
Abstract
Overgrazing significantly affects alpine meadows in ways similar to grasslands in other areas. Fencing to exclude grazers is one of the main management practices used to protect alpine meadows. However, it is not known if fencing can improve soil properties and soil organic carbon storage by restraining grazing in alpine meadows. We studied the long-term (nine-year) effects of fencing on soil properties, soil organic carbon and nitrogen storage compared with continued grazing in an alpine swamp meadow of the Qinghai–Tibetan Plateau, NW China. Our results showed that fencing significantly improved vegetation cover and aboveground biomass. There were significant effects of fencing on pH value, soil bulk density, and soil moisture. Long-term fencing favored the increase of soil total nitrogen, soil organic matter, soil organic carbon, soil microbial biomass carbon and soil carbon storage compared with grazed meadows. Our study suggests that long-term fencing to prevent disturbance could greatly affect soil organic carbon and nitrogen storage with regard to grazed meadows. Therefore, it is apparent from this study that fencing is an effective restoration approach of with regard to the soil’s storage ability for carbon and nitrogen in alpine meadow of the Qinghai–Tibetan Plateau.
Similar content being viewed by others


Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
Soils play an important role in the carbon and nitrogen cycles and storage in grassland ecosystems (Tiessen et al. 1994; Neill et al. 1997; Lal 2004; Piao et al. 2009). Soil carbon storage represents a significant sink of carbon from the atmosphere (Neill et al. 1997), and changes to surface soil organic carbon and nitrogen stocks and cycling patterns are also important indicators of soil fertility and utilization potential (Tiessen et al. 1994). Human alteration of natural ecosystems for agriculture is an important component of disturbance, and has the potential to alter soil carbon and nitrogen storage.
The grassland ecosystem is an important carbon-sink or source and soils are the largest source of uncertainty in the terrestrial carbon balance (Piao et al. 2009). Grassland ecosystems can influence global environmental change through their strong potentials for carbon and nitrogen sequestration (Wright et al. 2004). But, overgrazing may reduce soil carbon sequestration and storage in grassland ecosystem (Shrestha and Stahla 2008) and forest ecosystem (Abril et al. 2005). Soil management of ecosystems also can influence the rate of soil organic carbon sequestration (Lal 2004). Improved grazing methods have been considered as an important strategy to renew degraded grassland and to increase the soil carbon pool (Lal 2004). The exclusion of livestock through the use of mesh fencing to create large-scale enclosures has become a common grassland management strategy throughout the world in recent decades (Reeves 2000; Smith et al. 2000; Shrestha and Stahla 2008; Wu et al. 2009). Fencing to exclude stock has been widely regarded as a simple restoration method for degraded grassland and the fenced remnants have typically been thought to ‘look after themselves’ (Reeves 2000). Recent research has focused on the effects of fencing on aboveground vegetation succession and community structure (Pettit et al. 1995; Gibson et al. 2001; Spooner et al. 2002; Wu et al. 2009). Such management measures present a dilemma between grazing utilization and biodiversity protection for grassland (Smith et al. 2000). However, less information is available on effects of long-term fencing on soil organic carbon and nitrogen stocks in a grassland ecosystem, especially in alpine swamp meadow of the Qinghai–Tibetan Plateau.
In this paper, we examine changes in soil organic carbon, nitrogen stocks and other soil properties of a nine-year fenced swamp meadow relative to grazed meadow. The aim of this study was to better understand the effects of excluding grazing herbivores through fencing on high altitude meadow soil in the Qinghai–Tibetan Plateau and to determine whether long-term fencing can significantly advance soil carbon sequestration and storage as a grassland management tool. Specifically we examined the effects of long-term fencing on: (1) soil properties; and (2) soil organic carbon and nitrogen storage. Our long-term hope is that the study should contribute to the restoration of carbon and nitrogen sequestration ability in degraded grasslands in the Qinghai–Tibetan Plateau.
Methods
Study area
This experiment was conducted in alpine meadow at 3,500 m a.s.l. in the eastern Qinghai–Tibetan Plateau at Manrima village of the Maqu Wetland Protection Area (33°45′ N, 102°04′ E) in Gansu Province, PR China (Fig. 1). The mean daily air temperature is 1.2°C, ranging from −10°C in January to 11.7°C in July. Mean annual precipitation is 620 mm, mainly falling during the short, cool summer. The monthly mean temperature and precipitation, annual average precipitation and annual accumulated temperature (≥0°C) from 1969 to 2005 in Maqu County were reviewed in Wu et al. (2009). The annual cloud-free solar radiation is about 2,580 h. The vegetation is typical alpine meadow and is dominated by clonal Kobresia sp. (Cyperaceae), Festuca ovina, Poa poophagorum, Roegneria nutans, Agrostis sp. (Poaceae), Saussurea sp. (Asteraceae), and Anemone rivularis (Ranunculaceae) (Wu et al. 2009). The total Maqu Wetland Protection Area had 85.87 × 104 ha alpine meadow. The soil type of the study area was an alpine meadow soil.

