation root zone.
-
6.
During periods of low rainfall and high evaporation, tree water use is closely related to (and limited by) groundwater depth.
References
Akeroyd MD, Tyerman SD, Walker GR, Jolly ID (1998) Impact of flooding on the water use of semi-arid riparian eucalypts. J Hydrol 206:104–117
Allison GB, Barnes CJ, Hughes MW, Leaney FWJ (1983) Effect of climate and vegetation on oxygen-18 and deuterium profiles in soils. In: Isotope hydrology 1983. Proc. Int. Symp. on Isotope Hydrology in Water Resource Development, Sept. 1983. IAEA, Vienna, pp 105–123
Bari MA, Ruprecht JR (2003) Water yield responses of south–west Western Australia forests, Water and Rivers Commission, Water Resource Technical Series. No. SLUI31
Bari MA, Schofield NJ (1991) Effects of agroforestry-pasture associations on groundwater level and salinity. Agrofor Syst 16:13–31
Bari MA, Schofield NJ (1992) Lowering of a shallow, saline water table by extensive eucalypt reforestation. J Hydrol 133:273–291
Bell RW, Schofield NJ, Loh IC, Bari MA (1990) Groundwater response to reforestation in the Darling Range of Western Australia. J Hydrol 119:179–200
Benyon RG, Marcar NE, Crawford DF, Nicholson AT (1999) Growth and water use of Eucalyptus camaldulensis and E. occidentalis on a saline discharge site near Wellington, NSW, Australia. Agric Water Manage 39:229–244
Benyon RG, Theiveyanathan S, Doody TM (2006) Impacts of tree plantations on groundwater in south-eastern Australia. Aust J Bot 54:181–192
Biddiscombe EF, Rogers AL, Greenwood EAN, DeBoer ES (1985) Growth of tree species near salt seeps, as estimated by leaf area, crown volume and height. Aust For Res 15:141–154
Brunel J-P, Walker GR, Walker CD, Dighton JC, Kennett-Smith AK (1990) Using stable isotopes of water to trace plant water uptake. In: Stable isotopes in plant nutrition, soil fertility and environmental studies. Proceedings of an International Conference, IAEA, Vienna, pp 543–551
Brunel J-P, Walker GR, Kennett-Smith AK (1995) Field validation of isotopic procedures for determining sources of water used by plants in a semi-arid saline environment. J Hydrol 167:351–368
Chimner RA, Cooper DJ (2004) Using stable oxygen isotopes to quantify the water source used for transpiration by native shrubs in the San Luis Valley, Colorado U.S.A. Plant Soil 260:225–236
Costelloe JF, Payne E, Woodrow IE, Irvine EC, Western AW, Leaney FW (2008) Water sources accessed by arid zone riparian trees in highly saline environments, Australia. Oecologia 156:43–52
Craig H (1961) Isotopic variations in meteoric waters. Science 133:1702–1703
Cramer VA, Thorburn PJ, Fraser GW (1999) Transpiration and groundwater uptake from farm forest plots of Casuarina glauca and Eucalyptus camaldulensis in saline areas of southeast Queensland. Agric Water Manage 39:187–204
Dawson TE (1993) Water sources of plants as determined from xylem-water isotopic composition: perspectives on plant composition, distribution and water relations. In: Ehleringer JR, Hall AE, Farquhar GD (eds) Stable isotopes and plant carbon-water relations. Academic Press, San Diego, pp 465–492
Ehleringer JR, Dawson TE (1992) Water uptake by plants: perspectives from stable isotope composition. Plant Cell Environ 15:1073–1082
Ehleringer JR, Phillips SL, Schuster WFS, Sandquist DR (1991) Differential utilisation of summer rains by desert plants. Oecologia 88:430–434
Engel R (1986) Agroforestry management of saltland using salt tolerant eucalypts. Trees Nat Resour 28:12–13
Epstein S, Mayeda T (1953) Variation of O18 content of waters from natural sources. Geochim Cosmochim Acta 4:213–224
Farrington P, Salama RB (1996) Controlling dryland salinity by planting trees in the best hydrological setting. Land Degrad Dev 7:183–204
Farrington P, Turner JV, Gailitis V (1996) Tracing water uptake by jarrah (Eucalyptus marginata) trees using natural abundances of deuterium. Trees 11:9–15
Feikema PM (1995) Description of soil water and shallow groundwater under a eucalypt plantation near Kyabram, Victoria. MSc thesis, School of Earth Sciences, The University of Melbourne, Victoria, 130 p
Feikema PM (2000) Tree growth and water relations in eucalypt plantations over saline shallow groundwater. PhD thesis, Department of Civil Engineering Monash University, 238 pp
Fraser GW, Thorburn PJ, Cramer V (1995) Comparison of diffuse groundwater discharge from trees and bare soils in south east Queensland: Preliminary results. In: Murray Darling 1995 Workshop. Environmental Geoscience and Groundwater Division, Australian Geological Survey Organisation, Wagga Wagga, NSW, pp 100–103
George RJ (1990) Reclaiming sandplain seeps by intercepting perched groundwater with eucalypts. Land Degrad Rehabil 2:13–25