Map of the Maqu Wetland Protection Area showing the location of the study sampling sites. It is located in the south of Gansu Province and lie in the eastern Qinghai–Tibetan Plateau, China. Red circles are sampling sites and open squares are sampling quadrats. Five blocks were selected respectively in grazed meadow and fenced meadow and five quadrats were determined for soil sampling collection in each block. Total 50 squares (1.0 m × 1.0 m) were included in this study
Experimental design and soil sampling
An alpine swamp meadow dominated by Kobresia tibetica, Carex muliensis and including grasses and some forbs was selected for this study. The dominant botanical components and their biomass, and litter biomass in fenced and grazed meadows are showed in Table 1. Grazed meadow had lower total biomass and higher plant density, but ungrazed meadow had higher total biomass and lower plant density (Wu et al. 2009). This showed that ungrazed meadow had the larger individuals than grazed meadow. We selected five 50 m × 50 m blocks in each the grazed and fenced meadows using the line transect method (Fig. 1). This resulted in a total of 10 blocks for the study. Grazed meadows had become degraded because they were undergoing nomadic grazing from a medium density of Tibetan sheep and yaks (the approximate proportion of sheep vs yaks was about 1.6:1) all the time. And, fenced meadows were fenced from 1999 to now because of Natural Forest Protection Project of China. The fence completely excluded livestock grazing during the plant growth-season from April to October and slight grazing was done only during the hay-stage in winter.
We established five zonal random sampling quadrats (1 m × 1 m) per block in both the five grazed blocks and five fenced blocks. The quadrats were randomly arranged in every sampling block. Samples were taken in mid-August of 2008, when biomass had reached its highest. The covers, aboveground biomass and litter biomass were measured for these meadow communities. Meanwhile, we collected five soil samples by bucket auger within each quadrat from each sampling block in each the fenced and grazed blocks, then mixed five homogenised soil samples as a soil sample. Five mixed soil samples at depths of 0–30 cm in each sampling quadrat of each block were used to analyze soil properties. A total of 25 mixed soil samples were collected respectively for grazed meadow and fenced meadow.
All soil samples were air-dried and then passed through a 0.14 mm sieve. Soil pH was determined using a soil–water ratio of 1:5, soil water content before air drying was obtained by the oven-drying method, soil bulk density was measured using the soil cores (volume, 100 cm3) by the volumetric ring method (Soil Science Society of China 1983); soil total nitrogen and soil organic matter were measured by the K2Cr2O7 method using the modified Kjeldahl wet digestion procedure of Miller and Keeney (1982); the organic carbon content in the soil samples was measured using a Mebius method by the Walkley–Black acid digestion (Nelson and Sommers 1982); microbial biomass carbon was measured by the method of Vance et al. (1987). The content of each soil property was calculated by the proportions of its account for per soil dry weight. We calculated the total soil organic carbon storage density (TSOC, g cm−2) by the method of He et al. (2008) on a ground area basis up to a 30-cm depth as follows:
where Di, Pi, OMi, and S represent, respectively, the soil thickness (cm), bulk density (g cm−3), organic carbon concentration (%), and cross-sectional area (cm−2) of the ith layer; i = 1, 2, and 3.
Data analyses
All data were expressed as mean ± 1 standard error of mean. The mixed soil samples data for the 0–30-cm soil layer were used to analyze the soil properties and soil organic carbon storage potentials of the grazed and fenced meadows. ANOVA tests (followed by Tukey’s) were performed to test for differences of aboveground vegetation covers, aboveground biomass, litter biomass, soil properties and soil organic carbon storage between fenced and grazed meadows to assess the effects of long-term fence on aboveground and belowground properties and soil organic carbon storage. Significant differences for all statistical tests were evaluated at the level of P ≤ 0.05. All statistical analyses were performed using the software program SPSS, ver. 13.0 (SPSS Inc., Chicago, IL, USA).
Results
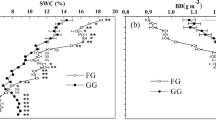
ANOVA analyses showed that the fenced meadows had greater total vegetation cover (F = 3.954, P < 0.001), aboveground biomass (F = 2.259, P < 0.001) and litter biomass (F = 1.433, P < 0.001) in comparison to the grazed meadows. Fencing the meadow for nine years increased respectively 72.94%, 57.66% and 98.42% on vegetation covers, aboveground biomass and litter biomass than grazed meadows (Table 1). Meanwhile, the long-term fencing significantly altered soil properties in alpine meadow. Long-term fencing of the meadows was associated with a reduction in pH from 7.67 to 7.20 (F = 4.828, P < 0.01) and in soil bulk density from 1.12 to 0.86 g cm−3 (F = 4.514, P < 0.001) compared with grazed meadows.
Long-term fencing of the meadows significantly increased soil water content (from 23.26% to 33.78%, F = 2.473, P < 0.001), soil total nitrogen (from 5.21 to 6.58 g kg−1, F = 1.327, P < 0.05), soil organic matter (from 106.09 to 158.58 g kg−1, F = 6.385, P < 0.001), soil organic carbon (from 62.94 to 77.44 g kg−1, F = 4.246, P < 0.01) and soil microbial biomass carbon (from 1.32 to 1.51 g kg−1, F = 2.647, P < 0.05) in the 0–30 cm soil depth compared with grazed meadows (Table 2). Nine-year-fencing of meadows significantly increased soil carbon storage in the surface 0–30 cm of soil from 2.97 in the grazed meadows to 3.55 kg m−2 (F = 3.541, P < 0.001) (Table 2), and the increased carbon storage due to reduced grazing for the area were estimated about 49.80 × 108 kg in 85.87 × 104 ha alpine meadow.
Discussion
Effects of fencing on ecosystem properties except soil carbon
Our results showed that the fenced meadow had higher aboveground biomass, litter biomass, vegetation cover, soil carbon, soil microbial biomass, soil moisture, soil nitrogen and soil organic matter, but a lower pH value and soil bulk density. The reduced disturbance had a direct effect on the dominant species of vegetation, vegetation cover and aboveground biomass, and reduced trampling may have resulted in a reduction in bulk density. Long-term trampling by livestock can lead to soil compaction, reduced soil infiltration rates, soil porosity, increased soil bulk density, and limited oxygen concentrations. These result in a decline of activity of edaphon in soil of grazed meadow (Holt 1997).
There is a buildup of litter in the ungrazed treatment due to the more grass and the less herbs, as offtake by herbivores is reduced. However, this build up has been slow suggesting that the increase in litter production may be offset by a more rapid litter decomposition rate in the moist, more neutral acidic and less dense soil of fenced meadow (Geissen and Guzman 2006). This may have contributed to the higher soil organic matter and carbon contents in the fenced areas.
Fenced meadows also had higher soil microbial biomass carbon than the grazed meadows; consistent with the results of Gu et al. (2007) which showed that overgrazing significantly reduced soil microbial biomass carbon in typical grasslands in Inner Mongolia of China. It has been interpreted by Iyyemperumala et al. (2007) that changes in soil properties provide a mosaic of environments for soil microbes and likely alters microbial ecophysiology and their mediated nutrient transformations. In addition, soil microbial biomass serves as a sink and source of N and thus plays an important role in regulating soil available N and preventing undesirable N loss from soil systems (Iyyemperumala et al. 2007). So, higher soil nitrogen contents may be related to higher soil microbial biomass carbon in fenced meadow. However, excretal returns of dung and urine from grazing animals can also result in the accumulation of soil nitrogen (Thomas et al. 2006). As well as increasing soil nitrogen, fencing also results in increased soil total phosphorus and available phosphorus (Wu et al. 2009).
Effects of fencing on soil carbon storage
Although aboveground vegetation plays an important role in regulating the biogeochemistry of ecosystems by fixing carbon and nutrients and preventing the loss of nutrients under disturbances (Bormann and Sidle 1990), it is clear from these results that management of alpine swamp meadows also has great impact on soil carbon and nitrogen storage. Fencing increased soil carbon sequestration and storage: with the assumption that past grazing has resulted in carbon losses from this system. Our results showed that soil organic carbon, soil microbial biomass carbon and soil carbon storage all increased after fencing for nine years; increases were of 23.04%, 14.39% and 19.53% compared to the grazed meadow, respectively. Similar studies have reported that there was positive contribution for soil carbon sequestration under exclosure conditions in degraded sandy grassland (e.g. Li et al. 2006).