George RJ (1991) Management of sandplain seeps in the wheatbelt of Western Australia. Agric Water Manage 19:85–104
George RJ, Nulsen RA, Ferdowsian R, Raper GP (1999) Interactions between trees and groundwaters in recharge and discharge areas—A survey of Western Australian sites. Agric Water Manage 39:91–113
Ghassemi F, Jakeman AJ, Nix HA (1995) Salinisation of land and water resources. Human causes, extent, management and case studies. University of New South Wales Press Ltd, Sydney, 526 p
Greacen EL, Walker GR, Cook PG (1989) Procedure for filter paper method for measuring soil water suction. CSIRO Division of Soils, Canberra
Greenwood EAN (1986) Water use by trees and shrubs for lowering saline groundwater. Reclam Reveg Res 5:423–434
Greenwood EAN, Milligan A, Biddiscombe EF, Rogers AL, Beresford JD, Watson GD, Wright KD (1992) Hydrologic and salinity changes associated with tree plantations in a saline agricultural catchment in southwestern Australia. Agric Water Manage 22:307–323
Heuperman AF (1995) Salt and water dynamics beneath a tree plantation growing on a shallow watertable. Institute of Sustainable Agriculture, Tatura Centre. Department of Agriculture, Energy and Minerals Victoria, 61 p
Heuperman AF, Stewart HTL, Wildes RA (1984) The effect of eucalypts on water tables in an irrigation area of northern Victoria. Water Talk 52:4–8
Jenkin JJ (1981) Terrain, groundwater and secondary salinity in Victoria, Australia. Agric Water Manag 4:143–171
Johnson AI (1967) Specific yield—compilation of specific yields for various materials. US Geological Survey Water Supply Paper 1662-D, 74 p
Jolly ID, Walker GR (1996) Is the field water use of Eucalyptus largiflorens F Muell affected by short-term flooding? Aust J Ecol 21:173–183
Jolly ID, Walker GR, Thorburn PJ (1993) Salt accumulation in semi-arid floodplain soils with implications for forest health. J Hydrol 150:589–614
Khanzada AN, Morris JD, Ansari R, Slavich PG, Collopy JJ (1998) Groundwater uptake and sustainability of Acacia and Prosopis plantations in southern Pakistan. Agric Water Manage 36:121–139
Knight JH (1998) Root distributions and water uptake patterns in eucalypts and other species: a literature review. In: Landsberg J (ed) The ways trees use water, Rural Industries Research and Development Corporation, ISBN 064278117
Mahmood K, Morris J, Collopy J, Slavich P (2001) Groundwater uptake and sustainability of farm plantations on saline sites in Punjab provine, Pakistan. Agric Water Manage 48:1–20
McJannet DL, Vertessy RA, Tapper NJ, O’Sullivan SK, Beringer J, Cleugh H (1996) Soil and litter evaporation beneath re-growth and old-growth mountain ash forest. Cooperative Research Centre for Catchment Hydrology, 59 p
Mensforth LJ (1996) Water use strategy of Melaleuca halmatorum in a saline swamp. PhD thesis, Botany Department, University of Adelaide, 239 p
Mensforth LJ, Walker GR (1996a) Root dynamics of Melaleuca halmaturorum in response to fluctuating saline groundwater. Plant Soil 184:75–84
Mensforth LJ, Walker GR (1996b) Water use of Melaleuca halmaturorum in a saline swamp. In: National conference on the productive use and rehabilitation of saline lands. Promaco Conventions Pty Ltd, Albany, Western Australia, pp 279–287
Mensforth LJ, Thorburn PJ, Tyerman SD, Walker GR (1994) Sources of water used by riparian Eucalyptus camaldulensis overlying highly saline groundwater. Oecologia 100:21–28
Morris JD, Collopy JJ (1999) Water use and salt accumulation by Eucalyptus camaldulensis and Casuarina cunninghamiana on a site with shallow saline groundwater. Agric Water Manage 39:205–227
Morris JD, Thompson LAJ (1983) The role of trees in dryland salinity control. Proc R Soc Vic 95:123–131
Morris J, Mann L, Collopy J (1998) Transpiration and canopy conductance in a eucalypt plantation using shallow saline groundwater. Tree Physiol 18:547–555
Northcote KH (1979) A Factual Key for the Recognition of Australian Soils (4th ed). Rellim Technical Publications, Adelaide, South Australia
Prenzel J (1979) Mass flow to the root system and mineral uptake of a beech stand calculated from 3-year field data. Plant Soil 51:39–49
Revesz K, Woods PH (1990) A method to extract soil water for stable isotope analysis. J Hydrol 115:397–406
Schofield NJ (1990) Determining reforestation area and distribution for salinity control. Hydrol Sci 35:1–19
Schofield NJ, Loh IC, Scott PR, Bartle JR, Ritson P, Bell RW, Borg H, Anson B, Moore R (1989) Vegetation strategies to reduce stream salinities of water resource catchments in south-west Western Australia. Water Authority of Western Australia
Shainberg I, Oyster JD (1978) Quality of irrigation water. International Irrigation Information Centre, Bet Dagan
Simpson HJ, Herczeg AL (1991) Salinity and evaporation in the River Murray basin, Australia. J Hydrol 124:1–27