There may be a range of potential mechanisms through which soil carbon increased in the fenced meadows. Firstly, the return of carbon from increased aboveground biomass and litter. As soil carbon input is mainly derived from litter, so that primary productivity is the main driver of soil carbon sequestration (De Deyn et al. 2008), even in alpine meadows of the Qinghai–Tibetan Plateau with a short growing season. Excluding grazing reduced the outflow of energy and nutrient from soil–plant system to consumers (livestock), especially for palatable grasses (Moretto and Distel 1997), Poaceae spp. and Cyperaceae spp. which are dominant species in this study area. Above-ground plant resources may be returned to the soil when litter layers decompose (Bardgett and Wardle 2003) in fenced meadow.
Secondly, belowground biomass (dead roots, mycorrhizae, and exudates) is an important element of soil carbon sequestration (Langley and Hungate 2003). Belowground biomass appears to be significantly decreased by grazing (Li et al. 2006), so grazing removal may potentially increase the amount of soil carbon sequestered in this way.
Thirdly, changes in vegetation composition and dominant plant functional group could affect soil carbon sequestration (De Deyn et al. 2008). The presence of C4 grasses in combination with other species has also been shown to increase soil C accumulation and biomass production in other grasslands (Fornara and Tilman 2008). So, we suggest the increase of C3 plant functional group species, Potentilla anserine, Elymus nutans, Kobresia humilis and decrease of C4 plant functional group species, Cyperaceae spp. and Poaceae spp. also may be a reason why there are higher soil C and N accumulation in fenced meadow soils than grazed meadow soils.
Grassland carbon management
Grasslands play an important role in the global carbon (C) cycle. Grassland in good condition should be in balance in terms of carbon input and carbon output or in the state where carbon input is greater than carbon output. At least one study has shown that carbon output is greater than carbon input in grazed grassland (Li et al. 2006). Our results indicate that fenced meadow results in higher soil carbon storage than grazed meadow. All these suggest that fencing is an effective restoration measure in that it favors of soil carbon sequestration and storage in this type of grassland.
Conclusions
Soil carbon, nitrogen and water content, soil organic matter and soil microbial biomass carbon and soil organic carbon storage were all increased by fencing meadow communities to prevent grazing. Higher soil organic carbon, soil total nitrogen and soil microbial biomass carbon storage may be attributed to the increase in plant aboveground vegetation and changes in soil physical characteristics after grazing is removed. Therefore, it is apparent from this study that fencing is an effective restoration approach of storage ability for soil carbon and nitrogen stocks in alpine meadow of the Qinghai–Tibetan Plateau. This may be true of other grassland without a long history of grazing. Although the soil sampling replications were sufficient for this alpine swamp meadow in this study, however, additional researches at more sites should be sampled to extend the findings in this study. However, it should be borne in mind that management for carbon may result in reductions in biodiversity, as both may not be maximized under the same management regime.
References
Abril A, Barttfelda P, Bucherb EH (2005) The effect of fire and overgrazing disturbes on soil carbon balance in the Dry Chaco forest. For Ecol Manag 206:399–405
Agriculture Chemistry Council, Soil Science Society of China (1983) General analysis methods of soil agriculture chemistry. Science, Beijing
Bardgett RD, Wardle DA (2003) Herbivore-mediated linkages between aboveground and belowground communities. Ecology 84:2258–2268
Bormann BT, Sidle RC (1990) Changes in productivity and distribution of nutrients in a chronosequence at Glacier Bay National Park, Alaska. J Ecol 78:561–578
De Deyn GB, Cornelissen JHC, Bardgett RD (2008) Plant functional traits and soil carbon sequestration in contrasting biomes. Ecol Lett 11:516–531
Fornara DA, Tilman D (2008) Plant functional composition influences rates of soil carbon and nitrogen accumulation. J Ecol 96:314–322
Geissen V, Guzman GM (2006) Fertility of tropical soils under different land use systems—a case study of soils in Tabasco, Mexico. Appl Soil Ecol 31:169–178
Gibson RS, Hewitt A, Sparling G, Bosch OJH (2001) Vegetation change and soil quality in central. Otago Tussock grasslands, New Zealand. Range J 22:190–204
Gu XJ, Zhao J, Wang J (2007) Soil microbial biomass of typical grassland in Inner Mongolia. J Agro-Environ Sci 26:1444–1448, in Chinese with English abstract