Skene JKM, Poutsma TJ (1962) Soils and land use in part of the Goulburn Valley, Victoria, comprising the Rodney, Tongala-Stanhope, North Shepparton and South Shepparton irrigation areas. Technical Bulletin No. 14. Department of Agriculture, Victoria, Australia
Socki RA, Karlsson HR, Gibson EK (1990) A rapid, inexpensive technique for analysis of 18O/16O in water using pre-evacuated containers. In: Proc. Seventh Int. Conf. Geochronolog. Cosmochronol. Isot. Geol, p. 49. Aust. Bur. Min. Res., Canberra
Stirzaker R (2002) Planting trees over shallow, saline watertables. In: Vertessy R, Silberstein R, Sarre A (eds) Trees, water and salt: an Australian guide to using trees for healthy catchments and productive farms. RIRDC/LWRRDC/FWPRDC Joint Venture Agroforestry Program, Canberra, pp 94–109
Taras MJ, Greenberg AE, Hoak RD (1975) Standard methods for the examination of water and wastewater. American Public Health Association, Washington, pp 613–614
Taylor PJ (1998) The potential impacts of interception belts on the management of dryland salinity PhD thesis, Department of Agronomy and Farming Systems, Faculty of Agricultural and Natural Resource Sciences, University of Adelaide
Thompson LAJ (1988) Salt tolerance in Eucalyptus camaldulensis and related species. PhD thesis, School of Agriculture and Forestry, University of Melbourne, 307 p
Thorburn PJ (1993) Groundwater uptake by Eucalyptus forests on the Chowilla floodplain, South Australia: implications for groundwater hydrology, and forest ecology and health. PhD thesis, School of Earth Sciences, The Flinders University of South Australia, 191 p
Thorburn PJ, Ehleringer JR (1995) Root water uptake of field-growing plants indicated by measurements of natural-abundance deuterium. Plant Soil 177:225–233
Thorburn PJ, Walker GR (1993) The source of water transpired by Eucalyptus camaldulensis: soil, groundwater or streams? In: Ehleringer JR, Hall AE, Farquhar GD (eds) Stable isotopes and plant carbon-water relations. Academic Press, San Diego, pp 511–527
Thorburn PJ, Hatton TJ, Walker GR (1993a) Combining measurements of transpiration and stable isotopes of water to determine groundwater discharge from forests. J Hydrol 150:563–587
Thorburn PJ, Walker GR, Brunel J-P (1993b) Extraction of water from Eucalyptus trees for analysis of deuterium and oxygen-18: laboratory and field techniques. Plant Cell Environ 16:269–277
Thorburn PJ, Walker GR, Jolly ID (1995) Uptake of saline groundwater by plants: an analytical model for semi-arid and arid areas. Plant Soil 175:1–11
Vertessy RA, Connell LD, Morris JD, Silberstein RP, Heuperman AF, Feikema PM, Mann L, Komarzynski M, Collopy JJ, Stackpole D (2000) Sustainable hardwood production in shallow watertable areas. Rural Industries Research and Development Corporation, Land and Water Research and Development Corporation, Forest and Wood Products Research and Development Corporation, 105 p
Walker CD, Brunel J-P (1990) Examining evapotranspiration in a semi-arid region using stable isotopes of hydrogen and water. J Hydrol 118:55–75
Walker CD, Richardson SB (1991) The use of stable isotopes of water in characterising the source of water in vegetation. Chem Geol (Isot Sect) 94:145–158
White JWC, Cook ER, Lawrence JR, Broecker WS (1985) The D/H ratios of sap in trees: implications for water sources and tree rings D/H ratios. Geochim Cosmochim Acta 49:237–246
Williamson DR (1986) The hydrology of salt affected soils in Australia. Reclam Reveg Res 5:181–186
Acknowledgments
The authors would like to thank Prof Russell Mein, Dr Glen Walker and Alfred Heuperman for their input to this study, and Dr Ian Cartwright, Ms Mary Jane, Ms Kerryn McEwan, Ms Megan Eaterbrook, and Messrs Andrew Holub, Fred Leaney and John Dighton for assistance with stable isotope analysis. Ms Louise Mann provided technical support, and Peter Clayton assisted with obtaining twig samples from tree canopies. We thank Dr M. Bari and two anonymous reviewers for constructive comments on earlier drafts. The first author acknowledges the assistance of an Australian Postgraduate Award. This work was funded by the Land and Water Resources Research and Development Corporation, and Monash University.
Author information
Authors and Affiliations
Corresponding author
Additional information
Responsible Editor: John McPherson Cheeseman.
Jim D. Morris - Deceased.
Rights and permissions
About this article
Cite this article
Feikema, P.M., Morris, J.D. & Connell, L.D. The water balance and water sources of a Eucalyptus plantation over shallow saline groundwater. Plant Soil 332, 429–449 (2010). https://doi.org/10.1007/s11104-010-0309-2
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11104-010-0309-2