He NP, Yu Q, Wu L, Wang YS, Han XG (2008) Carbon and nitrogen store and storage potential as affected by land-use in a Leymus chinensis grassland of northern China. Soil Biol Biochem 40:2952–2959
Holt JA (1997) Grazing pressure and soil carbon, microbial biomass and enzyme activities in semiarid Northeastern Australia. Appl Soil Ecol 5:143–149
Iyyemperumala K, Israel DW, Shi W (2007) Soil microbial biomass, activity and potential nitrogen mineralization in a pasture: Impact of stock camping activity. Soil Biol Biochem 39:149–157
Lal R (2004) Soil carbon sequestration impacts on global climate change and food security. Sciences 304:1623–1627
Langley JA, Hungate BA (2003) Mycorrhizal controls on belowground litter quality. Ecology 84:2302–2312
Li YQ, Zhao HL, Zhao XY, Zhang TH, Chen YP (2006) Soil respiration, carbon balance and carbon storage of sandy grassland under post-grazing natural restoration. Acta Prataculturae Sinica 15:25–31, in Chinese with English abstract
Miller RH, Keeney DR (eds) (1982) Methods of soil analysis. Part 2: chemical and microbiological properties, 2nd edn. American Society of Agronomy, Soil Science Society of America, Madison
Moretto AS, Distel RA (1997) Competitive interactions between palatable and unpalatable grasses native to a temperate semiarid grassland of Argentina. Plant Ecol 130:155–161
Neill C, Melillo JM, Steudler PA, Cerri CC, Moraes JFL, Piccolo MC, Brito M (1997) Soil carbon and nitrogen stocks following forest clearing for pasture in the southwestern Brazilian Amazon. Ecol Appl 7:1216–1225
Nelson DW, Sommers LE (1982) Total carbon, organic carbon, and organic matter. In: Page AL, Miller RH, Keeney DR (eds) Methods of soil analysis. American Society of Agronomy and Soil Science Society of American, Madison, pp 1–129
Pettit NE, Froend RH, Ladd PG (1995) Grazing in remnant woodland vegetation: changes in species composition and life form groups. J Vege Sci 6:121–130
Piao SL, Fang JY, Ciais P, Peylin P, Huang Y, Sitch S, Wang T (2009) The carbon balance of terrestrial ecosystems in China. Nature 458:1009–1013
Reeves GW (2000) Bushcare program: mid-term review. CSIRO, Canberra. Retrieved 20 October 2000. http://www.nht.gov.au/mtrfinrpt/bushcarefinalreport.pdf
Shrestha G, Stahla PD (2008) Carbon accumulation and storage in semi-arid sagebrush steppe: effects of long-term grazing exclusion. Agric Ecosyst Environ 125:173–181
Smith RS, Shiel RS, Millward D, Corkhill P (2000) The interactive effects of management on the productivity and plant community structure of an upland meadow: an 8-year field trial. J Appl Ecol 37:1029–1043
Spooner P, Lunt I, Robinson W (2002) Is fencing enough? The short-term effects of stock exclusion in remnant grassy woodlands in southern NSW. Ecol Manage Rest 3:117–126
Tiessen H, Cuevas E, Chacon P (1994) The role of soil organic matter in sustaining soil fertility. Nature 371:783–785
Thomas RJ, Logan KAB, Ironside AD, Bolton GR (2006) The effects of grazing with and without excretal returns on the accumulation of nitrogen by ryegrass in a continuously grazed upland sward. Grass Forage Sci 45:65–75
Vance ED, Brooks PC, Jenkinson DS (1987) An extraction method for measure soil microbial biomass C. Soil Biol Biochem 19:703–707
Wright AL, Hons FM, Rouquette FM Jr (2004) Long-term management impacts on soil carbon and nitrogen dynamics of grazed bermudagrass pastures. Soil Biol Biochem 36:1809–1816
Wu GL, Du GZ, Liu ZH, Thirgood S (2009) Effect of fencing and grazing on a Kobresia-dominated meadow in the Qinghai–Tibetan Plateau. Plant Soil 319:115–126
Acknowledgements
We thank Dr. Gerlinde De Deyn and two anonymous referees for their valuable comments on this manuscript and thank Prof. Robin Pakeman for supporting in English writing revision and thank colleagues of Maqu Grassland Station for valuable assistance with field work. The study was supported by Projects of Natural Science Foundation of China (NSFC30900177) and Projects to Dr. Wu Gao-Lin from West Light Foundation of CAS (2009), the frontier research fund (10502-Q5) from State Key Laboratory of Soil Erosion and Dryland Farming on the Loess Plateau of ISWC of CAS, and NWSUAF (22050205) China and Special Research Funds for Public Service of Ministry of Agriculture China (200903060).
Author information
Authors and Affiliations
Corresponding authors
Additional information
Responsible Editor: Gerlinde De Deyn.
Rights and permissions
About this article
Cite this article
Wu, GL., Liu, ZH., Zhang, L. et al. Long-term fencing improved soil properties and soil organic carbon storage in an alpine swamp meadow of western China. Plant Soil 332, 331–337 (2010). https://doi.org/10.1007/s11104-010-0299-0
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11104-010-0299-0